INTRODUCTION
Many of the chemical fungicides currently used for plant disease control are classified as carcinogens by the Environ-mental Protection Agency (EPA). They are toxic to wildlife and other non-target species. Biological control offers an attractive alternative to synthetic chemical fungicides. Bio-pesticides that include living organisms and their naturally produced compounds can be safer, more biodegradable, and less expensive to develop. Various microorganisms in soil, including Enterobacter aerogenes, Pseudomonas fluore-scens, Pseudomonas cepacia, and Bacillus species, have been reported for their role in promotion of plant growth and biological control of plant pathogenic microorganisms. It is believed that plant growth-promoting rhizobacteria act
by displacing or antagonizing plant pathogenic microorgan-isms. Many of these rhizobacteria produce antibiotics, the production of which is strongly correlated with inhibition of various pathogenic microorganisms in vitro and disease suppression in vivo.
The use of bacteria like Pseudomonas sp. and Bacillus sp., have been investigated because of their properties to produce antifungal metabolites and protect plants from fungal infec-tion (Radheshyam et al. 1990; Moita et al. 2005; Siddiqui et al. 2005; Nourozian et al. 2006). Bacteria of the genus Bacillus produce a variety of peptide antibiotics that are antibacterial and/or antifungal. Several Bacillus species, including B. subtilis, B. pumilus and B. cereus, have been shown to be antagonistic to plant pathogenic fungi and bac-teria. Phytopathogen-antagonistic strains of B. subtilis have been reported to produce two peptide antibiotics: bacilysin, a dipeptide that inhibits yeast and bacteria; and fengycin (fengymycin), a lipopeptide antagonistic against
phytopath-─ ─ 279 ──
Characteristics of Bacillus Strains with Antifungal
Activity against Phytopathogens
Young-Keun Lee and M. Senthilkumar*
Radiation Application Research Division, Korea Atomic Energy Research Institute, Sinjeong, Jeongeup 580-185, Korea
Abstract -- Four bacterial isolates that showed antifungal activity against Alternaria alternata and other phytopathogens were isolates from bean rhizosphere. 16S rDNA analysis and phylogenetic relationship indicated that these isolates belong to Genus Bacillus. Isolate A1 clustered with Bacillus licheniformis while other isolates A2, A3 and A4 clustered together with B. pumilus. n-Butanol extract of these isolates strongly inhibited the growth of A. alternata while, chloroform extract of isolate A2 and ethyl acetate extract of A1, A3, and A4 inhibited the test fungus partially. All the isolates except A4 produced chitinase enzyme. None of the isolates solubilized mineral phosphate. Radiation sensitivity of isolates A1, A2, A3 and A4 were assessed and the LD99values are deter-mined as 0.50, 6.69, 11.60, 1.53 kGy, respectively. Mutant libraries of each isolate were prepared by exposing them to gamma radiation at their respective LD99dose. Crude metabolite caused dras-tic changes on A. alternata hyphal morphology. Appearance of shrunken and collapsed hyphae could be due to the leak of cell wall or changes in membrane permeability.
Key words : Antifungal metabolite, Bacillus pumilus, Bacillus licheniformis, Gamma irradiation
* Corresponding authors: M. Senthilkumar, Tel. +82-63-570-3304, Fax. +82-63-570-3309, E-mail. [email protected]
ogenic fungi such as Rhizoctonia solani (Loeffler et al. 1986; Vanittanakom et al. 1986).
Stabb et al. (1994) have identified certain strains belong to B. subtilis, B. cereus, B. mycoides, and B. thuringiensis exhibit antifungal activity. These strains have been shown to produce zwittermicin-A and/or kanosamine (Milner et al. 1996) that are effective against the soil borne disease damp-ing off, caused by Phytophthora medicaginis, P. nicotianae, P. aphanidermatum or Sclerotinia minor. Zwittermicin-A is a water soluble, acid stable linear aminopolyol molecule (He et al. 1994) with broad spectrum activity against many fungal and bacterial plant pathogens. The application of spore-based formulations of certain B. cereus strains to soy-bean seeds or the soil surrounding the seeds has been shown to improve soybean yield at field sites (Osburne et al. 1995). Kanosamine also inhibits a broad range of fungal plant pathogens and a few bacterial species (Milner et al. 1996). Bacillus licheniformis strain PR1-36a, isolated from a rhi-zosphere of perennial ryegrass (Lolium perenne L.) exhibited strong antagonism against filamentous fungi such as Rhizo-ctonia solani and Magnaporthe poae. It produces a diffusi-ble antifungal principle that can be obtained by acid precipi-tation of the culture filtrate followed by ethanol extraction.
Loeffler et al. (1986) reported that B. subtilis, B. pumilus, B. licheniformis, and B. coagulans strains produced various antibiotics with antifungal and antibacterial activity. Extra-cellular antifungal metabolites produced by Bacillus pumilus inhibited mycelial growth of many species of Aspergillus, Penicillium and Fusarium (Munimbazi and Bullerman 1998). B. pumilus produced bacilysin and iturin A. Bacilysin is a very small compound with a molecular weight of 270 that inhibits only yeast. The iturins, which are soluble in polar solvents, have broad antifungal and antibacterial activity. Antifungal activity of Bacillus licheniformis strains were enhanced through mutation and transformation (Cornea et al. 2003; Mateescu et al. 2004). The present study is aimed to isolate the bacterial strains with antifungal property and to establish their biocontrol potentials.
MATERIALS AND METHODS
1. Bacterial and fungal isolates
Soil samples were collected from bean rhizosphere and dry fields in Ipyeong-ri, Jeollanam-do province, South Korea. Samples were serially diluted and plated on nutrient agar Young-Keun Lee and M. Senthilkumar
280
Bacillus coagulans strain C1H: GQ214131 Bacillus cereus strain MG209: GQ280808
0.01
Bacillus velesensis strain CR-11: AY605932 Bacillus subtilis strain ATCC9799: AF142574 760
871
Bacillus licheniformis strain G7-2: EU256501 Bacillus malacitensis AY603656.2
779 985 898
1000
Bacillus axarquiensis AY603657.2
A4
Bacillus pumilus strain JS-45: GQ280115 A1
1000 549
Bacillus circulans strain JSC_SF51: DQ870737 855
A2 A3
670 1000
Bacillus vietnamensis strain MSU2510: AY647286 Bacillus pichinotyi strain RS2: AF519464 Bacillus casamancensis AF519462 702
999
Bacillus mojavensis AB021191
Bacillus aquaemaris strain TF-12: AF483625
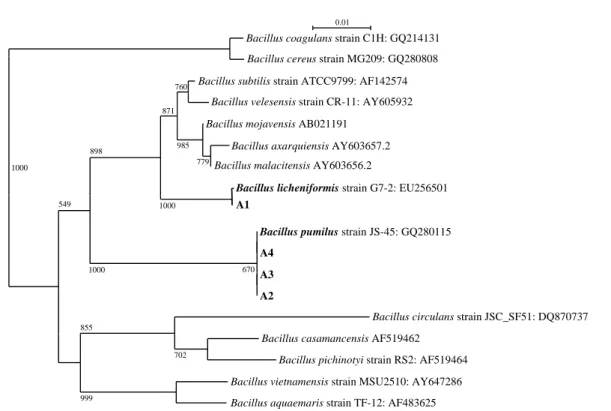
Fig. 1. 16S rDNA sequence based phylogenetic tree showing the relationship of Antifungal metabolite producing Bacillus isolates with other
(NA) plates. The colonies were bio-assayed on potato dex-trose agar (PDA) plates against Rhizoctonia solani and Alternaria alternata. Agar discs from the actively growing region of fungal pathogens were collected and placed at the center of the PDA plates. Bacterial isolated were streak ino-culated on the same plate at 2.5 cm away from the fungal disc and incubated at 28±2�C for 3~5 days. The plates were observed for the presence of a fungal growth inhibi-tion zone. The isolates showing antifungal property by inhi-biting the fungal growth were selected and maintained as 25% glycerol stock in nutrient broth under -80�C as well as refrigerated conditions. Fungal phytopathogens used in bio-assay as listed in Table 1, were maintained in PDA plates and stored under refrigerated condition.
2. Phylogenetic relationship of antifungal metabolite producing bacterial isolates
Bacterial isolates with antifungal trait were identified
based on 16S rDNA sequence analysis. Genomic DNA was extracted by using Qiagen DNeasy Plant Mini Kit (Qiagen, Valencia, CA). 16S rDNA was amplified by using the pri-mers 27F (5′-AGAGTTTGATCCTGGCTCAG-3′) and 1492R (5′-GGTTACCTTGTTACGACTT-3′) (Lane 1991). The sequences were determined by fluorescent dye termi-nators method using ABI prismTM BigdyeTM terminator cycle sequencing ready reaction kit V.3.1. Products were run on ABI 3730XL capillary DNA sequencer (ABI prism 310 Genetic analyzer, Tokyo, Japan). Sequences were sub-jected to BLASTn analysis with the NCBI data base and phylogenetic trees were generated with NJPLOT program-me of CLUSTAL X (Thompson et al. 1997).
3. Extraction of antifungal metabolite from Bacillus isolates
Bacillus isolates with antifungal activity were grown in 20 ml of potato dextrose broth for 62 h, 200 rpm at 28±2�C.
Fig. 2. Cell morphology of under Bacillus licheniformis strain A1 and strains A2, A3 and A4 of Bacillus pumilus under scanning electron
The culture supernatant was prepared by centrifugation at 5,000 rpm for 20 min and added with an equal volume of n-hexane. The organic phase was separated with separating funnel and tested for antifungal activity with A. alternata. The same aqueous phase was again extracted with equal volume of chloroform, ethyl acetate and n-butanol sequen-tially. Aqueous as well as organic fractions were tested for antifungal activity by disc diffusion assay with A. alternata. Active crude extracts were dried completely and dissolved in HPLC-grade methanol.
4. Chitinase activity and mineral phosphate solubilization
Ability of Bacillus isolates to produce chitinase was tested on nutrient agar medium supplemented with 0.5% colloidal chitin. Colloidal chitin was prepared by the method of Rod-riguez-Kabana et al. (1983) by partial hydrolysis of chitin (Sigma, St. Louis, USA) with 10 N HCl for 2 h at room tem-perature. The colloidal chitin was washed several times with large volumes of distilled water to adjust the pH to 7.0.
Phosphate solubilizing ability was detected by spotting the isolates on Pikovskaya agar (Pikovskaya 1948) as well as NBRIP-BPB agar plates (Mehta and Nautiyal 2001). Plates were then incubated at 28�C for 3 days and observed for the clearing zone around the colonies.
5. Radiation sensitivity
Cell pellets were collected by centrifuging 20 ml of a log phase bacterial isolate cultured in the LB broth. Pellets were washed twice with sterile-distilled water and suspended (~107~109CFU ml-1). 500μl of cell suspensions were
trans-ferred to 1.5 ml micro-centrifuge tubes and irradiated at dif-ferent doses expressed in kGy, as shown in Fig. 3. Samples were irradiated in a cobalt-60 irradiator (capacity: 250000 Ci, dose rate 920 Gy/hr, AECL) at the Advanced Radiation Technology Institute, Korea Atomic Energy Research Insti-tute, South Korea. The bacterial population in irradiated samples was determined by serial dilution and plate count method. The D10-value was determined by plotting a radia-tion dose (kGy) on the X-axis and a survival ratio on the Y-Young-Keun Lee and M. Senthilkumar
282 -1 0 -2 -1 00 0.5 1 1.5 2 2.5 0 2 4 6 8 -3 -2 -8 -7 -6 -5 -4 -3 0 -4 -9 0 -1 -5 -4 -3 -2 -1 -3 -2 -8 -7 -6 Radiation sensitivity (kGy)
Radiation sensitivity (kGy)
Radiation sensitivity (kGy)
0 1 2 3 4 5
Radiation sensitivity (kGy) Strain A1 Strain A3 Strain A4 Strain A2 Log(Nx/No) Log(Nx/No) Log(Nx/No) Log(Nx/No) 0 1 2 3 4 5 6 7 8 9 10
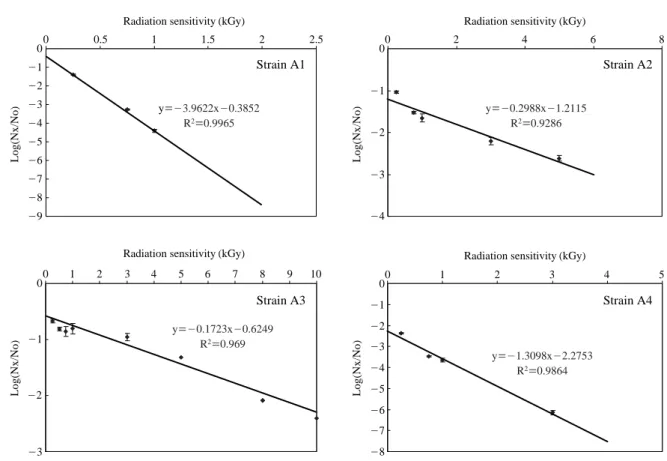
Fig. 3. Radiation sensitivity of Bacillus licheniformis strain A1 and strains A2, A3 and A4 of Bacillus pumilus. y==-3.9622x-0.3852 R2==0.9965 y==-0.1723x-0.6249 R2==0.969 y==-1.3098x-2.2753 R2==0.9864 y==-0.2988x-1.2115 R2==0.9286
axis. The negative reciprocal of the slope indicated the D10 -value. LD99value, the radiation dose required to kill 99% of viable bacterial cells was determined by using the formula LD99==log (0.01)/slope.
6. Scanning electron microscopy
Bacterial cells were fixed directly in glutaraldehyde buffer for 4 h and then dehydrated. Mycelium of R. solani was treat-ed with a crude antifungal metabolite with a concentration of 10 mg ml-1of water. Hyphae were removed after 5 h of
incubation, fixed in glutaraldehyde buffer for 4 h in room temperature, dehydrated in graded alcohol, and dried. Gold sputter-coating was carried out under reduced pressure in an inert argon gas atmosphere (Agar sputter coater P7340). After sputter-coating, specimens were examined under a scanning electron microscope (JSM 6390) operated at 30 kV.
RESULTS AND DISCUSSION
Rhizobacteria that are able to colonise the root region take
advantage of the root exudates released by the plant. This interaction is beneficial both for the plant as well as the bacteria. The presence of non-pathogenic bacteria at the roots confers protection against pathogenic microorganisms. This protection by bacteria can be mediated due to produc-tion of antibiotics harmful to other microorganisms (Wulff et al. 2002; Bais et al. 2004). Formation of biofilm on plant roots by the bacteria can make the plants less sensitive to infection (Bais et al. 2004; Rudrappa et al. 2008). In this study, bean rhizosphere soil and dry field soil samples were serially diluted and plated on nutrient agar plates. Colony with different morphology and colour were purified and stored under refrigerated condition. Bacterial isolates were tested for their antifungal activity against A. alternata by dual culture assay. Four bacterial isolates that inhibited the growth of A. alternata were selected and designated as A1, A2, A3 and A4. The antagonistic activity of these isolates against other fungal pathogens as listed in Table 1 was demonstrated by dual culture assay. All the Bacillus isolates inhibited the growth of A. alternata, A. solani, Botrytis cinerea, and Rhizoctonia solani in vitro by secreting
anti-Fig. 4. Effect of crude antifungal metabolite(s) on A. alternata. A) Untreated hyphae, B) Hyphae treated with crude antifungal metabolite(s)
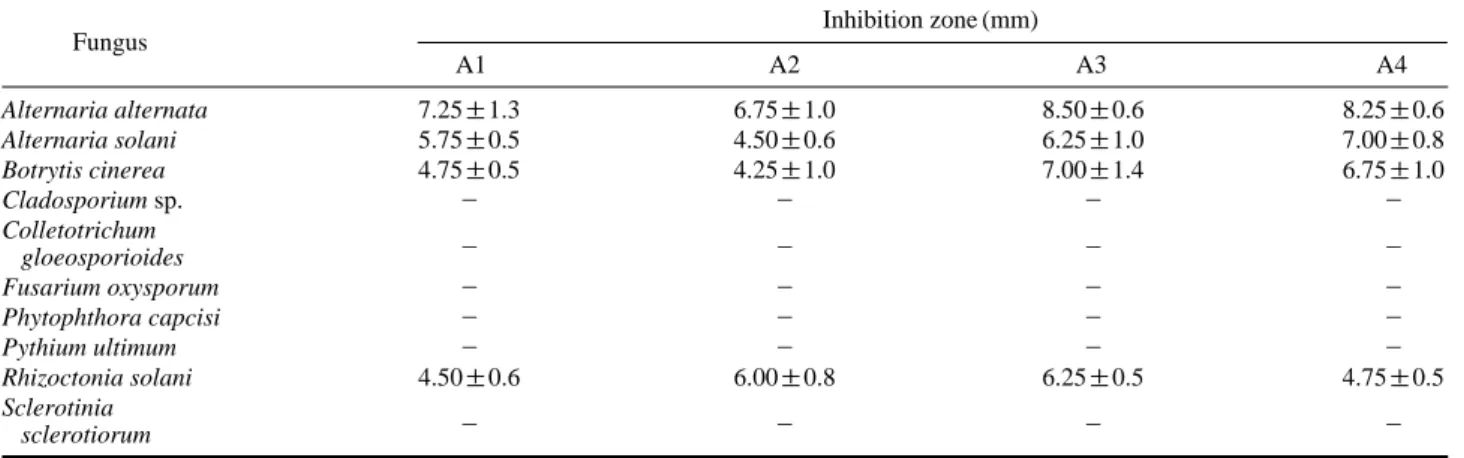
fungal compound(s) out of the bacterial cells, as indicated by the formation of inhibition zone. These isolates did not produce inhibition zone against other phytopathogenic fungi including Cladosporium, Colletotrichum gloeosporioides, Fusarium oxysporum, Phytophthora capcisi, Pythium ulti-mum and Sclerotinia sclerotiorum though the fungal growth was reduced. Three days after incubation in dual culture assay, the fungi over grew the bacterial colony.
All the four bacterial isolates are Gram positive rods. Cell size ranges from 0.3μm~0.4 μm width and 1.3 μm~1.8 μm length. 16S rDNA analysis indicated that the isolates belong to Genus Bacillus. To study the phylogenetic rela-tionship, neighbor-joining dendrogram was generated for our isolates and representative Bacillus sequences from Gen-Bank. Isolate A1 clustered with Bacillus licheniformis while other isolates A2, A3 and A4 clustered together with B. pumilus (Fig. 1). We attempted to extract the antifungal metabolites of our isolates. None of the n-hexane extracts showed the antifungal activity against A. alternata. Chlo-roform extract of isolate A2 partially inhibited the fungal growth. Chloroform extracts of other isolates did not inhibit the test fungus. Most of the antifungal metabolites from these isolates were extracted with n-butanol and showed strong zone of inhibition. Ethyl acetate extract of isolates A1, A3 and A4 partially inhibited the fungus and longer incubation time allowed the fungus to over grow the anti-biotic discs. The Bacillus isolates were also tested for the production of chitinase and mineral phosphate solubilization. Except isolate A4, others produced chitinase enzyme. None of our isolates solubilized mineral phosphate in Pikovskaya agar medium. The results were also confirmed in NBRIP-BPB agar plates. Radiation mutagenesis is considered as a
tool to modify the traits of rhizobacteria in order to enhance their activity and to understand their basic mechanisms of plant growth promotion. We initially studied the radiation sensitivity of our Bacillus isolates. The D10and LD99values were determined by plotting a radiation dose (kGy) on the X-axis and a survival ratio on the Y-axis. Isolates A1, A2, A3 and A4 recorded the D10values of 0.25, 3.35, 5.80, and 0.76 kGy respectively. LD99values of isolates A1, A2, A3 and A4 are 0.50, 6.69, 11.60, 1.53 kGy, respectively. Isolate A3 is comparatively resistant to radiation while isolate A1 is highly susceptible. Bacillus isolates were exposed to gam-ma radiation at their LD99dose. Mutant clones were stored as 25% glycerol stocks under -80�C. Mutant clones can be screened either for enhanced antifungal activity or defective mutants. Several antifungal antibiotics caused morphologi-cal abnormalities on filamentous test fungi. Polyoxin caused formation of characteristic bulges or swellings on the grow-ing hyphae and spores of sensitive filamentous fungi such as Piricularia oryzae and Mucor racemosus. Tunicamycin, a specific inhibitor of glycoprotein synthesis in yeast, caused bursting, swelling of Penicillium chrysogenum (Yoshiki et al. 1976). Griseofulvin induced hyphal curling of several fungi tested. The most remarkable curling was observed with Paecilomyces varioti. Inhibitors of protein synthesis such as cycloheximide and blasticidin S and inhibitors of energy metabolism such as antimycin and oligomycin showed slight or no effect on the morphology of the tested organisms in the case of higher concentrations than their MIC values (Gunji et al. 1983). Microscopic observation on the effect of crude metabolites of our Bacillus isolates on A. alternata hyphal morphology was performed under scanning electron microscope. Crude metabolite caused drastic changes on Young-Keun Lee and M. Senthilkumar
284
Table 1. Antifungal activity of Bacillus strains against phyto-pathogens
Fungus Inhibition zone (mm)
A1 A2 A3 A4 Alternaria alternata 7.25±1.3 6.75±1.0 8.50±0.6 8.25±0.6 Alternaria solani 5.75±0.5 4.50±0.6 6.25±1.0 7.00±0.8 Botrytis cinerea 4.75±0.5 4.25±1.0 7.00±1.4 6.75±1.0 Cladosporium sp. - - - -Colletotrichum - - - -gloeosporioides Fusarium oxysporum - - - -Phytophthora capcisi - - - -Pythium ultimum - - - -Rhizoctonia solani 4.50±0.6 6.00±0.8 6.25±0.5 4.75±0.5 Sclerotinia - - - -sclerotiorum
hyphal morphology. Depressions were developed on the sur-face of treated hyphae. Appearance of shrunken and col-lapsed hyphae could be due to the leak of cell wall or changes in membrane permeability. Hyphal swelling and lysis were also observed. In conclusion, present work established the antagonistic activity of Bacillus isolates A1, A2, A3 and A4 against major phytopathogens. Further research on puri-fication and characterization of antifungal metabolites will help to understand the detailed mechanism of plant disease suppression through biological control.
ACKNOWLEDGMENT
This study was carried out under the Nuclear R&D Pro-gram of the Ministry of Education, Science and Technology, Republic of Korea.
REFERENCES
Bais HP, Fall R and Vivanco JM. 2004. Biocontrol of Bacillus
subtilis against infection of Arabidopsis roots by Pseudo-monas syringae is facilitated by biofilm formation and
surfactin production. Plant Physiol. 134:307-319.
Cornea CP, Mateescu R, Grebenisan I, Vamanu E and Cam-peanu G. 2003. Isolation and characterization of new
Bacil-lus spp. strains-useful as biocontrol agents of plant
patho-gens. Rom. Biotechnol. Lett. 8:1115-1122.
Gunji S, Arima K and Beppu T. 1983. Screening of antifungal antibiotics according to activities inducing morphological abnormalities. Agric. Biol Chem. 47:2061-2069.
He H, Silo-Suh LA, Clardy J and Handelsman J. 1994. Zwitter-micin A, an antifungal and plant protection agent from Bacillus cereus. Tetrahedron Lett. 35:2499-2502.
Lane DJ. 1991. 16S/23S rRNA sequencing. pp. 115-175. In: Nucleic Acid Techniques in Bacterial Systematics (Stacke-brandt E and Goodfellow M eds.), Chichester, Wiley. Loeffler W, Tschen SM, Vamittanakoon N, Kugler M, Knorpp
E, Hsieh TF and Wu TG. 1986. Antifungal effects of bacily-sin and fengymycin from Bacillus subtilis F-29-3. A com-parison with activities of other Bacillus antibiotics. J.
Phyto-pathol. 115:204-213.
Mateescu R, Cornea CP, Grebenisan I, Babeanu N and Cam-peanu G. 2004. In vivo biocontrol activity of Bacillus spp. strains on Alternaria tenuis. Rom. Biotechnol. Lett. 9:1713-1721.
Mehta S and Nautiyal CS. 2001. An efficient method for
quali-tative screening of phosphate solubilizing bacteria. Curr
Microbiol. 43:51-56.
Milner JL, Silo-Suh L, Lee JC, He H, Clardy J and Handelsman J. 1996. Production of kanosamine by Bacillus cereus UW85. Appl. Environ. Microbiol. 62:3061-3066.
Moita C, Feio SS, Nunes L, Curto MJM and Roseiro JC. 2005. Optimization of physical factors on the production of active metabolites by Bacillus subtilis 355 against wood surface contaminant fungi. International J. Biodeterioration &
Biodegradation 55:261-269.
Munimbazi C and Bullerman LB. 1998. Isolation and partial characterization on antifungal metabolites of Bacillus
pumi-lus. J. Appl. Microbiol. 84:959-968.
Nourozian J, Etebarian HR and Khodakaramian G. 2006. Bio-logical control of Fusarium graminearum on wheat by anta-gonistic bacteria. J. Sci. Technol. 28:29-38.
Osburne RM, Milner JL, Oplinger ES, Smith RS and Handels-man J. (1995). Effect of Bacillus cereus UW85 on the yield of soybean at two field sites in Wisconsin. Plant Dis. 79: 551-556.
Pikovskaya RI. 1948. Mobilization of phosphorus in soil con-nection with the vital activity of some microbial species.
Microbiologiya 17:362-370.
Radheshyam K, Fernandez MA and Schroeder III RG. 1990. Isolation and characterization of a Pseudomonas strain that restrict growth of various phytopathogenic fungi. Appl.
Environ. Microbiol. 56:1053-1058.
Rodriguez-Kabana R, Godoy G, Morgan-Jones G and Shelby RA. 1983. The determination of soil chitinase activity: con-ditions for assay and ecological studies. Plant Soil. 75: 95-106.
Rudrappa T, Biedrzycki M, Bais HP. 2008. Causes and con-sequences of plant-associated biofilms. FEMS Microbiol.
Ecol. 64:153-166.
Siddiqui S, Siddiqui ZA and Ahmad I. 2005. Evaluation of fluorescent Pseudomonas and Bacillus isolates for the bio-control of a wilt complex of pigeonpea. World J. Microbiol.
Biotechnol. 21:729-732.
Stabb EV, Jacobson LM and Handelsman JO. 1994. Zwitter-micin-A producing strains of Bacillus cereus from diverse soils. Appl. Environ. Microbiol. 60:4404-4412.
Thompson JD, Gibson TJ, PleWniak F, Jeanmougin F and Hig-gins DG. 1997. The clustalx windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 24:4876-4882.
Vanittanakom N, Loeffler W, Koch U and Jung G. 1986. Fengy-cin-a novel antifungal lipopeptide antibiotic produced by
Bacillus subtilis F-29-3. J. Antibiot. 39:888-901.
Wulff EG, Mguni CM, Mortensen CN, Keswani CL and Hock-enhull J. 2002. Biological control of black rot (Xanthomonas
campestris pv. campestris) of Brassicas with an antagonistic
strain of Bacillus subtilis in Zimbabwe. Eur. J. Plant Pathol.
108:317-325.
Yoshiki K, Kuninaka A, Yoshino H, Takatsuki A, Yamasaki M and Tamura G. 1976. Formation of fungal multinuclear giant
cells by tunicamycin. Gen. Appl. Microbiol. 22:247-258.
Manuscript Received: December 4, 2009 Revision Accepted: December 18, 2009 Young-Keun Lee and M. Senthilkumar