저작자표시-비영리-변경금지 2.0 대한민국 이용자는 아래의 조건을 따르는 경우에 한하여 자유롭게 l 이 저작물을 복제, 배포, 전송, 전시, 공연 및 방송할 수 있습니다. 다음과 같은 조건을 따라야 합니다: l 귀하는, 이 저작물의 재이용이나 배포의 경우, 이 저작물에 적용된 이용허락조건 을 명확하게 나타내어야 합니다. l 저작권자로부터 별도의 허가를 받으면 이러한 조건들은 적용되지 않습니다. 저작권법에 따른 이용자의 권리는 위의 내용에 의하여 영향을 받지 않습니다. 이것은 이용허락규약(Legal Code)을 이해하기 쉽게 요약한 것입니다. Disclaimer 저작자표시. 귀하는 원저작자를 표시하여야 합니다. 비영리. 귀하는 이 저작물을 영리 목적으로 이용할 수 없습니다. 변경금지. 귀하는 이 저작물을 개작, 변형 또는 가공할 수 없습니다.
New Therapeutic Strategies for
Amyotrophic Lateral Sclerosis Treatment
by
Chan-Il Choi
Major in Neuroscience
Department of Medical Sciences
The Graduate School, Ajou University
New Therapeutic Strategies for
Amyotrophic Lateral Sclerosis Treatment
by
Chan-Il Choi
A Dissertation Submitted to The Graduate School of Ajou University
in Partial Fulfillment of the Requirements for the Degree of
Ph. D. in Neuroscience
Supervised by
Haeyoung Suh-Kim, Ph.D.
Major in Neuroscience
Department of Medical Sciences
The Graduate School, Ajou University
This certifies that the dissertation
of Chan-Il Choi is approved.
The Graduate School, Ajou University
December, 21st, 2009
i - ABSTRACT -
New Therapeutic Strategies
for Amyotrophic Lateral Sclerosis Treatment
Amyotrophic lateral sclerosis (ALS) is characterized by progressive dysfunction and degeneration of motor neurons in cerebral cortex, brain stem and spinal cord. Degeneration of motor neurons causes muscle weakness and respiratory paralysis, leading to decease of ALS patients. Approximately 10% of the cases are familial and 20% of those are dominantly inherited mutations in superoxide dismutase 1 (SOD1). The transgenic animal model of ALS was the G93A mouse that expresses a mutant form of human SOD1 carrying the Gly93 → Ala and develops motor neuron disease similar to familial ALS. With the lack of effective drug treatments for ALS and compelling preclinical data, new therapeutic strategies have highlighted this disease as a candidate for stem cell treatment.
Here we investigated whether neural induction enhances therapeutic potentials of mesenchymal stem cells (MSCs). We induced neural induction of MSCs by overexpressing Neurogenin1, a proneural transcription factor. Depending on transplantation time, systemic transplantation of Ngn1-expressing MSCs (MSCs-Ngn1) delayed the onset of neurobehavioral degeneration or dramatically improved motor functions compared to parental MSCs in mice carrying a high copy number of a human mutant SOD1(G93A) transgene. We found that compared to MSCs, more MSCs-Ngn1 were found in hippocampus, brainstem, and the spinal cord, where most of them remained undifferentiated. We also found that more motor neurons were preserved in the spinal cord of animals with MSCs-Ngn1, suggesting that MSCs-Ngn1 migrate to the central nervous system and exert
ii
beneficial effects on host neural cells through paracrine signaling for extended period of time. Our findings strongly suggest potential benefits of neural induction of MSCs to develop successful stem cell therapy for the treatment of ALS.
Glial cell line-derived neurotrophic factor (GDNF), which has been demonstrated to be the most potent neurotrophic factor for the proliferation, differentiation, and survival of spinal motor neurons, exhibits very good therapeutic potential for ALS. MSCs were infected with recombinant adenovirus GDNF (MSCs-GDNF). The MSCs-GDNF were transplanted into tail vein of ALS mice (13, 15 weeks of age), and the animals were evaluated on health and behavioral measures. Transplantation of the MSC-GDNF into tail vein delayed the onset of motor behavioral symptoms, and prolongs the life span.
The incidence of ALS is higher in men than women, but the female advantage disappears with increased age. Here, we report evidence that the female advantage is due to the protective role of estrogen. In an ALS mouse model carrying the human Cu/Zn superoxide dismutase (hSOD1) G93A transgene, ovariectomy did not alter the onset age of the disease while reducing the female lifespan by 7 days and making it comparable to that of the male transgenic mice. Treatment of ovariectomized females with 17β-estradiol (E2) did not delay the onset of disease, but prevented progression of ALS motor dysfunctions as shown by extension reflex test for a limited time window. Importantly, E2 treatment rescued the lifespans in overiectomized females. These findings will provide important new insights to strategy of ALS treatment.
__________________________________________________________________________ Key words: Mesenchymal Stem Cells (MSCs), Amyotrophic Lateral Sclerosis (ALS), Neurogenin1 (Ngn1), Glial cell line-Derived Neurotrophic Factor (GDNF), SOD1G93A mouse, gender, estrogen, ovariectomy
iii
TABLE OF CONTENTS
ABSTRACT ··· ⅰ TABLE OF CONTENTS ··· ⅲ LIST OF TABLES ··· ⅵ LIST OF FIGURES ··· ⅶ Ⅰ. INTRODUCTION ··· 1Ⅱ. MATERALS AND METHODS ··· 6
A. Materials ··· 6
B. Methods ··· 7
1. Common Methods ··· 7
1.1. Animals ··· 7
1.2. Evaluation of motor functions ··· 7
1.3. Assessment of disease onset and lifespan ··· 9
1.4. Statistical analysis ··· 9
2. (Part1) ··· 10
2.1. Preparation of MSCs-Ngn1 cells and in vitro differentiation ··· 10
2.2. Transplantation ··· 10
2.3. Histological analysis of transplanted cells ··· 11
2.4. Quantification of motor neurons and Iba1+ microglia ··· 12
3. (Part2) ··· 12
3.1. Western blot analysis for GDNF ··· 13
iv
3.3. Transplantation ··· 14
3.4. Motor neuron counting ··· 14
4. (Part3) ··· 14
4.1. Ovariectomy and estrogen replacement ··· 14
4.2. Measurement of E2 concentration and uterine weight ··· 15
Ⅲ. RESULTS ··· 16
A. (Part1) ··· 16
1. In vitro cultivation of MSCs-Ngn1 ··· 16
2. Distribution of Transplanted MSCs-Ngn1 in the Central Nervous System ··· 16
3. Protection of Spinal Motor Neurons ··· 21
4. Immune modulation by transplanted cells ··· 24
5. Delayed disease progression by MSCs-Ngn1 ··· 24
6. Enhanced therapeutic effects of MSCs-Ngn1 by repeated grafting ··· 28
B. (Part2) ··· 35
1. Evaluation of cytotoxicity and expression efficiency of adeno-GDNF virus ··· 35
2. Effect of MSCs-GDNF administration on motor neuron survival and disease progression ··· 38
C. (Part3) ··· 42
1. Gender differences in onset and disease progression ··· 42
v
Ⅳ. DISCUSSION ··· 51
Ⅴ. CONCLUSIONS ··· 61
REFERENCES ··· 63
vi
LIST OF TABLES
Table. 1. Experimental protocols and summary results (MSCs-Ngn1).
··· 31 Table. 2. Summary results (MSCs-GDNF).
··· 41 Table. 3. Summary of gender differences and E2 effects.
··· 49 Supplementary Table. 1. Summary results of male and female mice
(ExperimentⅠ). ··· 32 Supplementary Table. 2. Summary results of male and female mice
(ExperimentⅡ). ··· 33 Supplementary Table. 3. Summary results of male and female mice
vii
LIST OF FIGURES
Fig. 1. Evaluation of motor functions. ··· 8
Fig. 2. Neuronal differentiation of MSC-Ngn1 in vitro. ··· 17
Fig. 3. Migration of transplanted MSCs-Ngn1 into the CNS and immunophenotypes of MSCs-Ngn1 within the CNS. ··· 18
Fig. 4. Migration of transplanted MSCs-Ngn1 into other organs. ··· 19
Fig. 5. Expression of cytokine-receptors in MSCs and MSCs-Ngn1. ··· 20
Fig. 6. Counting of survived motor neuron. ··· 22
Fig. 7. Effect of MSCs and MSCs-Ngn1 on the neuroinflammation. ··· 23
Fig. 8 Onset of disease, total survival time and characteristics of disease progression in MSCs-Ngn1 transplanted ALS mice (treatment in the pre-onset stage). ··· 26
Fig. 9 Total survival time and characteristics of disease progression in MSCs-Ngn1 transplanted ALS mice (treatment in the disease-onset stage). ··· 27
Fig. 10. Total survival time and characteristics of disease progression in MSCs-Ngn1 transplanted ALS mice (treatment in the disease-onset stage, 2 times). ··· 29
Fig. 11. Total survival time and characteristics of disease progression in CsA injected ALS mice. ··· 30
Fig. 12. Safety and transduction efficiency test of recombinant adeno-GDNF virus. ··· 36
viii
Fig. 13. Confirmation of GDNF expression in vitro and in vivo. ··· 37
Fig. 14. Counting of survived motor neuron. ··· 39
Fig. 15. Total survival time and characteristics of disease progression in MSCs-GDNF transplanted ALS mice (treatment in the disease-onset stage, 2 times). ··· 40
Fig. 16. Gender differences in onset of ALS symptoms and eventual lifespan. ··· 43
Fig. 17. Gender differences in motor functions. ··· 44
Fig. 18. Verification of ovariectomy and E2 replacement. ··· 45
Fig. 19. Protective effects of estrogen in the ovariectomized mice. ··· 47
1
I. INTRODUCTION
A. Amyotrophic Lateral Sclerosis
Amyotrophic lateral sclerosis (ALS) is an adult-onset degenerative disease that is characterized by the death of motor neurons in the cortex, brain stem, and spinal cord. Patients with ALS exhibit progressive muscle atrophy and respiratory paralysis, and usually die within 3-5 years after symptom onset (Bradley 1995; Brown 2001; Przedborski 2003; Rothstein 2003). About 10% of ALS are known familial ALS (fALS), and 20% of fALS are associated with mutations in the Cu2+/Zn2+ superoxide dismutase 1 (SOD1) gene (Rosen 1993). The remaining 90% of ALS are diagnosed sporadic for which few genetic components are known (Fiszman 1999; Pramatarova 1995). These include oxidative stress and glutamate excitotoxicity (Rothstein 1996; Shaw 1997), viral infection (Berger 2000), autoimmune mechanisms (Alexianu 1995; Appel 1995; Niebroj-Dobosz 1999), abnormal accumulations of neurofilaments (Beaulieu 2000; Warita 1999), altered glial function (Cho 1999; Hall 1998; Levine 1999; Migheli 1999), mitochondrial dysfunction (Menzies 2002; Sasaki 1996), and the impaired trophic supports from the host environment (Hall 1998) (reviewed in (Garbuzova-Davis 2003)). Due to this diverse pathogenesis, there is no effective treatment available for this disease.
B. (Part1) Neural Induction with Neurogenin1 Prolongs Therapeutic Effects of Mesenchymal Stem Cells in an Amyotrophic Lateral Sclerosis Mouse Model.
Recently stem-cell treatment has emerged as a therapeutic candidate for the treatment of ALS. Multiple factors underlying ALS, it is difficult to envision that pathological changes
2
can be delayed merely by replacement or restoration of the motor neurons. More likely, transplanted stem cells of nonneuronal origin have additional mechanisms preventing degeneration of nearby motor neurons when transplanted in a mouse model of ALS carrying the human SOD1 transgene with G93A mutation. These cells include human umbilical-cord-blood derived cells (Ende 2000; Garbuzova-Davis 2008; Garbuzova-Davis 2003; Habisch 2007; Rizvanov 2008), bone marrow (mesenchymal) stem cells (Ende 2000; Habisch 2007; Morita 2008; Suzuki 2008; Vercelli 2008; Zhao 2007) and sertoli cells (Hemendinger 2005). Stem cells with neural origin such as human neuron-like cells (Garbuzova-Davis 2001; Garbuzova-Davis 2002; Garbuzova-Davis 2006; Willing 2001) and neural stem cells (Corti 2007; Klein 2005; Martin 2007; Morita 2008; Suzuki 2007; Xu 2006; Yan 2006) are effective to postpone the disease progression in animal model.
Mesenchymal stem cells (MSCs) are defined as pluripotent progenitor cells that retain the ability to generate cartilage, bone, muscle, tendon, ligament and fat. (Beyer Nardi 2006; Bianco 2000; Bianco 2001; Deans 2000; Prockop 1997; Sethe 2006). Clinical studies have suggested that these cells could improve the neurological symptoms after transplanted in patients with brain ischemia (Bang 2005) and in animal models of the spinal cord injury (Hofstetter 2002). Though stem cell therapy is an exciting candidate for the treatment of ALS, there are still many hurdles to be overcome before it can be translated into clinical treatment. Transplantation of MSCs into the spinal cord of patients with ALS did not yield clinical improvement (Ferrero 2008; Mazzini 2003; Mazzini 2006; Mazzini 2008). Moreover, there is little consensus on the effects of cell transplantation in animal models. Some experiments showed beneficial effects on animal clinical parameters (Chen 2000; Corti 2004; Ende 2000; Garbuzova-Davis 2002; Garbuzova-Davis 2003; Willing 2001),
3
while others showed no beneficial effects (Habisch 2007; Hemendinger 2005). It is still unknown which parameters such as the cell types, the time point of transplantation, and the administration route may affect the consequence of transplantation (reviewed in (Morita 2008)). Most of all, it is difficult to compare the effectiveness of cell types with neural and non-neural origins.
It is controversial whether MSCs can differentiate into ectodermal neuronal lineages or not (Azizi 1998; Coyne 2006; Deng 2001; Kopen 1999; Lu 2004; Munoz-Elias 2003; Neuhuber 2004; Woodbury 2000). Previously, we report that a proneural transcription factor, Neurogenin1 (Ngn1) is sufficient to reprogram the mesodermal cell fate of human MSCs into a neuronal one (Kim 2008). NeuroD and voltage-gated Ca2+ and Na+ channels are
expressed in Ngn1-expressing MSCs (MSCs-Ngn1). Moreover, MSCs-Ngn1 show higher therapeutic effects than MSCs in an animal stroke model. This is due to the fact that unlike MSCs, MSCs-Ngn1 can differentiate into neurons in the ischemic brain and functionally connect to host neurons. MSCs and MSCs-Ngn1 are equivalent to provide neurotrophic supports to the host brain. These data suggest that stem cells with neuronal property may be advantageous to treat neurological diseases.
In the present study, we compared effectiveness of the cell types (MSCs versus Ngn1-expressing MSCs) for the treatment of ALS in a hSOD1G93A transgenic mouse model. We
also compared the transplantation timing and the number of grafted cells in delaying the progression of ALS symptoms. The results of this study may help to interpret the conflicting results in the previous studies with stem cells, and provide insights to improve the stem cell technologies for the treatment of ALS.
4
C. (Part2) Transplantation of Mesenchymal Stem Cells Transduced with GDNF Prolongs Lifespan and Ameliorates Motor Dysfunctions in an Amyotrophic Lateral Sclerosis Mouse Model.
GDNF is an approximately 20 kDa, glycosylated polypeptide that exists in its native form as a homodimer (Lin 1993). The gene for GDNF has been mapped to human chromosome 5 at p12-p13.1 (Schindelhauer 1995), and gives rise to two alternatively-spliced forms that code for prepropeptides of 211 and 185 amino-acid residues respectively (Springer 1995; Cristina 1995). The 26 amino-acid residue deletion in the short form occurs in the pro-segment of the precursor. Thus both the long (a) and short (b) forms yield equivalent 134 amino-acid residue mature forms after proteolytic cleavage. As noted above, GDNF is now considered a member of the TGF-b superfamily (Lin 1993).
The activity most often attributed to GDNF is that of promoting neuron survival. This is manifested in many forms by many different neuron cell types. For example, dopaminergic neurons removed from embryonic midbrains can be rescued from destruction in vitro by GDNF (Krieglstein 1995). In addition, GDNF can interrupt the apoptotic programs of embryonic avian motor neurons in vivo. Furthermore, autonomic (involuntary) motor neurons of both sympathetic and parasympathetic systems also respond to GDNF by maintaining their numbers in vitro at significant levels relative to controls (Ebendal 1995). In
vivo, following transection of (voluntary) facial motor neuron axons, exogenously applied
GDNF has been shown to rescue virtually all damaged neurons from death, while transected, non-treated neurons almost all perish (Yan 1995). The action of GDNF as a neurotrophic factor for motor neurons suggests the potential usefulness of this factor in treating diseases affecting motor neurons. The purpose of this study was to show that transplantation of MSCs
5
transduced with adeno-GDNF virus into the tail vein of a transgenic ALS model mouse would migrate widely into the CNS and improve motor deficits.
D. (Part3) Effects of estrogen on lifespan and motor functions in female Amyotrophic Lateral Sclerosis Mouse Model.
Clinical studies have shown that among individuals younger than 50 years of age, men are at a higher risk for ALS than women (Haverkamp 1995). However, this gender difference declines gradually with equal numbers of males and females seen among older ALS patients, suggesting that female hormones may play a beneficial role against the onset and progression of ALS.
The most widely used animal models for ALS are mice carrying a transgene that encodes human SOD1 harboring a glycine to alanine mutation at amino acid 93 (hSOD1G93A).
The phenotypes of these mice closely recapitulate human ALS in terms of clinical and histo-pathological symptoms, with the added benefits of relatively early onset and rapid disease progression (Gurney 1994). Although earlier studies reported gender differences in some aspects, the female advantage in the ALS animal model is not clearly established.
In this study, we investigated the gender differences in the ALS animal models in multiple aspects such as onset ages, progression of motor symptoms, and lifespan. We also determined the molecular identity responsible for the female advantage. These findings will provide important new insights to interpretation of disease progression in post-menopausal female ALS patients,
6
II.
MATERIALS AND METHODS
A. Materials
Transgenic ALS mice carrying a human SOD1 transgene with a missense mutation that substitutes alanine for glycine at codon 93 (G93A) were obtained from the Jackson Laboratories. All surgical instruments were purchased from Solco Biomedical (Pyungtaek, Korea). Rotarod tester was purchased from UGO BASILE (Acceler Rota-Rod 7650, Varese, Italy). Paw Grip Endurance tester was purchased from Nil-Bio (Suwon, Korea). Anti-neurogenin1 specific antibody was purchased from Chemicon International (Temecula, CA, U.S.A); Anti-GDNF antibody from R&D systems (U.S.A); Taq DNA polymerase from SUPERBIO CO., LTD (Korea); PDVF membrane from Millipore (Bedford, MA, U.S.A); Super Signal kit from Pierce (Rochford, IL, U.S.A).
Retroviral vector, pMSCV-puro was purchased from CLONTECH Laboratory (Palo Alto, CA, U.S.A). Ad-GDNF virus was provided from Sun-Yong Baek et al., (Pusan National University); Human recombinant bFGF from Dong-A Pharmaceutical Research Laboratories (Yong-In, Korea). DMEM and FBS ware purchased from GibcoBRL (Grand Island NY, U.S.A). estrogen pellet containing 0.18 mg of 17β-estradiol (E2) (IRA, Sarasota, FL, USA). Enzyme Immunoassay kit for estradiol (Oxford Biomedical Research Inc., Oxford, MI, USA). Besides, all other chemicals were purchased from Sigma-Aldrich (St. Louis, MO, U.S.A).
The human bone marrow aspirates were obtained from normal children, Department of Pediatrics in Ajou University Medical Center (Suwon, Korea).
7 B. Methods
1. Common Methods 1.1. Animals
Transgenic mice harboring the hSOD1G93A (B6SJL-TgN(SOD1-G93A)1Gur) transgene
described by Gurney et al.65 were obtained from the Jackson Laboratories (Bar Harbor, ME). Hemizygotic transgenic progeny were obtained by mating transgenic males with F1 hybrid females obtained by crossing C57BL6 females with Swiss Jim Lambert (SJL) males. DNA was obtained from the tails of transgenic progeny, and the presence of the hSOD1 G93A transgene was confirmed by polymerase chain reaction (PCR) amplification using specific primers (forward, 5'-CAT CAG CCC TAA TCC ATC TGA -3' and reverse, 5'- CGC GAC TAA CAA TCA AAG TGA-3') and 35 cycles of amplification with an annealing temperature of 60°C. Food and water were available ad libitum, and animals were weighed weekly. All experimental procedures were examined and approved by the Institutional Animal Research Ethics Committee at the Ajou University Medical Center.
1.2. Evaluation of motor functions (Fig. 1.)
Extension reflexes were evaluated according to the procedure and scoring system described by Pascal Barneoud et al. (Barneoud 1997) and Patrick Weydt et al. (Weydt 2003). Briefly, mice were suspended by the tail, and the degree of motor deficit was scored from 0 to 2 as follows: a normal extension reflex in both hindlimbs was scored as 2; imbalanced extension in the hindlimbs as 1.5; extension reflex in only one hindlimb as 1.0; the absence of any hindlimb extension as 0.5; and total paralysis as 0.
8
Fig. 1. Evaluation of motor functions. Extension reflexe (A). Paw Grip Endurance (PaGE)
9
Basic motor function was measured using the paw grip endurance (PaGE) test, which requires only balance and grip strength (Weydt 2003). Briefly, we measured the time a given animal held on to the inverted lid of cage. Each mouse was given 3 trials and the longest latency was recorded. The cut-off time was 90 seconds.
More advanced motor functions were assessed using the rotarod test, which is a complex task requiring good motor coordination and balance in addition to strength (Sango 1996; Barneoud 1997; Karl 2003; Miana-Mena 2005). Briefly, we measured the time a given animal remained on the rotating cylinder (3.5 cm in a diameter) of a rotarod apparatus (Acceler Rota-Rod 7650, UGO BASILE, Varese, Italy) revolving at a constant speed of 16 rpm. Each animal was given 3 trials, and the longest latency before falling was used. The arbitrary cut-off time was 300 seconds.
1.3. Assessment of disease onset and lifespan
The onset of disease was defined as the time point at which the mouse could no longer remain on the rotarod apparatus for 300 sec at a speed of 16 rpm. Mortality was scored as the age of death or the age at which the mouse was unable to right itself within 30 sec after being placed on their sides.
1.4. Statistical analysis
Statistical analysis was carried out in SPSS version 15.0 (SPSS Inc., Chicago, IL). Data were analyzed by one-way analysis of variance (ANOVA) where appropriate. Significant differences were further evaluated using Tukey’s HSD post hoc test. All results are expressed as the mean ± SEM. Survival was analyzed with the Kaplan-Meier curve, and significance of
10
associations with log rank, Breslow and Tarone-Ware tests.
2. (Part1) Neural Induction with Neurogenin1 Prolongs Therapeutic Effects of Mesenchymal Stem Cells in an Amyotrophic Lateral Sclerosis Mouse Model.
2.1. MSCs-Ngn1 and in vitro differentiation
Human mesenchymal stem cells were isolated from bone marrow with approval of the Institutional Review Board of Ajou University Medical Center (Suwon, Korea) by plastic adherence as described previously(Kim 2005) and cultured in growth medium (Dulbecco’s modified Eagle’s medium [DMEM] supplemented with 10% fetal bovine serum [FBS] and 10 ng/ml basic fibroblast growth factor [bFGF]). MSCs-Ngn1 were previously described(Kim 2008).
For terminal differentiation into neuronal cells in vitro, MSCs-Ngn1 were grown on poly-D-lysine/collagen-coated coverslips in growth medium containing 10 µM 5-aza-deoxycytidine (Aza-dC; Sigma-Aldrich, St. Louis, http://www.sigmaaldrich.com) for 3 days and then in DMEM/F12 medium (Invitrogen, Carlsbad, CA, http://www.invitrogen.com) containing 10% FBS and 10 µM forskolin (Sigma-Aldrich) for 14 days. Neuronal differentiation was verified by immunocytochemistry using an anti-microtubule associated protein 2 (MAP2; Sigma-Aldrich, 1:500) and an anti-neurofilament 200 (NF200; Chemicon, Temecula, CA, http://www.chemicon.com, 1:500), as a neuronal marker.
2.2. Transplantation
11
genders. We injected 106 cells of either MSCs or MSCs-Ngn1 in 0.1 ml Phosphate Buffered Saline (PBS) into the lateral tail vein of the mice. Donor cells were infected with adenviral vectors expressing nuclear localization signal (NLS)-linked β-galactosidase (m.o.i.=100) before transplantation for in vivo identification. Experiments were carried out in three transplantation protocols (Table 1). In the first set of experiment, animals received the cells at 8 weeks before the onset of the ALS symptom. In the second experiment, the animals received the cells on the same week that animals first failed the paw grip endurance (PAGE) test between 13-16 weeks after birth. In the third experiment, animals received the cells twice at 13 and 15 weeks. The negative control group received PBS.
2.3. Histological analysis of transplanted cells
Two weeks after transplantation, animals were sacrificed under deep anesthesia and perfused transcardially with 0.9% saline and 2% paraformaldehyde in 0.1-M phosphate buffer (pH 7.4). The brains, spinal cords, livers, hearts, lungs, kidneys, spleens and muscles were trimmed in a 3-4 mm thickness, post-fixed in 2% paraformaldehyde for 12 h, and finally incubated at 37°C overnight in X-gal staining solution (1mg/ml X-gal, 5 mM potassium ferrocyanide, 5 mM potassium ferricyanide, 2 mM MgCl2, 0.02% nonidet P-40, 0.1 M phosphate buffer pH 7.4). Tissues were embedded in paraffin and serially sectioned in a 5 µm-thickness for light microscopical analysis.
Five µm thick spinal cord sections containing -galactosidase positive cells were immunostained with primary antibodies against human mitochondria (1:100), NeuN (1:500), CNPase [2',3'- cyclic nucleotide 3'-phosphodiesterase] (1:500), choline acetyltransferase [ChAT] (1:500) from Chemicon; glial fibrillary acidic protein [GFAP] (1:500) from
Sigma-12
Aldrich. Immunoreactivity was visualized using biotinylated goat anti-mouse secondary antibodies (1:200) [ABC kit, Vector Laboratories, Burlingame, CA, http://www. vectorlabs.com] for 1 h and for 30 min and then with 0.05% diaminobenzidine and 0.03% H2O2 in 50 mM Tris-HCl, pH 7.0. After mounting, sections were examined with a Zeiss Axiophot microscope (Zeiss, Jena, Germany, http://www.zeiss.de).
2.4. Quantification of motor neurons and Iba1+ microglia
At 16 weeks of age, the mice were sacrificed under deep anesthesia and perfused. The cervical and lumbar segments of the spinal cord were post-fixed with 2% paraformaldehyde in 0.1-M phosphate buffer (pH 7.4) and cryoprotected in 30% sucrose in 0.1M phosphate buffer (pH 7.4) overnight. Ten 30 µm-thickness sections with a 300 µm distance were obtained per cervical or lumbar spinal cord and stained with Nissl staining. Large cells with a diameter greater than 20 µm, a distinct nucleus, nucleoli, and at least one thick process in the ventral horn below a lateral line from the central canal were counted.
In order to estimate activated microglia, cervical spinal cord sections were incubated overnight at 4 °C with anti-Iba1 (Wako Chemicals USA. Inc., Richmond, VA, 1:1000), and antibody reactions were visualized using a ABC kit. Since microglia undergo morphological changes from ramified to amoeboid-like, we measured the total immunoreactivity. The ventral horn area occupied by Iba-1 immunoreactivity was obtained using Image J. 1.34vi software. Data from 5 mice per group were presented as the means S.E.
3. (Part2) Transplantation of Mesenchymal Stem Cells Transduced with GDNF Prolongs Lifespan and Ameliorates Motor Dysfunctions in an Amyotrophic Lateral
13 Sclerosis Mouse Model.
3.1. Western blot analysis for GDNF
The MSCs were incubated with the GDNF adenoviral supernatant (m.o.i.=100) in the growth medium for 2 hours. Cell lysate preparation was performed at 48 hours after transduction. For preparation of cell lysate, cells were washed three times with PBS and harvested in 1.5㎖ tube using cell scrapper. After centrifugation at 3000 rpm for 5 min, the resulting pellet was suspended in 50 mM Tris (pH 7.5), 150 mM NaCl, 2 mM EDTA, 1% Sodium deoxycholate, 0.1% sodium dodecyl sulfate (SDS), 1% Triton X-100 and protease inhibitors, then incubated for 10 min at room temperature. After centrifugation at 12,000 rpm for 3 min at 4 ℃, the supernatant was collected. 100 ㎍ of extract was subjected to 9 % SDS-PAGE. The proteins on the gel transferred electrophoretically onto the PVDF membrane. After the transfer, the membrane was blocked in 5 % skim milk, and incubated in anti-GDNF polyclonal antibody (1:1000; R&D systems, U.S.A). Finally, the membrane was incubated with horseradish peroxidase-conjugated anti-goat IgG antibody (1:5000; Pierce, Rochford, IL, U.S.A) and the proteins were visualized using an enhanced chemiluminescence (ECL) kit following the manufacture’s recommendation.
3.2. Immunocytochemistry
The MSCs-GDNF was fixed with 4% buffered paraformaldehyde solution for 10 min. The fixed was washed with PBS solution, three times and incubated with 10% normal goat serum in PBS for 1 hr. After sufficient washing, the cells were incubated with primary antibodies against GDNF (1:500; goat-polyclonal, R&D systems) for 3 hr at room
14
temperature. Following several washing with 0.1% Triton X-100 containing PBS solution, cells were incubated in Alexa488 conjugated anti-goat IgG (1:1000) secondary antibody for 1 hr in dark chamber. After sufficient washing with PBS, counter stained with Hoechst 33258, and cells were mounted on glass slides by using Vectashield (mounting medium for fluorescence, Vector). Immunoreactivity to GDNF was investigated on Fluorescence microscope with FITC filter.
3.3. Transplantation
The male mice were divided into three groups with the matched ages. We injected 106 cells of either MSCs or MSCs-GDNF cells in 0.1 ml PBS into the lateral tail vein of the mice. In order to identify the donor-derived, the cells were infected with an NLS-linked β-galactosidase-expressing adenovirus (m.o.i.=100) before transplantation. The animals received the cells on the same week that animals first failed in Paw grip endurance (PAGE) test between 13-16 weeks after birth. In the third experiment, animals received cells twice at 13 and 15 weeks. The negative control animals received PBS.
3.4. Motor neuron Counting
(See Methods 2.4)
4. (Part3) Effects of estrogen on lifespan and motor functions in female Amyotrophic Lateral Sclerosis Mouse Model.
15
Ovariectomies (OVX) were performed at postnatal day 50. Briefly, each female mouse was anesthetized with an intraperitoneal injection of 50 mg/kg ketamine and 2.5 mg/kg rumpun. The hair on the lumbar dorsum was shaved bilaterally, a 0.5-1 cm dorsal flank incision was used to penetrate the abdominal cavity, and the parovarian fatty tissue was retracted. The ovarian vasculature was ligated to maintain hemostasis, and the exposed ovaries were removed. The incision was sutured in layers and topical antibiotic was applied to the incision site. Sham operations were carried out in the same manner, except that ovaries were not removed. One week after OVX, estrogen treated group received an estrogen pellet containing 0.18 mg of 17β-estradiol (E2) (IRA, Sarasota, FL, USA) subcutaneously to deliver a continuous supply of estrogen for 60 days (Heikkinen 2002).
4.2. Measurement of E2 concentration and uterine weight
For determination of serum E2 concentrations and uterine weights, 5 animals from each experimental group were sacrificed at the age of 16 weeks. The uterus was removed from each animal and weighed. Blood samples were taken and serum (100 ul) was extracted with 1 ml ethyl ether. After the organic phase was vacuum dried, the pellet was dissolved in the extraction buffer supplied by the manufacturer and subjected to enzyme-linked immunosorbent assay (ELISA) using the Enzyme Immunoassay kit for estradiol (Oxford Biomedical Research Inc., Oxford, MI, USA).
16
III. RESULTS
A. (Part1) Neural Induction with Neurogenin1 Prolongs Therapeutic Effects of Mesenchymal Stem Cells in an Amyotrophic Lateral Sclerosis Mouse Model.
1. In vitro cultivation of MSCs-Ngn1
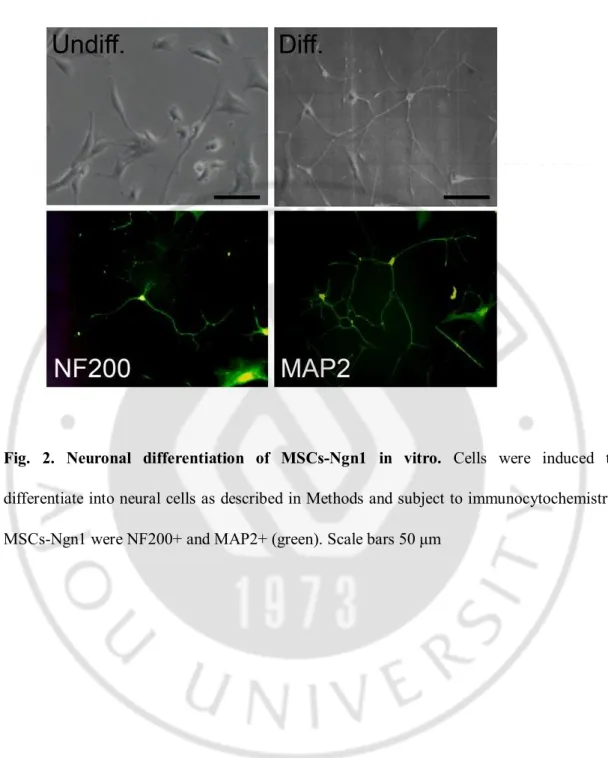
Neural induction of MSCs was performed as previously described (Kim 2008). Briefly, human bone marrow MSCs were stably transduced with retroviral vectors encoding Ngn1 to obtain MSCs-Ngn1. Consistent with our previous results, MSCs-Ngn1 retained the MSCs’s fibroblastic morphology when grown in the growth medium, but expressed neuronal phenotypes following terminal differentiation with Aza-dC and forskolin. MSCs-Ngn1 extended neurites and expressed neuron-specific proteins, including NF200 and MAP2 as shown by immunocytochemistry (Fig. 2). Under the same conditions, these proteins were not detected or very minimally expressed in MSCs (data not shown).
2. Distribution of Transplanted MSCs-Ngn1 in the Central Nervous System (CNS)
Since cell death of motor neurons occurs in a diffusive manner over the entire CNS in hSOD1G93A mice starting from 9 weeks (Martin 2007b), we systemically injected the cells
through the tail vein at 8 weeks of age before the animals exhibited ALS behavior. The cells were pre-labeled with adenoviral vectors encoding NLS-linked β-galactosidase for identification of the grafted cells. Two weeks after intravenous administration, we sacrifice the mice and stained the tissues for expression of LacZ gene by X-gal staining. The X-gal positive cells also were bearing human mitochondrial antigen, verifying that they were the
17
Fig. 2. Neuronal differentiation of MSCs-Ngn1 in vitro. Cells were induced to
differentiate into neural cells as described in Methods and subject to immunocytochemistry. MSCs-Ngn1 were NF200+ and MAP2+ (green). Scale bars 50 μm
18
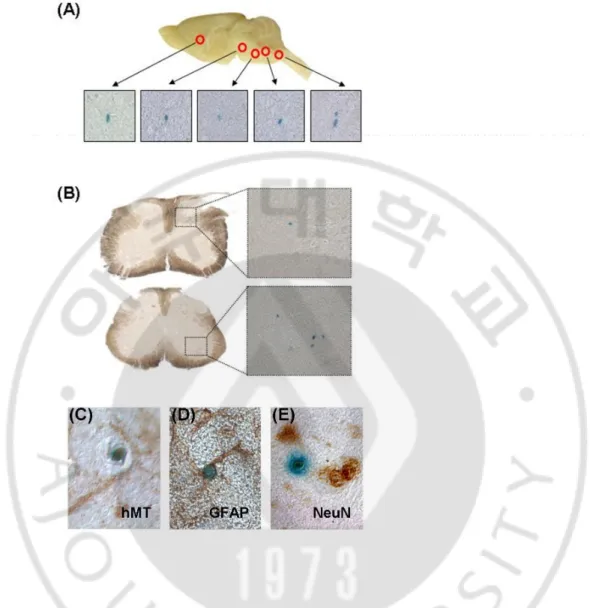
Fig. 3. Migration of transplanted MSCs-Ngn1 into the CNS and immunophenotypes of MSCs-Ngn1 within the CNS. The nuclear-LacZ expressing MSCs-Ngn1 were transplanted
into tail vein of the ALS mice (8 weeks of age). The brain and spinal cord were isolated after 2 weeks, and the tissues were stained by X-gal. The LacZ positive cells were detected in widespread area of the brain (A) and spinal cord (B). Every X-gal positive cells were stained with anti-human mitochondria (hMT) antibody (C). Few cells were double stained with LacZ+/GFAP+ (D) or LacZ+/NeuN+ (E). Most grafted cells did not express any specific neural markers.
19
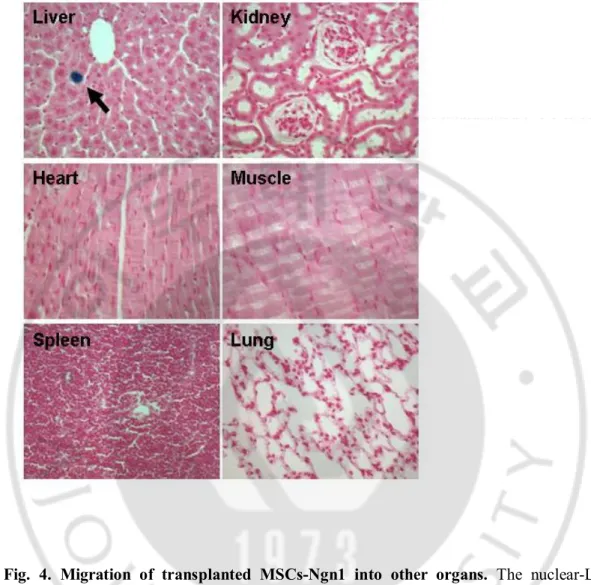
Fig. 4. Migration of transplanted MSCs-Ngn1 into other organs. The nuclear-LacZ
expressing MSCs-Ngn1 were transplanted into tail vein of the ALS mice (8 weeks of age). After intravenous administration of MSCs-Ngn1, the cells were not found in all peripheral organs except liver. In the liver, the MSCs-Ngn1 were present in the parenchyma.
20
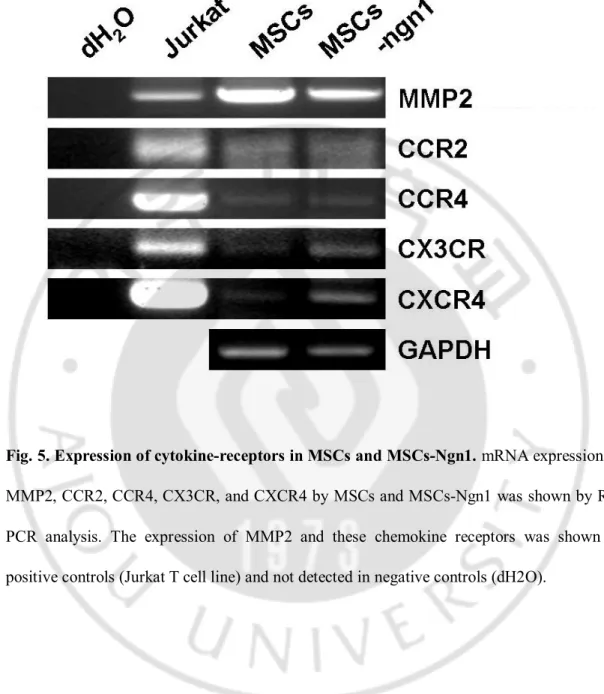
Fig. 5. Expression of cytokine-receptors in MSCs and MSCs-Ngn1. mRNA expression of
MMP2, CCR2, CCR4, CX3CR, and CXCR4 by MSCs and MSCs-Ngn1 was shown by RT-PCR analysis. The expression of MMP2 and these chemokine receptors was shown in positive controls (Jurkat T cell line) and not detected in negative controls (dH2O).
21
grafted human cells (Fig. 3. C). Systemic injection led to dispersion of the cells throughout the body, and X-gal positive cells were also found in liver but not in other peripheral organs such as spleen, lung, kidney, heart, and muscle (Fig. 4.). Thus, migration efficiency of the grafted cells to the CNS could not be practically quantified, although X-gal positive cells were easily detected throughout the entire CNS of animals with MSCs-Ngn1 but to a lesser degree in animals with MSCs (Fig. 3.). X-gal positive MSCs-Ngn1 mostly remained devoid of NeuN, a neuronal marker, or GFAP, an astrocyte maker (data not shown). However, very rarely we found X-gal positive cells that were also expressing NeuN or GFAP (Fig. 3. D, E).The results suggest that transplanted MSCs-Ngn1 have potential to migrate into the CNS regions efficiently where they remain presistently.
3. Protection of Spinal Motor Neurons
To determine the protective roles of the MSCs and MSCs-Ngn1, we transplanted the cells at 8 weeks before the onset of the ALS symptoms and compared the survival of spinal motor neurons with Nissl staining at 16 weeks of age (Fig. 6.). In hSOD1G93A mice, the cells with a cell body greater than 20 µm and a distinct nucleus in the ventral horn were expressing choline acetyl transferase (ChAT), indicating that they were motor neurons. Compared to the values of healthy wild type (WT) litter mates (cervical: 58±2.1; lumbar: 51±2.2), the number of motor neurons were reduced to 25% in the cervical region and to 15% in the lumbar region of PBS-injected hSOD1G93A control mice (cervical: 14±1.0;
lumbar: 8±1.2). Interestingly, transplantation of MSCs-Ngn1 prevented the loss of ventral motor neurons, thus 45% of the cervical and 25% of the lumbar motor neurons survived
22
Fig. 6. Counting of survived motor neuron. The MSCs and MSCs-Ngn1 were transplanted
into tail vein of the ALS mice (8 weeks of age) and the spinal cords were isolated at 16 weeks of age. For motor neuron counting, the spinal cord sections were processed by Nissl staining (A). Number of large ventral horn neurons was measured on the spinal cord sections (B). hSOD1G93A mice contained obvious degenerating motor neurons, but wildtype mice
showed no evidence of degeneration. The result showed that MSCs-Ngn1 improves motor neuron survival. Data are shown as means SEM (**p<0.01 both comparisons by Tukey's HSD).
23
Fig. 7. Effect of MSCs and MSCs-Ngn1 on the neuroinflammation. Pathological changes
following ALS disease progression trigger inflammatory responses. Thus, we tested for differences in inflammatory cells (Iba1+) at 16 weeks of age (A). The inflammatory index based on Iba1 immunoreactive area was significantly reduced to a similar degree after transplantation of MSCs and MSCs-Ngn1 (B). Data are shown as means SEM (**p<0.01 both comparisons by Tukey's HSD).
24
(cervical: 22±1.1, lumbar: 12.±1.1). In contrast, transplantation of parental MSCs did not prevent the motor neuron loss, and the numbers of motor neurons were similar to those found in the PBS control (cervical: 13±0.7, lumbar: 7±0.7).
4. Immune modulation by transplanted cells
Since chronic inflammation is suggested as a common factor associated with ALS, we assessed microglia activation, a hallmark of brain inflammation. Since microglia radically transform their morphology from ramified to amoeboid types when activated(Davis 1994; Streit 2000), it is difficult to distinguish between active and resting microglia when counting the number of Iba1-positive microglia in the spinal cord, thus, we measured the total Iba-1immunoreactivity in the spinal cord. The Iba-1 immunoreactivity was minimal in the healthy WT control (5±0.6), but enhanced by 5.3-fold in the control hSOD1G93A mice with
PBS (26±3.1) (Fig. 7. A, B), indicating that inflammation was in progress in hSOD1G93A mice. Systemic injection of MSCs or MSCs-Ngn1 similarly suppressed the Iba-1 immunoreactivity (MSCs-Ngn1: 11±2.1; MSCs, 12±2.4) (Fig. 7. B). The data indicate that MSCs were indistinguishable with and without Ngn1 in reducing the inflammatory responses in the ALS model.
5. Delayed disease progression by MSCs-Ngn1
To determine whether survival rates of motor neurons correlated with behavior improvement, we performed behavior studies with three experimental paradigms (Table. 1.). Before transplanting the cells, we measured the weights of hSOD1G93A mice and the baseline
25
equally grouping the animals. For Experiment I, we systemically injected the cells at the age of 8 weeks into the tail vein and monitored animals for following 12 weeks until they were declared to be dead (Fig. 8. A). Early transplantation of MSCs-Ngn1 before motor neurons started to degenerate delayed the average onset of disease by 5 days from 16.1±0.5 to 16.8±0.3 weeks and extended the average lifespan from 127.4±3.4 days to 130.5±4.6 days (Fig. 8. B, C). MSCs-Ngn1-treated group performed extension reflex, rotarod, and PaGE tests between 15-17 weeks better than the PBS-treated control group (compare filled and white bars in Fig. 8. D-F; Experiment I in Table. 1.). In contrast, MSCs did not alter the onset ages or the lifepan, but only improved the scores in the extension reflex test (Fig. 8. D). The results indicate that early transplantation of MSCs-Ngn1 may be moderately effective to delay disease onset and improve the motor functions.
Next, we investigated the influence of transplantation timing in disease progress. We transplanted the cells right after the animals exhibited first symptom of disease in the PaGE test between 14-16 weeks. Transplantation of MSCs-Ngn1 increased the average lifespan by 7 days, from 127.9±2.9 to 135.3 ± 1.9 days. Moreover, the MSCs-Ngn1-treated mice showed marvelous performance in all tests between 15-17 weeks (filled bars in Fig. 9. C-E; Experiment II in Table. 1.). The MSCs-treated group performed rotarod test better than PBS-treated group at 16 weeks (MSCs, 203.6±35.9; PBS, 116.0±45.2 sec), although they had similar lifespan compared to the PBS-treated group (Fig. 9. B). Interestingly, the beneficial effects of MSCs did not last till 17 weeks as shown in rotarod and PaGE tests (PaGE: PBS, 30.4±11.0 and MSCs 32.5±9.3 sec; rotarod: PBS, 56.9±28.3 and MSCs 54.8±17.4 sec). The results indicate that neural induction with Ngn1 increases the paracrine functions of MSCs to extend the lifespan and delay the disease progression. The results also indicate that the
26
Fig. 8 Onset of disease, total survival time and characteristics of disease progression in MSCs-Ngn1 transplanted ALS mice (treatment in the pre-onset stage). Details of the
Experimental Protocol (A), cumulative probability of onset of rotarod deficits in ALS mice (B), and survival of ALS mice after transplant (C). Evaluation of the Extension Reflex in the different experimental groups (D), motor coordination of experimental groups measured using the Rotarod test (E), and muscle strength measured using the Paw Grip Endurance test (F). Data are shown as means SEM.
27
Fig. 9 Total survival time and characteristics of disease progression in MSCs-Ngn1 transplanted ALS mice (treatment in the disease-onset stage). Details of the Experimental
Protocol (A), and survival of ALS mice after transplant (B). Evaluation of the Extension Reflex in the different experimental groups (C), motor coordination of experimental groups measured using the Rotarod test (D), and muscle strength measured using the Paw Grip Endurance test (E). Data are shown as means SEM (**p<0.01 both comparisons by Tukey's HSD).
28
therapeutic effects of MSCs-Ngn1 are higher when the cells are transplanted at disease-onset ages than pre-onset ages.
6. Enhanced therapeutic effects of MSCs-Ngn1 by repeated grafting
To test whether short-lived effects of MSCs can be extended by multiple treatments, we transplanted cells two times with a two weeks interval; first at 13 weeks of age near the disease onset and second at 15 weeks (Fig. 10.). This time, all animals were immunosuppressed with cyclosporine A (CsA, 4 mg/kg I.P every other day) in order to eliminate possible immune rejection possibly evoked by repeated grafting (Yan 2006). Treatment with CsA alone did not influence the disease progression so that the survival rates and behavioral scores were not significantly different between the CsA- and vehicle-treated animals (Fig. 11.). Importantly, when injected twice, MSCs were as effective as MSCs-Ngn1 in rotarod test at 15 weeks (Fig. 10. D), as well as PaGE and extension reflex tests at 16 weeks (Fig. 10. C, E), although the difference between MSCs- and PBS-treated animals was not statistically significant. Consistently, multiple treatment of MSCs-Ngn1 significantly extended the average lifespan by 9 days compared with the PBS-control from 125.1±2.8 to 133.9±2.1 days. During the symptomatic period between 15 and 16 weeks, the scores from extension reflex, rotarod, and PaGE tests were significantly higher in the animals treated twice with MSCs-Ngn1 than in the PBS-treated control. Compared to the single treatment in Experiment II, the difference between the MSCs-Ngn1 and PBS-treated control animals was statistically significant (p < 0.01). Our results suggest that first treatment of MSCs-Ngn1 first at 13 weeks delay the disease onset and the second treatment secure the improvement of behavioral performance during symptomatic period
29
Fig. 10. Total survival time and characteristics of disease progression in MSCs-Ngn1 transplanted ALS mice (treatment in the disease-onset stage, 2 times). Details of the
Experimental Protocol (A), and survival of ALS mice after transplant (B). Evaluation of the Extension Reflex in the different experimental groups (C), motor coordination of experimental groups measured using the Rotarod test (D), and muscle strength measured using the Paw Grip Endurance test (E). Data are shown as means SEM (*p<0.05, **p<0.01 both comparisons by Tukey's HSD).
30
Fig. 11. Total survival time and characteristics of disease progression in CsA injected ALS mice. No statistically significant differences between CsA-treated and vehicle-treated
animals under certain condition. Details of the Experimental Protocol (A), and survival of ALS mice (B). Evaluation of the Extension Reflex (C), the Rotarod test (D), and the Paw Grip Endurance test (E). Data are shown as means SEM.
31
Table. 1. Experimental protocols and summary results (MSCs-Ngn1).
Experiment I (Fig.8) II (Fig.9) III (Fig.10)
Group PBS MSC-Ngn1 MSC PBS MSC-Ngn1 MSC PBS MSC-Ngn1 MSC
No. of animals M:4, F:4 M:4, F:4 M:4, F:4 M:4, F:4 M:4, F:4 M:4, F:4 M:6, F:6 M:6, F:6 M:6, F:6 Transplantation
(Days) 60 109.0±2.2 109.3±4.1 109.3±1.9 90, 105 (twice)
Immune
suppressant - - - CsA (4mg/kg, I.P every other day)
Onset time (Weeks) 16.1±0.5 16.8±0.3 16.1±0.2 15.6±0.3 15.6±0.6 15.6±0.3 15.9±0.3 16.6±0.3 16.6±0.2 Lifespan (Days) 127.4±3.4 130.5±4.6 128.5±3.5 127.9±2.9 135.3±1.9 127.3±3.4 125.1±2.8 133.9±2.1** 129.7±2.1 Ex-Reflex-15W (Point) 1.31±0.11 1.53±0.15 1.56±0.08 1.50±0.11 1.75±0.08 1.63±0.13 1.63±0.10 1.90±0.06* 1.85±0.06 Rotarod-16W (Sec) 143.3±45.6 169.8±42.5150.8±34.7 116.0±45.3 277.8±11.1** 203.6±35.9 111.5±34.0 224.8±27.7* 203.2±27.1 PaGE-17W (Sec) 32±9.1 47.9±11.4 33.5±10.4 30.4±11.0 47.1±9.6 32.5±9.3 26.8±10.0 64.8±9.7* 58.5±10.5
32
33
34
35
B. (Part2) Transplantation of Mesenchymal Stem Cells Transduced with GDNF Prolongs Lifespan and Ameliorates Motor Dysfunctions in an Amyotrophic Lateral Sclerosis Mouse Model.
1. Evaluation of cytotoxicity and expression efficiency of adeno-GDNF virus
To optimize the efficiency of adenoviral vector-mediated gene expression, MSCs were infected with the adeno-GDNF virus at different MOIs. After exposed to various concentrations of viruses, the cytotoxicity of virus vector was determined using immunocytochemistry (Fig. 12. A). The result showed that the cytotoxicity of adenoviral vector was dose-dependent. GDNF protein expression was detected by western blot (Fig. 12. B). The highest levels of GDNF expression in MSCs were observed at an MOI of 100 and 1000. If the MOI is higher than 100, adenovirus can induce the cell death directly. If the MOI is lower than 100, GDNF expression was too low. From these results, we decided to use an MOI of 100 for the experiment.
MSCs were infected with the recombinant adenovirus GDNF, MSCs-GDNF, high levels of GDNF expression were observed in MSCs-GDNF by immunocytochemistry (Fig. 13. A) and western analysis (Fig. 13. B). The nuclear-LacZ expressing MSCs-GDNF were transplanted into tail vein of the hSOD1G93A mice (8 weeks of age). After two weeks, many
gal positive cells were distributed in the brain and spinal cord (data not shown). Every X-gal positive cells were stained with anti-GDNF antibody (Fig.13. C).
36
Fig. 12. Safety and transduction efficiency test of recombinant adeno-GDNF virus. To
test the safery and efficiency of adeno-GDNF virus, MSCs were transduced at sirial multiplicity of infection (MOI). After 2 days, we detected the GDNF expression by Immunocytochemistry (A) and Western blot (B).
37
Fig. 13. Confirmation of GDNF expression in vitro and in vivo. The GDNF expression of
MSCs-GDNF was evaluated by Western blot (A) and Immunocytochemistry (B). The nuclear-LacZ expressing MSCs-GDNF were transplanted into tail vein. Spinal cord were isolated after 2 weeks, and the tissues were stained by X-gal. The LacZ positive cells were were stained with anti-GDNF (C).
38
2. Effect of MSCs-GDNF administration on motor neuron survival and disease progression
The MSCs, MSCs-GDNF and PBS were transplanted into tail vein of the ALS male mice two times (first: 13 weeks of age, second: 15 weeks of age). All animals were immunosuppressed with 4mg/kg cyclosporine every 2 days for the duration of the study. To assess the neuroprotective effect of MSCs-GDNF on the survival of motor neurons, we compared the numbers of spinal motor neurons in the different groups at 16 weeks of age. In the cresyl violet staining (Fig. 14. A), motor neurons in cervical and lumbar spinal cord were significantly preserved in MSCs-GDNF treated group than in PBS treated groups (cervical: PBS, 13.5±0.52; MSCs-GDNF, 15.2±0.50, lumbar: PBS, 8.8±0.43; MSCs-GDNF, 9.9±0.38, p<0.05).
Multiple MSCs-GDNF treatment significantly expended the lifespan by an average of 9 days compared with PBS injected animals (PBS group 118.5±1.67 days; MSCs-GDNF group 126.9±2.21 days; MSCs group 125.9±3.56 days) (Fig. 15. A). The delay in the motor impairment – assessed by the Extenstion Reflex, Rotarod, PaGE test – was significant in mice treated with MSCs-GDNF compared with the control animals (Fig. 15.).
In PBS treated mice, extension reflex began to decrease at 14 weeks and reached to 1.7±0.09 point at 15 weeks and 0.6±0.13 point at 17 weeks. In MSCs-GDNF treated mice, extension reflex began to decrease at 15 weeks and progressed at slower rate than in control mice. Thus, the extension reflex was 1.9±0.07 point at 15 weeks and 1.1±0.12 point at 17 weeks (p<0.05) (Fig. 15. B). During symptomatic period (16 weeks of age), MSCs-GDNF mice showed significantly higher scores (P<0.05) than control in the rotarod test (Fig. 15. D).
39
Fig. 14. Counting of survived motor neuron. The MSCs-GDNF were transplanted into tail
vein of the ALS male mice (13, 15 weeks of age) and the spinal cords were isolated at 16 weeks of age. For motor neuron counting, the spinal cord sections were processed by Nissl staining (A). Number of large ventral horn neurons was measured on the spinal cord sections (B). The result showed that MSCs-GDNF improves motor neuron survival. Data are shown as means SEM (*p<0.05 both comparisons by Student’s t-test).
40
Fig. 15. Total survival time and characteristics of disease progression in MSCs-GDNF transplanted ALS mice (treatment in the disease-onset stage, 2 times). Survival of ALS
mice after transplant (A). Evaluation of the Extension Reflex in the different experimental groups (B), and muscle strength measured using the Paw Grip Endurance test (C). Motor coordination of experimental groups measured using the Rotarod test (D), and Data are shown as means SEM (*p<0.05, **p<0.01 both comparisons by Tukey's HSD).
41 Table. 2. Summary results (MSCs-GDNF).
Latency of fall (sec) Survival (days)
15 week 16 week 17 week
Ex-reflex 1.7 ± 0.09 1.3 ± 0.18 0.6 ± 0.13 Rotarod 265 ± 17.6 83 ± 40.6 16 ± 14.7 PBC (n=7) 118 ± 1.67 PaGE 86 ± 04.0 64 ± 12.4 17 ± 09.6 Ex-reflex 1.9 ± 0.07 1.6 ± 0.09 1.1 ± 0.12* Rotarod 300 ± 00.0 241 ± 36.5* 117 ± 44.3 MSCs-GDNF (n=7) 127 ± 2.21 PaGE 90 ± 00.4 85 ± 05.5 67 ± 10.5** Ex-reflex 1.8 ± 0.14 1.6 ± 0.15 0.9 ± 0.19 Rotarod 263 ± 36.7 206 ± 43.9 76 ± 39.3 MSCs (n=7) 126 ± 3.56 PaGE 89 ± 00.8 72 ± 10.4 58 ± 15.3*
42
The scores from the PaGE test seemed to be higher in MSCs-GDNF group than control group at 17 weeks of age (PBS 17.2±9.67 sec; MSCs-GDNF 66.7±10.49 sec, p<0.01) (Fig. 15. C). Interestingly, the MSCs were also significantly effective at 17 weeks (MSCs 57.9±15.31, p<0.05, vs PBS group) although less potent than MSCs-GDNF. The beneficial effects of MSCs may be through synergistic effect of repeated injection and immunosuppressant. Our results demonstrate that repeated injection of MSCs-GDNF at disease onset period showed remarkable efficacy.
C. (Part3) Effects of estrogen on lifespan and motor functions in female Amyotrophic Lateral Sclerosis Mouse Model.
1. Gender differences in onset and disease progression
To determine gender differences in the hSOD1G93A transgenic mice, we compared male and female hSOD1G93A mice with respect to disease onset ages, motor functions, and lifespan.
The overall population began displaying symptoms around 14.5 weeks; the mean onset age was 16 weeks for male hSOD1G93A mice, whereas it was significantly delayed to 16.5 weeks
in female mice (p<0.05) (Fig. 16. A). Following onset of symptoms, male mice began losing weight significantly at 15 weeks (p<0.05), whereas female mice maintained their body weights until 16 weeks of age (Fig. 16. B). The mean lifespan was 10 days longer in female mice; 125±1.3 days and 135±1.3 days in male and female mice, respectively (Fig. 16. C). Consistent with these observations, motor functions began to decline earlier in males than females. In male mice, extension reflex began to decrease at 14 weeks and reached to 1.49 point at 15 weeks and 0.75 point at 17 weeks. In female mice, extension reflex began to
43
Fig. 16. Gender differences in onset of ALS symptoms and eventual lifespan. Rotarod
tests were carried out twice a week with male and female hSOD1G93A mice (n = 35 per
group) to determine the disease onset ages (A), the average body weights (B), and the clinical endpoints (C). Data are shown as mean ± SEM. (Student’s t-test :* p<0.05; # p<0.01).
44
Fig. 17. Gender differences in motor functions. Male and female hSOD1G93A mice (n = 35
per group) were tested for extension reflexes (A) and rotarod tests (B), and PaGE tests (C). Female mice 14-17 weeks of age showed higher scores in all three tests compared to age-matched males. Data are shown as means ± SEM (Student’s t-test:* p<0.05, # p<0.01).
45
Fig. 18. Verification of ovariectomy and E2 replacement. Ovariectomy was performed at
postnatal day 50. (A, B) At 16 weeks of postnatal ages, the uterine weights were compared from five females per group. In sham-operated animals, the uterus without ovaries was used. (scale bar, 1 cm). (C) The concentration of E2 in serum was measured from the same groups. Data are shown as means ± SEM (#p<0.01; both comparisons by Tukey’s HSD).
46
decrease at 15 weeks and progressed at slower rate than in male mice. Thus, the extension reflex was 1.69 point at 15 weeks and 1.37 point at 17 weeks (Fig. 17. A). During this symptomatic period between 14 and 17 weeks, female mice showed significantly higher scores than age-matched males in the rotarod test (Fig. 17. B). The scores from the PaGE test seemed to be higher in female than male mice, although the difference between two genders was not statistically significant and omitted in the subsequent experiments (Fig. 17. C). These results indicate that the onset and progression of ALS symptoms are delayed in female hSOD1G93A mice compared to males. (Table. 2.)
2. Effects of female hormones in female hSOD1G93A mice
To examine whether the observed gender differences were due to female hormones, we removed the ovaries of female mice at postnatal day 50 and examined symptoms in mice with and without estrogen replacement. To confirm the effects of ovariectomy and estrogen replacement, ELISA was used to determine serum E2 levels, and the weight of uterus was measured at 16 weeks (Fig. 18.). As shown in Figure. 18. C, ovariectomized mice (OVX) showed lower circulating levels of E2 than sham-operated mice (16.8±6.9 pg/ml versus 67.2±12.4 pg/ml, respectively). The mice with E2-pellets (OVX+E2) showed higher E2 levels than sham-operated mice (OVX+E2; 136.8±31.3 pg/ml). As expected, ovariectomy caused uterine atrophy (Fig. 18. A, B); OVX mice had dried, closed vaginas, and a low mean of uterine weight (11.3±1.0 mg), whereas the OVX+E2 mice had swollen, moist vaginal tissues and openings, as well as the uterine weights comparable to those in the sham-operated group (131.3±14.9 mg versus 115.1±12.7mg, respectively). These results verified that ovariectomy successfully lowered serum E2 concentrations, which could be replaced by
47
Fig. 19. Protective effects of estrogen in the ovariectomized mice. (A) The onset ages of
the OVX (n=12), OVX+E2 (n=13), and sham-operated (n=10) groups were not significantly different. Body weights (B) and lifespan (C) were significantly reduced in the OVX group compared to OVX+E2 and sham-operated groups. Data are shown as means ± SEM (*p<0.05; OVX vs OVX+E2 by Tukey’s HSD).
48
Fig. 20. Beneficial effects of estrogen on motor functions. The decline of extension reflex
was significantly accelerated in the OVX group compared to the OVX+E2 group (A). Rotarod test did not show significant differences between the groups. Data are shown as mean ± SEM (*p<0.05, #p<0.01; OVX versus OVX+E2 by Tukey’s HSD).
49
Table. 3. Summary of gender differences and E2 effects.
Group Onset (weeks) Lifespan (days) n Male 16.0 ± 0.2 125 ± 1.3 35 Female 16.5 ± 0.2 135 ± 1.3 35 Sham 16.8 ± 0.4 136 ± 2.7 10 OVX 17.1 ± 0.3 129 ± 1.5 12 OVX + E2 17.1 ± 0.3 137 ± 2.0 13
50
implantation of E2-pellets.
Then the motor symptoms of the ovariectomized mice were examined with and without E2 treatment. E2 treatment did not affect the onset of clinical symptoms in hSOD1G93A
female mice (Fig. 19. A). Females in all groups first exhibited onset at around 14 weeks, with mean onset ages of 16.8±0.4, 17.1±0.3 and 17.1±0.3 weeks in the sham, OVX, and OVX+E2 groups, respectively. Importantly, the mean time to reach the clinical endpoint was 129±1.5 days in the OVX group, as compared to 137±2.0 days in the OVX+E2 group and 136±2.7 days in the sham-operated group (p<0.05; Fig. 19. C). Additionally, body weights were significantly reduced in the OVX group at 17 weeks, whereas there were no differences in body weight between mice in the OVX+E2 and sham-operated groups (Fig. 19. B).
Evaluation of motor functions such as simple motor function revealed that the decrease of extension reflex was significantly accelerated in the OVX group compared to the OVX+E2 and sham-operated groups (Fig. 20. A). However, more advanced motor functions as tested rotarod test did not show statistically significant differences between the groups (Fig. 20. B). Collectively, these results indicate that ovariectomy accelerates progression of ALS symptoms in female hSOD1G93A mice and thereby reduces the survival rates, which can
51
IV. DISCUSSION
A. (Part1) Neural Induction with Neurogenin1 Prolongs Therapeutic Effects of Mesenchymal Stem Cells in an Amyotrophic Lateral Sclerosis Mouse Model.
This study shows that transplantation of MSCs in the systemic circulation can delay motor impairment in the ALS animal model for a limited period of time between 15-16 weeks, and the short-lived beneficial effects of MSCs can be prolonged by neural induction with Ngn1.
Recently MSCs emerge as therapeutic candidates for the treatment of a variety of diseases including myocardial infarction (Barbash 2003; Bittira 2003; Gershlick 2001; Hahn 2008; Olivares 2004), skeletal muscle injury (Natsu 2004), brain injury,(Garbuzova-Davis 2003; Mahmood 2004; Mahmood 2003; Shichinohe 2004; Zhao 2002), neurodegenerative diseases (Dezawa 2004; Li 2001), spinal cord injury (Akiyama 2002; Hofstetter 2002; Ohta 2004), and peripheral nerve injury (Dezawa 2001; Tohill 2004). The therapeutic functions of MSCs are mostly derived from their strong tropism toward the lesion sites (Bakshi 2006; Deng 2006; Hofstetter 2002; Hou 2003; Ji 2004; Lu 2001; Mahmood 2004; Nakamizo 2005; Satake 2004; Wang 2002b), where they secrete neurotrophic factors including BDNF, GDNF, CNTF, bFGF, VEGF, HGF and IGF (Kurozumi 2005; Nagai 2007; Wieczorek 2003) and modulate immune responses in pathological conditions (Rasmusson 2006; Uccelli 2006). We previously showed that MSCs improved the recovery through those paracrine functions in the animal stroke model. Intracranial transplantation of MSCs reduced the neuronal cell death in the host brain, promoted endogenous neurogenesis, and suppressed inflammatory
52
responses(Yoo 2008). In the same model, we also showed that therapeutic effects of MSCs cells could be enhanced by neural induction of the cells with Ngn1(Kim 2008). MSCs-Ngn1 transdifferentiated into neurons and integrated to the host neural circuits while maintaining the original paracrine functions of MSCs(Yoo 2008). However, although MSCs-Ngn1 improved the behavioral performance in the ALS model, we rarely found neuronal cells that were transdifferentiated from them (Fig. 3.), suggesting that MSCs-Ngn1 differentially react to diverse pathological host environments between progressive diseases such as ALS and acute brain injury such as stroke(Park 2006). Nevertheless, MSCs-Ngn1 could integrate into CNS environment of the ALS animals with higher efficiency than MSCs (Fig. 3.), protect spinal motor neurons (Fig. 6.), and suppress inflammatory reaction (Fig. 7.). As a result, MSCs-Ngn1 delayed motor impairment more efficiently than MSCs (Fig. 9-10.) and lifespan in hSOD1G93A transgenic mice in which degeneration of motor neurons and subsequent death
of the animals are inevitable.
Degeneration of motor neurons occurs in a diffusive manner in spinal cord, motor cortex, and brainstem nuclei of ALS patients, thus, direct application of the therapeutic cells into the spinal cord may be insufficient to block disease progression (Mazzini 2004; Mazzini 2003; Mazzini 2006). Our study shows that application into the systemic circulation can effectively deliver the neural cells such as MSCs-Ngn1 in the wide range of CNS (Fig. 3.). Tropism of MSCs and MSCs-Ngn1 to the CNS might be partly attributed to enhanced immune reactions in the region where motor neurons degenerate. It has been reported that numerous reactive astrocytes and infiltrated T lymphocytes in the CNS of ALS patients or animals(Alexianu 2001; Hall 1998; McGeer 2002) secrete cytokines and chemokines including SDF-1, MCP-1, MIP-1 and MIP-2 (Chen 2004; Elliott 2001). Particularly, MCP1