저작자표시-비영리-동일조건변경허락 2.0 대한민국 이용자는 아래의 조건을 따르는 경우에 한하여 자유롭게 l 이 저작물을 복제, 배포, 전송, 전시, 공연 및 방송할 수 있습니다. l 이차적 저작물을 작성할 수 있습니다. 다음과 같은 조건을 따라야 합니다: l 귀하는, 이 저작물의 재이용이나 배포의 경우, 이 저작물에 적용된 이용허락조건 을 명확하게 나타내어야 합니다. l 저작권자로부터 별도의 허가를 받으면 이러한 조건들은 적용되지 않습니다. 저작권법에 따른 이용자의 권리는 위의 내용에 의하여 영향을 받지 않습니다. 이것은 이용허락규약(Legal Code)을 이해하기 쉽게 요약한 것입니다. Disclaimer 저작자표시. 귀하는 원저작자를 표시하여야 합니다. 비영리. 귀하는 이 저작물을 영리 목적으로 이용할 수 없습니다. 동일조건변경허락. 귀하가 이 저작물을 개작, 변형 또는 가공했을 경우 에는, 이 저작물과 동일한 이용허락조건하에서만 배포할 수 있습니다.
The effects of a 1.8 GHz continuous
electromagnetic fields on
mucociliary transport of human
nasal mucosa
by
Seung Min In
Major in Medicine
Department of Medical Sciences
The Graduate School, Ajou
The effects of a 1.8 GHz continuous electromagnetic
fields on mucociliary transport of human nasal
mucosa
by
Seung Min In
A Dissertation Submitted to The Graduate School of
Ajou University in Partial Fulfillment of the Requirements
for the Degree of Ph.D.
in Medicine
Supervised by
Hyun Jun Kim, M.D., Ph.D.
Major in Medicine
Department of Medical Sciences
The Graduate School, Ajou University
This certifies that the dissertation
of Seung Min In is approved.
SUPERVISORY COMMITTEE
Keehyun Park
Yun-Hoon Choung
Hyun Jun Kim
Chul- Ho Kim
Yoon- myoung Gimm
The Graduate School, Ajou University
December, 14, 2012
i
-ABSTRACT-
The effects of a 1.8 GHz continuous electromagnetic fields on mucociliary
transport of human nasal mucosa
The aim of this study was to investigate the effects of a 1.8 GHz continuous electromagnetic fields (EMF) on human nasal mucociliary transport and to determine the pathophysiology of ciliary beat frequency (CBF) during an EMF-induced change. Human nasal mucosa cells were exposed to a 1.8 GHz EMF (SAR= 1.0 W/kg), and CBF was analyzed using an optical flow technique with the peak detection method. The 1.8 GHz EMF-exposed group showed a decreased CBF when compared to the control group. In the cytotoxicity assay, the difference in survival rates was not found between the two groups. In the EMF-exposed group, protein kinase C (PKC) activity was increased during a PKC activity assay. The broad PKC inhibitor, Calphostin C abolished the EMF-induced decrease of CBF. The EMF-induced decrease of CBF was abolished by GF 109203X, a novel PKC (nPKC) isoform inhibitor, whereas the decrease was not attenuated by Gö-6976, a specific inhibitor of conventional PKC (cPKC) isoform. EMF may inhibit CBF via an nPKC-dependent mechanism. Therefore, we have confirmed that EMF could decrease CBF by increasing PKC activity.
Key word : Electromagnetic fields, Ciliary motility, Mucociliary transport, Protein kinase C, Mobile
ii
TABLE OF CONTENTS
ABSTRACT ……….……..………..i
TABLE OF CONTENTS ………..………….………..ii
LIST OF FIGURES ………..……….………..iii
I. I N T R O D U C T I O N … … … . . 1
II. M AT ER I AL AN D M ET H O DS …… ………… …… …… ……… …… …… … 3
1. Nor ma l huma n na sa l mucos a cult ur e ……….. 3
2. N H N E c e l l c u l t u r e … … … …… …… … … 3
3. Exp os ur e t o EM F ……… ……… ………… ………… ………… ……… …4
4. M T T a s s a y … … … . 6
5. Measur ement of CBF ……….6
6. The PKC kinase activity assay ………..7
7. Elucidation of the role of PKC in EMF-induced decrease of CBF ………8
8. Determination of PKC isoforms involved in the EMF-induced decrease of CBF……8
9. T he m ea s u r e m e n t of r ea l t i m e t e m p er a t u r e … … … …… . 9
10. The effect of 0.2 ℃ temperature difference ………9
11. The measurement of pH for 24 hours ………9
12. The measurement of intracellular calcium concentration ………10
13. Transmission electron microscope ……….10
14. Statistical analysis ……….11
III. RESULTS 1. The CBF response after a 1.8 GHz EMF exposure ……….12
2. MTT assay ………..……….13
3. PKC activity assay ………..…..………14
4. The role of PKC in the EMF-induced decrease of CBF …………..…………15
5. Det er minat ion of PKC subt ypes ………..16
6. The measurement of real time temperature ………18
7. The effect of 0.2 ℃ temperature difference ………19
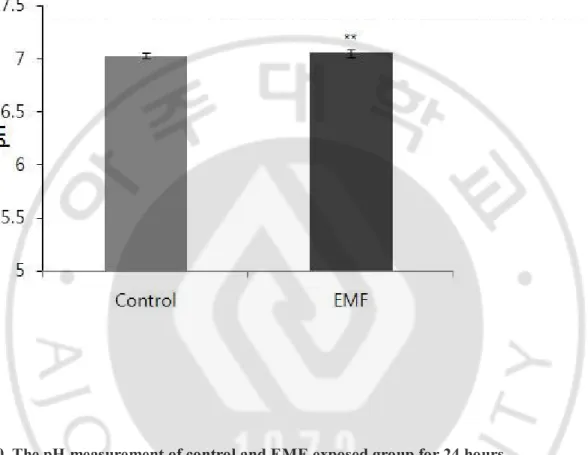
8. The measurement of pH for 24 hours ………20
9. The measurement of intracellular calcium concentration ………21
10. Trans miss ion electron micros cope ………22 IV. D I S C U S S I O N … … … 2 3 V. C O N C L US I O N … … … 2 9
iii
R E F E R E N C E S … … … . 3 0 국 문 요 약 … … … . . 3 4
iv
LIST OF FIGURES
Fig. 1. The schema of 1.8 GHz exposure device set up ………5
Fig. 2. The 1.8 GHz EMF-induced responses on CBF ………12
Fig. 3. MTT assay ………13
Fig. 4. PKC activity assay ………14
Fig. 5. The effect of Calphostin C on a 1.8 GHz EMF-induced response on CBF ………15
Fig. 6. The effect of Gö-6976 on a 1.8 GHz EMF-induced response on CBF ………16
Fig. 7. The effect of GF 109203X on a 1.8 GHz EMF-induced response on CBF …………17
Fig. 8. The real time temperature measurement of control and EMF-exposed group ………..18
Fig. 9. The effect of 0.2 ℃ on CBF without EMF exposure ………..19
Fig. 10. The pH measurement of control and EMF-exposed group ……….20
Fig. 11. The measurement of intracellular calcium concentration ………21
- 1 -
I.
INTRODUCTION
Cilia are omnipresent in the human body. They can be found in the upper and lower respiratory tract, the olfactory tracts, the vestibular-cochlear organ, the urogenital tracts, and even in the central nervous system. The main function of the cilia in respiratory tract is to eliminate inhaled particles. This is accomplished by a fast and synchronous beating pattern that transports the debris in a layer of mucus, much like a conveyor belt. This mucociliary transport (MCT) is a major respiratory tract host defense mechanism that works against inhaled microbes and noxious materials. The amounts and contents of mucus and periciliary fluid, in addition to cilia movement are essential for this fundamental defense mechanism. Cilia beat like automatic “sweeping broom” with its pivotal axis located at mucosal surface, therefore, plays dynamic role in this MCT system. Ciliary beat frequency (CBF) is the quantitative measurement of ciliary beats and the most important factor responsible for regulating mucociliary transport (Braveman et al., 1998). Impaired CBF leads to mucus secretion accumulation and allows bacteria or inhaled substances to invade the mucosa, leading to various respiratory tract disease entities, such as rhinosinusitis and bronchitis. The human respiratory system is continuously exposed to various environments such as bacteria, viruses, fungi, cigarette smoke, and air pollution. In modern urban life, electromagnetic fields (EMF) might be one of the common environments, and we are exposed to a great amount of EMF from electronic equipment. Especially, the mobile phones and other wireless gadgets are held close to the body and are used frequently with exposure duration around 1 to 2 hours per day, these devices are potentially the most dangerous sources of electromagnetic radiation that the average person possesses. Therefore, mobile phone frequency band (0.9-1.8 GHz) EMF are of special interest. Numerous studies have found that EMF can cause various effects. A positive effect includes healing of bone fractures and treatment of osteoarthritis (Hulme et al., 2002; Kovacic and Somanathan, 2010). A negative effect includes DNA damage, reproductive disorders, immunosuppression, and neurologic disorders (Habash et al., 2009). But so far, there has been a controversy whether EMF has a negative effect on human health or not.
Protein kinase C (PKC) has been implicated in slowing ciliary beating (Kobayashi et al., 1989; Kobayashi et al., 1992; Wyatt et al., 2000; Wong et al., 1998). This role of decreasing CBF upon PKC activation has been consistent in all mammalian cilia observed. But, the mechanisms by which PKC inhibits CBF are not fully understood. Based on homologies and profiles of activation by specific co-factors, PKC have been classified into three major groups. There are the classical or conventional PKC (cPKC), the calcium-insensitive novel PKC (nPKC), and the atypical PKC (aPKC). The physiological
- 2 -
functions of cPKC, nPKC, and aPKC isoforms are well known. Thus we want to know whether any of these PKC isoforms were responsible for the EMF-induced decrease in CBF.
Particularly, the widespread and rapidly growing use of mobile phones has led to a wide-ranging discussion about possible interactions of EMF with the human body. However, there are few studies examining the effects of mobile phone frequency range EMF on the human respiratory system. In this study, we focus on the effects of EMF on human respiratory CBF and we aimed to investigate the effects of a 1.8 GHz EMF on human mucociliary transport and also to investigate the underlying regulatory mechanisms.
- 3 -
II. Materials and methods
A. Normal human nasal mucosa culture
Nasal mucosa was obtained from 18 patients (10 males, 8 females) who underwent nasal surgery for nasal obstruction, nasal biopsy, trans-sphenoidal hypophysectomy, and rhinoplasty. The patients were all non-smokers, non-asthmatic, on no medication and had no respiratory illness, including recent colds. Mucosa samples were washed with normal saline and cut into small pieces (0.5x0.5 cm). The samples were cultured in 6-well plates containing Dulbecco’s Modified Eagle Medium-Ham’s nutrient F12 (DMEM/F12; Invitrogen, USA) with 10 IU/mL penicillin and 10 μg/mL streptomycin. The samples were stabilized for 24 hours in an incubating chamber (CU-109, Live Cell Instrument Inc., Korea) at 37 ℃, 5% CO2, and 100% humidity. The culture medium was exchanged 24 hours later to remove mucus and
cellular debris. The cultured nasal mucosa was prepared for CBF measurement under different conditions. This culture was used for CBF response after a 1.8 GHz EMF exposure and normal human nasal epithelial (NHNE) cell culture was used for the other experiments. The experimental protocol used in this study was approved by Institutional Review Board of Ajou University Hospital and informed consent was obtained from parents or guardians of all patients. (AJIRB-05-184)
B. NHNE cell culture
Human nasal inferior turbinate specimens were collected in the same manner as described above. The passage-2 NHNE cells were prepared as previously described (Yoon et al., 2001). Briefly, the epithelial cells from the turbinates were treated 1% Pronase (Type XIV pronase, Sigma Aldrich Chemical Co., St. Louis MO) for 18 to 24 hours at 4 ℃. To remove fibroblasts, endothelial cells, and myoepithelial cells, isolated cells were placed in a plastic dish and cultured for 1 hour at 37 ℃. Isolated epithelial clusters were divided into single cells by incubating them with 0.25% trypin/EDTA. Passage-2, NHNE cells (6x105 cells/well) were seeded in 0.5ml of culture medium into 24.5 mm, 0.45 μm pore size,
Transwell-Clear (Costar Co., Cambridge, MA, USA) culture inserts. Cells were cultured using a 1:1 mixture of bronchial epithelial cell growth medium (BEGM) to DMEM containing all supplements. Cultures were grown submerged for the first 7 days, during which time the culture media were changed on every other day. The air-liquid interface (ALI) was created on day 8 by removing the surface of the
- 4 -
monolayer and the cells fed via the basal chamber. The culture medium was changed daily after the ALI was initiated, and experiments used NHNE cells on day 14 after the creation of the ALI. Cultures were stabilized for 24 hours in incubating chamber and maintained at 37℃ in a humidified atmosphere of 5% CO2 in air.
C. Exposure to EMF
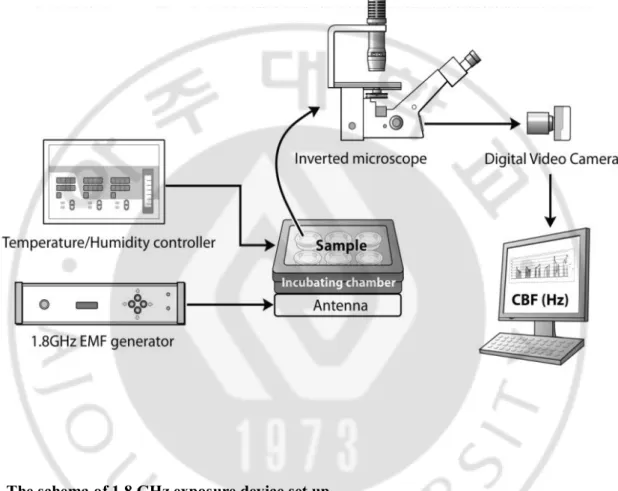
Samples were exposed to a 1.8 GHz continuous wave driven by a dipole antenna for three days using a signal source (SS-01, EMF Safety Inc., Seoul, Korea). EMF generator provided 1 W (27 dBm) during the exposure period (Fig. 1). Mucosa samples were positioned in close contact above the dipole antenna. Six well plates were exposed simultaneously. These plates were placed in the H-field maxima. Each of the three plates (exposed, not-exposed) was placed inside an incubator to ensure constant environmental conditions. In addition, the temperature and the specific absorption rate (SAR) were monitored at the location of the plates during exposure and strictly controlled during the experiments. SAR for mucosa samples at the central point of the media, whose measured relative dielectric constant was 76.07, and conductivity was 2.45 S/m at 1.8 GHz, was set for 1 W/kg. The SAR value was measured by an E-field probe (ET25, EMF Safety Inc., Seoul, Korea). Each exposure time was 0 h, 0.1 h, 0.3 h, 0.5 h, 1 h, 3 h, 6 h, 9 h, 24 h, 48 h, and 72 h after initial exposure. The control group was not exposed to an EMF and protected from electromagnetic wave with aluminum foil, although all the other conditions were identical to the experimental group.
- 5 -
Fig. 1. The schema of 1.8 GHz exposure device set up.
Samples were exposed to a 1.8 GHz continuous wave and Ciliary motility is observed and recorded using a microscope and workstation. A analysis software in workstation measured CBF.
- 6 -
D. MTT assay
The MTT (3-(4,5-dimethylthiazol-2yl)-2,5-diphenyl-2H-tetrazolium bromide) assay was used to examine the cytotoxicity of EMF-exposed cells. Passage-2, NHNE cells were seeded into 96-well plate at evenly density of 1 x 105 cells/ ml. These cells were incubated with 0.5 mg/ml MTT (Sigma, St.Louis,
USA) for 4 hour in humidified CO2 incubator at 37 ℃, 5% CO2, and exposed to a 1.8 GHz EMF. After
incubation, the supernatant was removed and 100 μl DMSO was added to each well. Then the plate was strongly shaked and measured by spectrophotometer using a reference wave length of 540nm. Data were calibrated by measuring the absorbance of the control wells, which represents the maximum growth (100%) in complete growth medium.
E. Measurement of CBF
Ciliary movement was analyzed using a program with an optical flow technique with a peak detection method for CBF measurement (Kim et al., 2011). Ciliated cells on the mucosal epithelium, at 37 ℃ under 5% CO2, were magnified using 400x objective under an inverted microscope (Axiovert 40 CFL, Carl
ZEISS, Göttingen, Germany) that was equipped with a charged coupled device camera (Moticam 2000, Motic Inc., Hong Kong, China). This digital camera projected an image of the ciliary beat onto a monitor. Actively moving cilia were recorded 50-100 frames per second using digital camera and stored in a workstation. Application was developed to use Microsoft Visual C++ 2005 (Microsoft, Redmond, WA, USA). The developed application automatically measured CBF from a stored image. This study applied optical flow for image processing. Optical flow is one of the most general studies of motion recovery approaches. Signal processing was done by peak detection algorithm. The peak detection method finds a peak of signals over average values with weight calculated by the percentage of the average.
We analyzed CBF data between program and manual count group during total experiment period. The measured CBF was compared with manually measured CBF using a digital high-speed video method and there was no difference between program and manual count group. The gold-standard used in this paper was the digital-speed video method, for which CBF is measured manually by an expert observing the video in slow play. Videos were presented to the observer in a random order with no information given regarding the CBF. Ciliary beating was observed frame by frame. The observer pressed the button of the
- 7 -
application, which moves one frame, and measured ciliary beating from the obtained image. The observer measured until 10 cycles, and noted the starting time and ending time of the image sequence. CBF was calculated using following equation (Kim et al., 2011).
CBF =
x 10
Before EMF exposure, samples were stabilized for 12 h, and basal CBF was measured. Samples were exposed to a 1.8 GHz EMF using a signal source, and CBF was measured at 0 h, 0.1 h, 0.3 h, 0.5 h, 1 h, 3 h, 6 h, 9 h, 24 h, 48 h, and 72 h thereafter.
F. The PKC kinase activity assay
Protein Kinase C (PKC) is a large superfamily of serine/threonine kinases that mediate essential cellular signals required for activation, proliferation, differentiation and survival. There are at least ten PKC isotypes that are closely related in structure but that have distinct patterns of tissue distribution and function. The PKC isotypes can be subdivided into three classes based on primary structure and biochemical properties. All PKC isotypes share a characteristic sequence motif C1 in addition to a serine/threonine-protein kinase domain. The PKC Kinase Activity Assay was obtained from Enzo Life Sciences (Plymouth Meeting, PA). This assay kit is based on a solid phase enzyme-linked immune-absorbent assay (ELISA) that utilizes a specific synthetic peptide as a substrate for PKC and a polyclonal antibody that recognizes the phosphorylated form of the substrate phosphorylated by PKC. PKC activity was performed according to instructions of manufacturer. In brief, the substrate, which is readily phosphorylated by PKC, is pre-coated on the wells of the provided PKC Substrate Microtiter Plate. The samples to be assed are added to the appropriate wells, followed by the addition of ATP to initiate the reaction, cells were incubated at 30℃ for 90 mins, and kinase reaction is terminated by Phosphospecific Substrate Antibody to bind specifically to the phosphorylated peptide substrate. The phosphospecific antibody is subsequently bounded by a peroxidase conjugated secondary antibody. The assay is developed with tetramethylbenzidine (TMB) substrate and a color develops in proportion to PKC phosphotransferase activity. The intensity of the color is measured in microplate reader at 450nm. The PKC activity was expressed as a relative activity.
- 8 -
G. Elucidation of the role of PKC in EMF-induced decrease of CBF
The role of PKC in the EMF-induced responses of CBF was studied using the non- specific PKC inhibitor, Calphostin C (Calbiochem, Japan). Calphostin C (C44H38O14) is a naturally occurring
perylenequinone antibiotic from the fungus Cladosporium cladosporoides FERM BP-1285, which was originally isolated from a block fence in Osaka, Japan. It has been shown to have a pleiotropic biological activity profile. a number of studies have demonstrated that this natural product can inhibit PKC, and cause apoptotic cell death by an as yet undefined mechanism.
After the measurement of baseline CBF for a minimum of five minutes, the Calphostin C solvent, dimethylsulfoxide (DMSO) group (i.e., negative control) was used to rule out the influence on CBF. Following this, 10 μmole Calphostin C was added to the ciliated cells with DMSO and exposed to the 1.8 GHz continuous wave. The control group was not exposed to an EMF. However, all other conditions were identical between the control and experimental groups.
H. Determination of PKC isoforms involved in the EMF-induced decrease of CBF
PKC is a serine/threonine protein kinase that consists of three superfamilies. The conventional PKC (cPKC) isoforms are Ca2+ and diacylglycerol (DAG) dependent, whereas novel PKC (nPKC) isoforms are
Ca2+ independent and DAG dependent. The atypical PKC (aPKC) isoforms are Ca2+ and DAG
independent. To determine if any of these isoforms are responsible for the EMF-induced decrease of CBF, two types of PKC inhibitors were used, Gö-6976 (cPKC inhibitor) and GF 109203X (nPKC inhibitor). After the measurement of baseline CBF for a minimum of five minutes, each group of the individual PKC inhibitor alone (i.e., negative controls) were used to rule out the influence on CBF when compared to the control group. The ciliated cells with these PKC inhibitors (1 μM Gö-6976 and 10 μΜ GF 109203X) were exposed to a 1.8 GHz EMF (SAR= 1.0 W/kg). The control group was not exposed to an EMF. However all other conditions between the experimental and control groups were identical.
- 9 -
I. The experiments of external environmental factors
External environmental factors that affect CBF are temperature, pH, and humidity. Most the effects of these environmental factors were excluded using a controlled incubator. We have measured temperature and pH of incubator using microsensor.
J. The measurement of real time temperature
The real time temperature measurement was done to exclude temperature effect experimentally between two groups (control vs EMF exposed group).
K. The effect of 0.2 ℃ temperature difference
The maximal temperature difference was 0.13 ℃ in prior experiment. Additional experiment was performed again to identify the effect of 0.2 ℃ on CBF without EMF exposure. CBF was measured under 0.2 ℃ temperature difference setting, although all the other conditions were identical.
L. The pH measurement for 24 hours
The pH measurement was done to exclude pH effect experimentally between two groups (control vs EMF exposed group). Briefly, cells grown on coverslips were washed with standard N-2-hydroxyethylpiperazine-N-2-ethane sulphonic acid (HEPES) solution and loaded with 2.5 μM BCECF-AM at 37 ℃ for 15-30 min and again rinsed 3 times. For fluorescent measurements, a Lambda DG4 excitation system (Sutter, Novato, CA, USA) was used with 10 nm wide excitation filters centered on 480 and 405 nm (Chroma Technology Corp., Brattleboro, VT, USA). Nigericin was used to calibrate pHi measurement. BCECR-loaded cells were perfused with calibration solutions containing 15μM
nigericin and 130 mM KCl at pH 6.8, 7.2, 7.5 and 7.8, respectively while the fluorescence ration was measured. The ratio data were normalized to the ratio at pH 7.2 (normalized fluorescence ratio: NFR). each ratio value was accepted when the ratio reached a steady state level at a given calibration pH. Then,
- 10 -
the calibration curve was constructed by plotting the average NFR values against the corresponding pH values. After calibration, cells was exposed to EMF and the each fluorescence ratio value (480/405 nM) was measured.
M. The measurement of intracellular calcium concentration ([Ca2+]i)
Cells were loaded with Fura-2 by incubation at 37 ℃ for 30 minutes in 10 µM Fura-2 AM (Calbiochem, USA) in Ca2+-containing (1.3 mM) Hanks’ buffered salt solution without phenol red (HBSS, Bibco-BRL),
but additionally buffered with 25 mM HEPES (HEPES buffered-HHBSS, pH 7.4). Cells were subsequently centrifuged, resuspended, washed twice and then plated onto poly-l-lysine coated 32mm square glass cover-slips and incubated at room temperature for 30 min to allow for the de-esterification of the Fura-2 AM. The cover-slips were mounted on the stage of an inverted microscope and placed in small using holding chamber (AKI Institute, U of Copenhagen, Denmark) and exposed to EMF. Cells were visualized on the same inverted microscope. Fluorescence was detected with a camera, recorded by an optical memory disc recorder at 30 fps, and digitized by computer. Images of [Ca2+]
i were calculated from
single wave length recordings at 380 nm referenced to ratiometric measurement. Initial [Ca2+]
i reference
images were based on 10 frames recorded at 350 and 380 nm. Changes in [Ca2+]
i were recorded by
monitoring changes in fluorescence with an excitation wavelength of 380nm. Additional reference images were taken at 350nm every 30 seconds. All images were subjected to background subtraction and correction for shading and bleaching. The measurement of intracellular calcium concentration was done to confirm EMFinduced change of [Ca2+]i experimentally between two groups (control vs EMF exposed
group).
N. Transmission electron microscope (TEM)
For transmission electron microscope(TEM), the cultured cells on the porous membrane were fixed with chilled 2.5% glutaraldehyde for 4-6 hour and were then washed with 0.1 M PBS. The cells were post-fixed with 1% osmium tetroxide for 2 hours. The specimens of control and EMF-exposed group were observed with transmission electron microscope (902A TEM, Zeiss, Jena, Germany) to identify the structural changes by EMF.
- 11 -
O. Statistical analysis
All experimental data are expressed as the mean ± S.E.M. MTT assay and PKC activity data were examined using an unpaired t test and the CBF response after a 1.8 GHz EMF exposure data was examined using Mann-Whitney test. Values of p less than 0.05 were considered to be statistically significant.
- 12 -
III. RESULTS
A. The CBF response after a 1.8 GHz EMF exposure
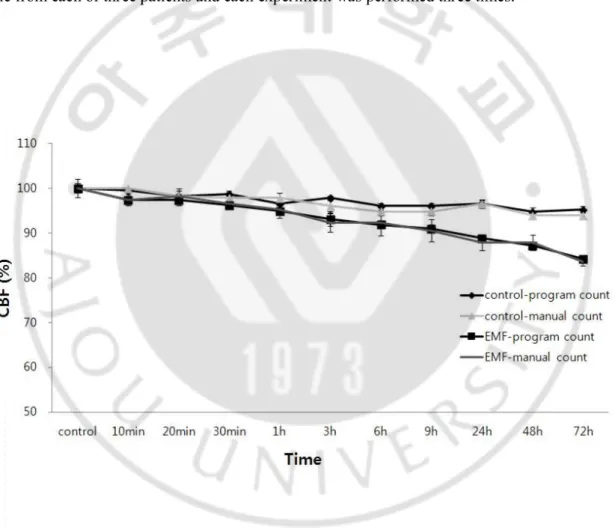
Baseline CBF of control group was 14.43 Hz and baseline CBF of EMF-exposed group was 14.50 Hz. CBF in the 1.8 GHz EMF-exposed group was decreased in comparison to the control group. In the EMF-exposed group, CBF began to decrease from the initiation of EMF exposure in comparison to the control group, with a maximum difference of 11% at 3 days (Fig. 2). In each experiment, we obtained one sample from each of three patients and each experiment was performed three times.
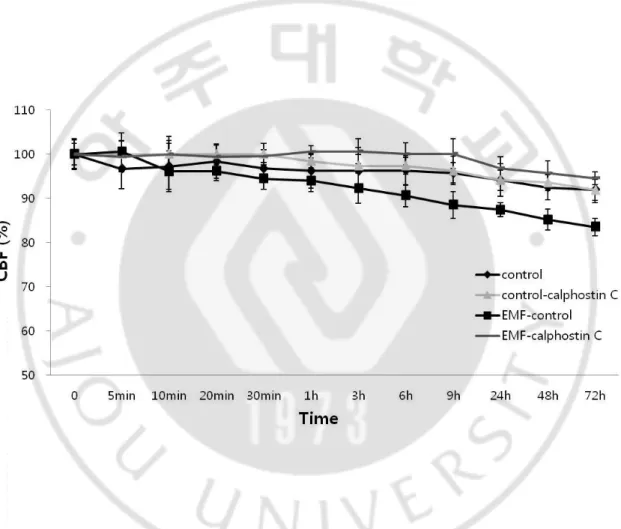
Fig. 2. The 1.8 GHz EMF-induced responses on CBF.
Human nasal mucosal cells were exposed to a 1.8 GHz EMF for three days. The control group was not exposed to EMF, although all other conditions were identical. CBF was calculated by using the program and manual count.
- 13 -
B. MTT assay
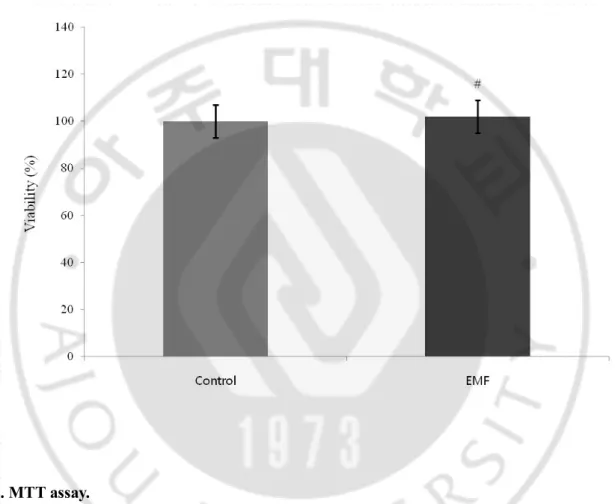
The MTT assay showed no significant difference (p>0.05) in the survival rates between the 1.8 GHz EMF-exposed and control groups (Fig. 3).
Fig. 3. MTT assay.
NHNE cells were exposed to a 1.8 GHz EMF and absorbance was measured at 540 nm. The control group was not exposed to EMF, although all other conditions were identical. Results are reported as mean ± S.E.M. # p>0.05, compared with control group.
- 14 -
C. PKC activity assay
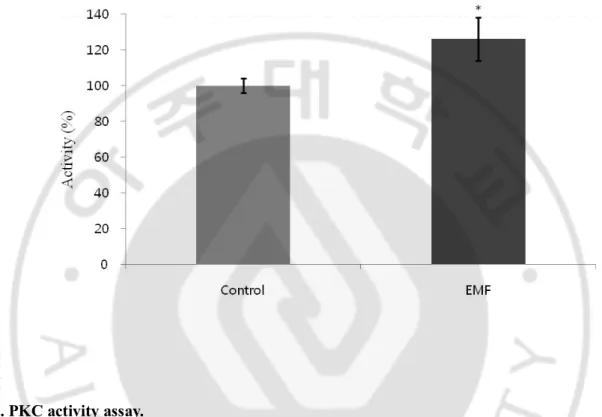
A significant increase (p<0.05) in PKC activity was observed in the 1.8 GHz EMF-exposed group (Fig. 4).
Fig. 4. PKC activity assay.
A 1.8 GHz EMF-exposed sample protein was assayed using the manufacturer’s instructions with absorbance measured at 450 nm, and kinase activity (O.D./μg protein) was quantified by the change in absorbance over background levels. Results are reported as mean ± S.E.M. * p<0.05, compared with control group.
- 15 -
D. The role of PKC in the EMF-induced decrease of CBF
To explore if the EMF-induced inhibition of CBF was due to the activation of PKC, broad PKC inhibitor, Calphostin C, was used. Calphostinc C competes at the diacylglycerol (DAG) and phorbol 12-myristate 13-acetate (PMA) binding site of the cysteine rich regulatory domain of the PKC. DMSO alone did not affect CBF, and addition of 10 mM Calphostin C did not change CBF. As expected, Calphostin C abolished the EMF-induced decrease in CBF, thus providing additional evidence that the activation of PKC was involved in the EMF-induced suppression of CBF (Fig. 5).
Fig. 5. The effect of Calphostin C on a 1.8 GHz EMF-induced response on CBF.
The role of PKC in the EMF-induced responses of CBF was studied using the non- specific PKC inhibitor, Calphostin C. Calphostin C (10 μΜ) was added to the ciliated cells with dimethylsulfoxide (DMSO), which were then exposed to the EMF. The Calphostin C solvent, DMSO alone group was used as a negative control. The control group was not exposed to the EMF, although all other conditions were identical.
- 16 -
E. Determination of PKC subtypes
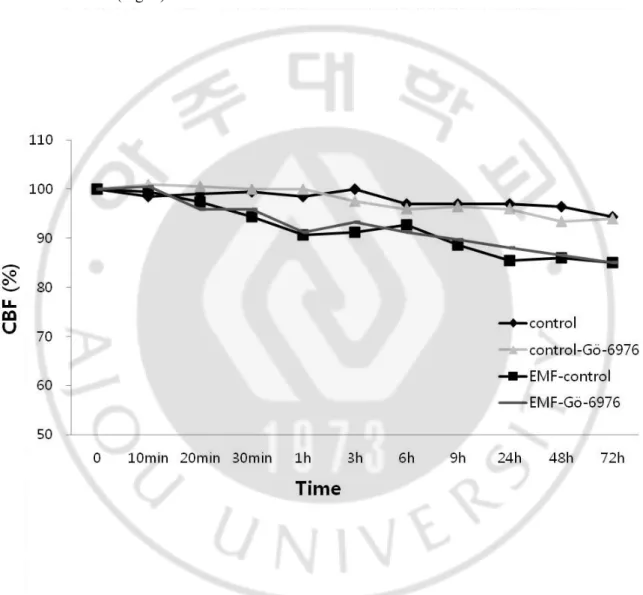
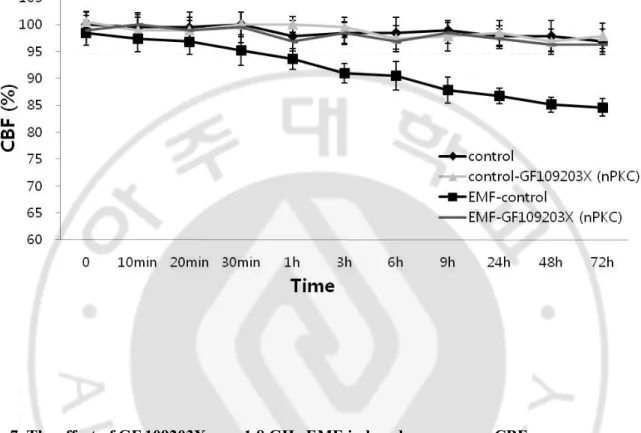
Gö-6976(cPKC inhibitor) alone did not affect CBF in comparison to the control group, and the EMF-induced CBF decrease was not affected in the presence of Gö-6976 (Fig. 6). A structural analog of staurosporine, GF 109203X (nPKC inhibitor), was then used to determine if the EMF-induced inhibition of CBF was mediated through nPKC isoforms. GF 109203X alone did not affect CBF in comparison to the control group, and pretreatment of the cells with GF 109203X prevented any decrease of an EMF-induced CBF (Fig. 7).
Fig. 6. The effect of Gö-6976 on a 1.8 GHz EMF-induced response on CBF.
Gö-6976 (cPKC inhibitor) was used to determine if any of PKC isoforms are responsible for the EMF-induced response of CBF. 1 μΜ Gö-6976 alone was used as a negative control to rule out the influence on CBF in the comparisons with the control group. The ciliated cells with 1 μM Gö-6976 added were exposed to a 1.8 GHz EMF. The control group was not exposed to EMF, although all other conditions were identical.
- 17 -
Fig. 7. The effect of GF 109203X on a 1.8 GHz EMF-induced response on CBF.
GF 109203X (nPKC inhibitor) was used to determine if any of PKC isoforms are responsible for the EMF-induced response of CBF. 10 μM GF 109203X alone was used as a negative control to rule out the influence on CBF in the comparisons with the control group. The ciliated cells with 10 μM GF 109203X added were exposed to a 1.8 GHz EMF. The control group was not exposed to EMF, although all other conditions were identical.
- 18 -
F. The measurement of real time temperature
Between two groups (control vs EMF-exposed group), the average temperature difference was 0.08 ℃ and the maximal temperature difference is 0.13 ℃ (Fig. 8).
Fig. 8. The real time temperature measurement of control and EMF-exposed group.
The real time temperature measurement was done to exclude temperature effect experimentally. Between two groups (control vs EMF exposed group), the average difference was 0.08 ℃ and the maximal temperature difference is 0.13 ℃.
- 19 -
G. The effect of 0.2 (℃) temperature difference
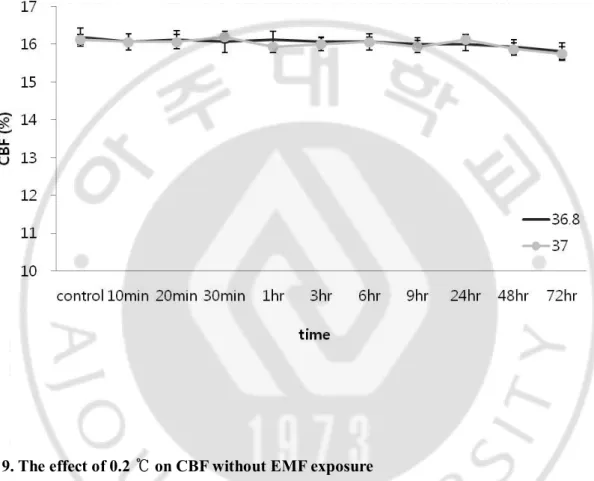
There was no difference in CBF between two groups (Fig. 9).
Fig. 9. The effect of 0.2 ℃ on CBF without EMF exposure
CBF was measured under 0.2 ℃ temperature difference setting to exclude temperature effect
experimentally. Results showed no significant difference (p>0.05) between 36.8 ℃ and 37.0 ℃ group. Results are reported as mean ± S.E.M. p>0.05, compared with control group.
- 20 -
H. The measurement of pH for 24 hours
There was no difference in CBF between two groups (Fig. 10).
Fig. 10. The pH measurement of control and EMF-exposed group for 24 hours.
The real time pH measurement showed no significant difference (p>0.05) in pH between the control and 1.8 GHz EMF-exposed groups. Results are reported as mean ± S.E.M. ** p>0.05, compared with control group.
- 21 -
I. The measurement of intracellular calcium concentration ([Ca2+]i)
The measurement of intracellular calcium concentration was done to confirm EMFinduced change of
[Ca2+]
i experimentally between two groups (control vs EMF exposed group). The average [Ca2+]i of
control group was 270 nM. The average [Ca2+]
i of EMF-exposed group was 229 nM. Exposure to EMF
caused a statistically significant decrease of intracellular calcium concentration ([Ca2+] i).
Fig. 11. The measurement of intracellular calcium concentration ([Ca2+] i)
Intracellular calcium concentration was measured using Fura-2/AM. Exposure to EMF caused a statistically significant decrease of intracellular calcium concenetration ([Ca2+]
i). [Ca2+]i was quantified by
the fluorescence ratio of 350/380 nM. Results are reported as mean ± S.E.M. ## p<0.05, compared with
- 22 -
J. Transmission electron microscope (TEM)
There was no difference in the ciliary ultrastructure between control and EMF-exposed group. As seen in the cross section (Fig. 11), EMF-exposed group has a normal ring of nine microtubular doublets, each with a pair of dynein arm, connected by radial spoke to a central pair of microtubules (9+2 microtubulets).
Fig. 12. The cross section of ciliary ultrastructure by TEM.
EMF-exposed group was examined with transmission electron microscope (TEM) to identify the structural changes by EMF. As seen in the cross section, EMF-exposed group has a normal ring of mine microtubular doublets, each with a pair of dynein arm, connected by radial spoke to a central pair of microbutules (9+2 microtubules).
- 23 -
IV. Discussion
The primary function of MCT system in the respiratory tract is to eliminate inhaled particles, which is facilitated by the propulsion of mucus. The overall propulsive effect depends on the arrangement of the cilia, their metachronal relationships, and the CBF (Satir and Sleigh, 1990). Functioning as a key factor for MCT regulation, CBF controls MCT efficiency as substantial increase in MCT efficiency requires an equally substantial rise in CBF. When measured in vitro, human CBF is approximately 12-15 Hz at normal body temperature (Salathe, 2007). Alteration of CBF may occur by various harmful materials, and lead to an impairment of MCT.
The EMF is one of the four fundamental forces in the universe. Modern technical progress is associated with a growing number of artificial EMFs in the environment. These artificial EMFs can interfere with basic biological processes, thus potentially affecting the health of the population. EMF is generated by the interaction between an electrical currents and a magnetic field, resulting from the acceleration of an electric charge, and then propagates at the speed of light in waves of a specific wave length that oscillate at a defined frequency. So EMF is often described by its frequency, which is the number of oscillations of the perpendicular electric and magnetic fields per second and is expressed in hertz (Hz). Depending on the frequency, an EMF can be classified by the human effect consideration by ICNIRP guidelines as follows: extremely-low frequency (ELF, 0-3 kHz) and radiofrequency radiation (RFR, 3 kHz - 300 GHz). Electromagnetic waves with wavelengths shorter than RFR include infrared, visible light, ultraviolet, x-ray, and gamma-ray.
The global system for mobile phones operates in the frequency band between 0.8 and 3.5 GHz. Frequencies ranging from 0.9 to 1.8 GHz are of special interest because this frequency band is occupied by the most of the mobile phones. The biological effects of this mobile phone frequency range have been a worldwide focus of study for many decades due to the rapid growing use of this technology. The effects of EMF on human body are largely thermal and non-thermal effects (Braveman et al., 1998), and exposure of human to radiofrequency radiation (RFR) is known to induce internal body currents and energy absorption in tissue, depending on frequencies and coupling mechanisms. Thermal effects produced by temperature rise are well understood. However, the focus of the present study with respect to the health of human exposure to RFR is to evaluate the possibility of non-thermal effects and what extent non-thermal RFR field exposure may exert on human health. RFR at frequencies from 900 MHz to 1.8 GHz are special interest because of rapid expansion of wireless technology using this frequency range.
- 24 -
The field strengths used by most wireless devices are so low that thermal effects are likely to explain any possible health effects (Hietanen, 2006). Foster et al. showed that RFR emitted by cellular phones does not cause damage to tissue via thermal means of radiation damage (Foster et al., 2000). In our study, we controlled temperature and humidity by an incubator, and there was no difference in temperature between EMF-exposed and control group. Thus it is assumed that the effect of EMF on nasal mucosa is non-thermal.
Biological effects of EMF have been the subject of the study for many decades worldwide, however EMF of cellular phone has especially been controversial with regard to health, because of rapidly growing use and concern that the EMF may exert negative effects on health. Potential biological effects of mobile phone frequency range on the human body include enhanced healing of bone fractures, cellular growth, phagocytic activity, permeability of the blood-brain barrier, vasodilatation, and the progression of cataracts (Fritze et al., 1997; Kolomyceva et al., 2001; Kwee and Raskmark, 1998; Paredi et al., 2001; Satter et al., 1999; Ye et al., 2001). Especially, the adverse effects of mobile phone EMF (800- 2,500 MHz) on human body include largely genotoxicity, cytotoxicity, teratogenicity, carcinogenicity, immunodeficiency, neurological, and behavioral effects (Behari J, 2010). Genotoxicity includes DNA damage and chromosomal abnormalities (Maes et al., 1993; Lai and Singh, 1996). Cytotoxicity includes reduced cellular growth of skin fibroblast after exposure 960 MHz EMF (Velizarov et al., 1999). Immunologically, phagocytic activity of human neurotrophil was reduced by 50% after electromagnetic wave exposure (Kolomyceva et al., 2001). With regard to central nerve system disorders, visual reaction time delay and decreased short-term memory loss have been observed after AM radio band exposure. In addition to, it has been reported that mobile phone use has been associated with temporal lobe and occipital lobe brain tumors (Hardell et al., 2000). Therefore, it appears highly likely that the emission of these RFRs may effect on human nasal mucosa too. However, until now there has been no study to determine the effects of the EMF on MCT of human nasal mucosa.
To determine if cell death was caused by the EMF, an MTT assay was performed. The results indicated no significant difference between the EMF-exposed group and control group. This suggests that the reduction of CBF was associated with decreased ciliated cell function, and that the EMF had no cytotoxic effect on NHNE cells. EMF-exposed group was examined with transmission electron microscope (TEM) to identify the structural changes by EMF, but there was no difference in the ciliary ultrastructure between control and EMF-exposed group. EMF-exposed group has a normal ring of nine microtubular doublets, each with a pair of dynein arm, connected by radial spoke to a central pair of microtubules (9+2 microtubules). The inner pair of microtubules in cilia with the 9+2 configuration is critical for the back and forth beating pattern.
- 25 -
External environmental factors that affect CBF are temperature, pH, and humidity. CBF is not critically dependent upon temperatures in the range of 32℃ to 40℃ and pH levels in the range of 3.5-10.5 (Clary-Meinesz et al., 1998; Green et al., 1995). In the present study, the effects of these external environmental factors were excluded using a controlled incubator. Also, we have measured temperature and pH of incubator in real time using microsensor. Between two groups (control vs EMF exposed group), the average temperature difference was 0.08 ℃ and the maximal temperature difference is 0.13 ℃. Additional experiment was performed again to identify the effect of 0.2 ℃ on CBF without EMF exposure. CBF was measured under 0.2 ℃ temperature difference setting, although all other conditions were identical. As a result, there was no difference in CBF between two groups. We also have estimated pH between two groups, there was no difference. Therefore, we confirmed that there were no differences in the temperature & pH effect on CBF between the EMF-exposed and control groups experimentally.
The most commonly used method for CBF measurement is the combination of image processing based on difference mage, and signal processing based on fast Fourier transform (FFT). In our study, we measured CBF using the newly developed image processing system. This system is based on optical flow for image processing and peak detection for signal processing. Optical flow has the advantage of tracking of overlapping objects. For this reason, we used optical flow for measurement of CBF. In our previous paper, we found that one peak height is one ciliary beating cycle (Kim et al., 2011). Therefore, the peak detection method is simpler than the FFT and the peak detection method finds a peak of signals over average values with weight calculated by the percentage of the average. The gold-standard of CBF measurement is the digital high-speed video method with a manual count of CBF in slow motion video play. There are many of methods have been developed for CBF measurement. For example, there are synchronization of a stroboscope, high-speed cinematography, photomultiplier and photodiode technique using a photo-electric signal (Yager J et al., 1978; Teichtachl et al., 1986). The limitation of these methods is time-consuming, high price, indirect estimation and microscopic image, and observer fatigue. For this reason, we analyzed ciliary movement using an optical flow technique with a peak detection method for CBF. This technique offers several advantages, including relatively low cost of equipment, ease of use, and highly compatible digital recording media.
- 26 -
For the minimum sampling rate for CBF measurement, we abide by the Nyquist criterion, which postulates that the sampling interval has to be equal to or less than one-half the period of the frame rate being recorded. The limitation of previous video image capture methods is slow sampling rate at 30 fps, thus decreased accurate CBF measurements to frequencies of < 15 Hz. In our study, baseline CBF was 14.43 Hz. We used a > 50 fps recorded image to solve these limitations. This frequency rate expands most CBFs measured in higher reference animals.
A quantity frequently used to measure the dosimetry of electromagnetic radiation absorbed by a tissue is the specific absorption rate (SAR), which is measured in W/kg. The regulated exposure limit of mobile phones by American Federal Communication Commission (AFCC) is a SAR of 1.6 W/kg for 1 gram sample. In our study, an EMF within the limit of human protection guideline 1 W/kg SAR was used, and CBF was decreased in the EMF exposed group. These results imply that EMF-induced homeostasis mechanisms for EMF exposure did not properly compensate. Previous studies showed that CBF in the mucosal explants from the majority of patients with chronic sinusitis was not different from that measured in the explants of healthy tissue (Kim et al., 2007).However, the increase of CBF by ATP showed differences between the two groups. In the normal sinus mucosa group, 100 μM of ATP strongly stimulated CBF with an average increase of 30.1%, whereas the sinusitis group showed only a slight increase of CBF by ATP with a range of 3.2~12.2%. These findings indicate that sinusitis mucosa initially compensate for mucosal damage or inflammation, but ultimately can not afford to compensate for abnormal conditions, suggest that repeated exposures to EMFs over a long period of time could be influential to cilia function and possibly lead to a pathological state.
MCT is influenced by ciliary activity and properties of the mucus layer. Determinants of MCT include swallowing, sniffing, gravity, nasal anatomy, and properties of respiratory gas [Warwick, 1983]. Duchateau et al. found a linear correlation between CBF and MCT when measured with the saccharine dye test (Duchateau et al., 1986), and Seybold et al. claimed that changes in ciliary beat cause a fourfold change in mucociliary transport (Seybold et al., 1992). In another study, a relatively small increase in CBF (16%) was shown to result in a large increase (56%) in surface liquid velocity (SLV) (Seybold et al., 1990). Although CBF and MCT may not directly be proportional for all kinds of stimuli, the fact that the change of MCT is larger than that of CBF is common. In our study, the EMF-exposed group showed an approximate 11% CBF reduction. Therefore, MCT might have been inhibited more than 11%. Thus, the novel finding that the EMF acts directly on ciliated cells to decrease CBF may suggest its potential to produce impaired mucociliary function in persons with diseases of the airways. As a result, the impairment of MCT by the EMF leads to increased susceptibility to respiratory infections such as rhinosinusitis.
- 27 -
Ciliary beating can be stimulated by a variety of mechanisms, one of which is cyclic nucleotides. Several studies have shown that intracellular cyclic nucleotides act as important regulators of ciliary motility (Lansley et al., 1992; Wyatt et al., 2005). By increasing the levels of cAMP, ciliary motility is stimulated through the activation of the cAMP-dependent protein kinase A (Wyatt et al., 1998). Conversely, agents that activate protein kinase C (PKC) have been associated with the lowering of CBF (Salathe et al., 1993; Wong et al., 1998, Wyatt et al., 2000). PKC, a ubiquitous enzyme that mediates the transmembrane signal transduction process has been recognized as a second messenger (Nishizuka, 1988). Until now, PKC is the only known signal transduction pathway to decrease CBF (Salathe, 2007). The finding of decreasing CBF upon PKC activation has been consistent in all mammalian cilia examined. For example, cigarette smoke, H202, and neuropeptide Y (NPY) have been shown to induce PKC activity in
airway epithelial cells and decrease CBF through a PKC-dependent pathway (Kobayashi et al., 1992; Simet et al., 2009; Wong et al., 1998).Particularly, NPY was found to inhibit CBF in cultured human tracheal and bronchial ciliated cells, presumably mediated through a novel PKC isoform-mediated pathway (Wong et al., 1998). Therefore we proposed that the intracellular mechanisms whereby EMF suppressed CBF in human nasal mucosa can be explained by following hypothesis. EMF can activates nPKC, which increases the activaties of the Ca2+-ATPase pumps. This in turn causes a decrease in
intracellular Ca2+ concentration [Ca2+]
i to a very low level. The decreased CBF have been associated with
a substantial decrease in the basal intracellular Ca2+ concentration. We also identified decreased
intracellular calcium level experimentally.
These studies show that the decrease of CBF is mediated through a PKC-dependent pathway, and that the rate of decreased CBF by PKC is approximately 11~23%. In our present study, we identified increased PKC activity in the EMF-exposed group and nPKC was involved during the inhibitor studies. Overall the decreased CBF rate (11%) by EMF exposure was not significantly different from these studies. The earlier studies together with our present results indicate that nPKC can modulate ciliated cells, therefore slowing the response to various stimuli.
Our study is solely in vitro experiment. Therefore, it may be different from actual mobile phone usage due to several factors. First, when using a mobile phone, the EMF can reach nasal mucosa through skin, soft tissue, and bone. Thus, the net EMF effect would be smaller than expected because of the small amount of energy that actually reaches the nasal mucosa. Second, the hours of mobile phone use must be considered. Although continuous exposure for several days, as occurred during this experiment, does not occur in everyday life, the use of wireless communications has exponentially increased over the past several decades. In addition, there has been expansive growth in wireless communications infrastructure and services. This includes mobile phone handsets, digital data communication devices, bluetooth
- 28 -
interfaces, wireless local area networks, including wireless fidelity (Wi-Fi), 3-G networks, radar stations, microwave ovens, and wireless keyboards. The amount of exposure accumulated in the body from these sources should not be ignored. The majority of studies on RFR have been conducted with short-term exposures, (i.e., a few minutes to several hours), and therefore, little is known about the effects of long-term exposure which would be experienced by people living near telecommunications installations, who have been exposed for months or years. Persson et al.reported an increase in permeability of the blood-brain barrier in mice, suggesting that a short-term/high intensity exposure can produce the same effect as a long-term/low intensity exposure (Persson et al., 2001). Indeed, there is evidence that effects of RFR accumulate over time (Heikkinen et al, 2003; Repacholi, 2001; Wyatt et al., 1998). Philips et al. reported DNA damage in cells after 24 hours of exposure to low intensity RFR, leading to gene mutations that accumulate over time (Philips et al., 1998). This could be another indication that RFR effects can accumulate over time. Thus, these cumulative exposures might interfere MCT of the human nasal mucosa. Third, although the effect of EMFs may be minimal in healthy people, children and the elderly with pathologic conditions can be more severely affected, even with small quantities of EMF. A particular consideration is mobile phone use by children. Although no specific evidence exists, there is a general concern that children or adolescents may be more sensitive to RFR exposure than adults. Children are more vulnerable, because their bodies are developing and their respiratory defense system is immature. Also they are exposed to greater absorptions of energy per unit mass because their weight is less, and they are exposed to a longer lifetime of exposure because they were exposed from their early life and to a expanded future life period. Consequently, children in this generation will likely have a higher risk of exposure than the previous generations.
To the best of our knowledge, this is the first study showing that EMF can affect the MCT of human respiratory tract mucosa. Future in vivo animal and epidemiological studies might be required to demonstrate the adverse effects of the EMFs and regulatory mechanisms governing these processes.
- 29 -
V. Conclusion
Human nasal mucosa was influenced by EMF. CBF is significantly affected by EMF exposure while MTT assay showed no difference. EMF can decelerate the CBF of cells from the human respiratory tract, which means EMF, in vivo, can slow down mucus clearance and cause respiratory tract disease. We conclude that mobile phone frequency EMF exposure has potentially to alter normal MCT through an nPKC-dependent mechanism. Therefore, care must be taken when EMF exposure is expected on human airway including nasal cavity.
- 30 -
REFERENCES
1. Behari J: Biological responses of mobile phone frequency exposure. Indian J Exp Biol 48(10): 959-981, 2010
2. Braveman I, Wright ED, Wang CG, Eidelman D, Frenkiel S: Human nasal ciliary beat frequency In normal and chronic sinusitis subjects. J Otolaryngol 27:145-152, 1998
3. Clary-Meinesz C, Mouroux J, Cosson J, Huitorel P, Blaive B: Influence of external pH on ciliary beat frequency in human bronchi and bronchioles. Eur Respir J 11:330-333, 1998
4. Duchateau GS, Graamans K, Zuidema J, Merkus FW: Correlation between nasal ciliary beat frequency and mucus transport rate in volunteers. Laryngoscope 95:854-859, 1986
5. Foster KR: Thermal and nonthermal mechanisms of interaction of radio-frequency energy with biological system. IEEE Trans plasma Sci 28:15-23, 2000
6. Fritze K, Sommer C, Schmitz B: Effects of global system for mobile communication (GSM) microwave exposure on blood-brain barrier permeability in rat. Acta Neuropathol(Berl) 94:465-470, 1997
7. Green A, Smallman LA, Logan ACM. The effect of temperature on nasal ciliary beat frequency.
Clin Otolaryngol 20:178-180, 1995
8. Hardell L, Nasman A, Pahlson A, Hallqist A: Case-control study on radiology work, medical x-ray investigations, and use of cellular telephones as risk factors for brain tumors. MedGenMed 4:2:E2, 2000
9. Habash R.W, Elwood JM, Krewski D, Lotz WG, McNamee JP, Prato FS: Recent advances in research on radiofrequency fields and health: 2004-2007. J Toxicol Environ Health B Crit Rev 12:250-288, 2009
10. Heikkinen P, Kosma VM, Alhonen L, Huuskonen H, Komulainen H, Kumlin T, Laitinen JT, Lang S, Puranen L, Juutilainen J: Effects of mobile phone radiation on UV-induced skin tumorigenesis in ornithine decarboxylase transgenic and non-transgenic mice. Int J Radiat Biol 79:221-233, 2003
- 31 -
11. Heitanen M: Health risks of exposure to non-ionizing radiation –myths or science-based evidence. Med Lav 97(2):184-188, 2006
12. Hulme J, Robinson V, DeBie R, Wells G, Judd M, Tugwell P: Electromagnetic fields for the treatment of osteoarthritis. Cochrane Database Syst Rev 1:CD003523, 2002
13. Kim IS, Rhee CS, Lee JH, Heo JH, Park J, Lee CH: Effects of purinergic stimulation on ciliary beat frequency and chloride secretion in sinusitis. Laryngoscope 117:1677-1682, 2007
14. Kim WJ, Han TH, Kim HJ, Park MY, Kim KS, Park RW: An automated measurement of ciliary beating frequency using a combined optical flow and peak detection detection. Healthc Inform
Res 17:111-119, 2011
15. Kobayashi K, Salathe M, Pratt MM, Cartagena NJ, Soloni F, Seybold ZV, Wanner A :
Mechanism of hydrogen peroxide-induced inhibition of sheep airway cilia. Am J Respir Cell Mol
Biol 6:667-673, 1992
16. Kobayashi K, Tamaoki J, Sakai N, Chiyotani A, Takizawa T: Inhibition of ciliary activity by phorbol esters in rabbit tracheal cells. Lung 167:277-284, 1989
17. Kolomyceva MP, Gapeyev AB, Sadovnikov VB, Chemeris NK: Suppression of nonspecific resistance of the body under the effect of extremely high frequency electromagnetic radiation of low intensity. Biofizika 47:71-77, 2001
18. Kovacic P, Somanathan R: Electromagnetic fields: mechanism, cell signaling, other bioprocesses, toxicity, radicals, antioxidants and beneficial effects. J Recept Signal Transduct Res 30:214-226, 2010
19. Kwee S, Raskmark P: Changes in cell proliferation due to environmental non-ionizing radiation, Microwave radiation. Bioelectrochem Bioenerg 44:251-255, 1998
20. Lai H and Singh NP: Single and double strand breaks in rat brain cells after acute exposure to radiofrequency electromagnetic radiation. Int J Radiat Biol 69:513-521, 1996
21. Lansley AB, Sanderson MJ, Dirksen ER: Control of the beat cycle of respiratory tract cilia by Ca2+ and cAMP. Am J Physiol 263:232-242, 1992
22. Maes A, Verschaeve L, Arrovo A, De Wagter C, Vercruyssen L: In vitro cytogenetic effects of 2450 MHz waves on human peripheral blood lymphocytes. Bioelectromagnetics 14: 495-501,
- 32 -
1993.
23. Magras IN, Xenos TD: RF radiation-induced changes in the prenatal development of mice.
Bioelectromagnetics 18:455-461, 1997
24. Nishizuka Y: The molecular heterogeneity of protein kinase C and its implications for cellular regulation. Nature 334:661-665,1988
25. Paredi P, Kharitonov SA, Hanazawa T, Barnes PJ: Local vasodilator response to mobile phones.
Laryngoscope 111:159-162, 2001
26. Persson BRR, Salford LG, Brun A: Blood-brain barrier permeability in rats exposed to electromagnetic fields used in wireless communication. Wireless Network 3:455-461, 1997 27. Phillips JL, Ivaschuk O, Ishida-Jones T, Jones RA, Campbell-Beachler M, Haggren W: DNA
damage in Molt-4 T lymphoblastoid cells exposed to cellular telephone radiofrequency fields in vitro. Bioelectrochem Bioenerg 45:103-110, 1998
28. Repacholi MH: Health risks from the use of mobile phones. Toxicology Letters 120: 323-331, 2001
29. Salathe Matthias: Regulation of mammalian ciliary beating. Annu Rev Physiol 69:401-422, 2007 30. Salathe M, Pratt MM, Wanner A: Protein kinase C-dependent phosphorylation of a ciliary
membrane protein and inhibition of ciliary beating. J Cell Sci 106:1211-1220, 1993 31. Satir P, Sleigh MA: The physiology of cilia and mucociliary interactions. Annu Rev
Physiol ;52:137-155, 1990
32. Satter SA, Islam MS, Rabbani KS, Talukder MS: Pulsed electromagnetic fields for the treatment of bone fractures. Bangladesh Med Res Counc Bull 25: 6-10, 1999
33. Seybold ZV, Abraham WM, Gazeroglu W, Wanner A: Impairment of airway mucociliary transport by Pseudomonas aerogenosa products. Am Rev Respir Dis 146:1173-1176, 1992 34. Seybold ZV, Mariassy AT, Stroh D, Kim CS, Gazeroglu H, Wanner A: Mucociliary interaction in
- 33 -
35. Simet SM, Sisson JH, Pavlik JA, Devasure, JM, Boyer C, Liu X, Kawasaki S, Sharp JG, Rennard SI, Wyatt TA: Long-term cigarette smoke exposure in a mouse model of ciliated epithelial cell function. Am J Respir Cell Mol Biol 43:635-640, 2009
36. Teichtahl H, Wright PL, Kirsner RL: Measurement of in vitro ciliary beat frequency: a television-video modification of the transmitted light technique. Med Biol Eng Comput 24:193-196, 1986
37. Velizarov S, Raskmark P, Kwee S: The effect of radiofrequency fields on cell proliferation are non-thermal. Bioelectrochem Bioenerg 48:177-180, 1999
38. Warwick WJ. Mechanisms of mucous transport: Eur J Respir Dis Suppl 64:162-167, 1983 39. Wong LB and Yeats DB: Dynamics of epithelial function. Comments Theor Biol 4:183-208, 1997 40. Wong LB, Park CL, Yeates DB: Neuropeptide Y inhibits ciliary beat frequency in human ciliated
cells via nPKC, independently of PKA. Am J Physiol 275:440-448, 1998
41. Wyatt TA, Forget MA, Adams JM, Sisson JH: Both c AMP and c GMP are required for maximal ciliary beat stimulation in a cell-free model of bovine ciliary axonemes. Am J Physiol Lung Cell
Mol Physiol 288:546-551, 2005
42. Wyatt TA, Schmidt SC, Rennard SI, Tuma DJ, Sisson JH: Acetaldehyde-stimulated PKC activity in airway epithelial cells treated with smoke extract from normal and smokeless cigarettes.
Proc Soc Exp Biol Med 225:91-97, 2000
43. Wyatt TA, Spurzem JR, May K, Sisson JH: Regulation of ciliary beat frequency by both PKA and PKG in bovine airway epithelial cells. Am J Physiol 275:827-835, 1998
44. Yager J, Chen TM, Dulfano MJ: Measurement of frequency of ciliary beats of human respiratory epithelium. Chest 73:627-633, 1978
45. Ye J, Yao K, Lu D, Wu R, Jiang H: Low power density microwave radiation induced early changes in rabbit lens epithelial cells. Chin Med J 114:129-134, 2001
46. Yoon JH, Kim KS, Kim SS, Lee JG, Park IY: Secretory differentiation of serially passaged normal human nasal epithelial cells by retinoic acid: expression of mucin and lysozyme. Ann
- 34 - - 국문 요약 -