저작자표시-비영리-변경금지 2.0 대한민국 이용자는 아래의 조건을 따르는 경우에 한하여 자유롭게 l 이 저작물을 복제, 배포, 전송, 전시, 공연 및 방송할 수 있습니다. 다음과 같은 조건을 따라야 합니다: l 귀하는, 이 저작물의 재이용이나 배포의 경우, 이 저작물에 적용된 이용허락조건 을 명확하게 나타내어야 합니다. l 저작권자로부터 별도의 허가를 받으면 이러한 조건들은 적용되지 않습니다. 저작권법에 따른 이용자의 권리는 위의 내용에 의하여 영향을 받지 않습니다. 이것은 이용허락규약(Legal Code)을 이해하기 쉽게 요약한 것입니다. Disclaimer 저작자표시. 귀하는 원저작자를 표시하여야 합니다. 비영리. 귀하는 이 저작물을 영리 목적으로 이용할 수 없습니다. 변경금지. 귀하는 이 저작물을 개작, 변형 또는 가공할 수 없습니다.
이학석사학위논문
초파리의생체시계에서CLOCK의
PERIOD 결합 도메인의역할
아주대학교대학원
의생명과학과/신경과학전공
강두현
초파리의생체시계에서CLOCK의
PERIOD 결합 도메인의역할
지도교수김은영
이논문을이학석사학위논문으로제출함.
2012년8월
아주대학교대학원
의생명과학과/신경과학전공
강두현
강두현의이학석사학위논문을인준함.
아주대학교대학원
2012년06월22일
- 국문요약 -
초파리 생체시계에서 CLOCK 의 PERIOD 결합 도메인의 역할
생체시계의 작동 기전은 Transcriptional/Translational Feeldback Loop (TTFL)이다. 초파리에서는 전사인자인 CLOCK (dCLK)과 CYCLE (CYC)가
결합하여 period (per)와 timeless (tim)의 발현을 촉진하고, PER 와 TIM
단백질은 결합하여 CLK/CYC 의 활성을 억제함으로써 자신의 발현을 저해하며 하는 되먹임 고리가 TTFL 의 핵심 고리를 이룬다. PER/TIM 이합체에서 PER 가 CLK/CYC 의 활성을 억제하는 역할을 하는데 어떤 생화학적 기전을 통하여 억제하는가에 대한 이해가 많이 부족하다.본 연구에서는 CLK 의 아미노산을 각각 100~200 개 정도의 단위로내부적으로 제거하여 변이체를 만들고 PER 가 결합하는 도메인을 찾고자 하였다.Schneider2 (S2) 초파리 세포라인에서 각각의 CLK 변이체들과 PER 와의 결합정도를 면역침강법으로 비교한 결과 아미노산 657-707 이제거된 dCLK (dCLKD657-707 로 명명)이 PER 와 결합 정도가 감소하는 것을 관찰하였다. 흥미롭게도 S2 세포에서 CLKD657-707 은 E-box 의존적인 전사활성을 나타내지 못하였다. 그러나 dCLKD657-707 이 CYC 단백질과는 정상적으로 결합하는 것으로 보아 아미노산 657-707 의 제거로 인한 구조 변화와 같은 비특이적 결함에 의해서가 아니라 아미노산 657-707 부위가 dCLK 단백질의 전사활성 및 PER 단백질과의 결합에 필요한 도메인임을 의미하는 것이다.아미노산 657-707 의in vivo 역할을 탐색하기 위하여 dCLK△657-707 을 발현시키는 초파리를
제작하고 dCLK 을 발현하지 못하는 ClkOUT 유전적 배경 (dClk△657-707;ClkOUT 으로 명명)에서 분자적•행동학적 리듬을 관찰하였다.dClk△657-707;ClkOUT 초파리는 생체리듬을 나타내지 못하여 초파리 세포라인에서 얻은 결과와 마찬가지로 아미노산 657-707 부위가 CLK 이 생체시계를 작동시키는 데 있어 중요한 역할을 하는 도메인임을 입증할 수 있었다. 흥미롭게도 S2 에서 dCLK△657-707 은 전사활성을 전혀 나타내지 못하였는데, 초파리에서는 PER 와 TIM 단백질이 정상 초파리에서 보다는 약간 낮은 정도로 만들어지는 것으로 확인되어 이에 대한 후속 연구가 진행되어야 할 것으로 보인다. 핵심어 :초파리, 생체시계, dCLOCK,dPERIOD, 전사활성, 결합도메인
차
례
국문요약 ··· iv 차례 ··· vi 그림차례 ··· viii 표차례 ··· x Ⅰ. 서론 ··· 1 Ⅱ. 재료 및 방법 ··· 4 A. plasmid ··· 4 1. S2 에서 발현시키기 위한 plasmid 들 ··· 4 (1) plasmid 종류···4(2) Site-directed mutagenesis 를 위한 primer···4
(3) Site-directed mutagenesis···6
2. 형질전환 초파리 제작을 위한 plasmid ··· 8
B. Schneider 2 (S2) 세포의 배양과 transfection ··· 9
C. 형질전환 초파리 (Transgenic Drosophila melanogaster)···10
D. Locomotor activity assays···11
E. Immunoblot analysis···12
F. Immunoprecipitation(IP) ···15
G. Luciferase Reporter Assay 와 β-Galactosidase Enzyme Assay System···18
Ⅲ. 결과 ···21 A. S2 세포에서 dCLK(△579-778) 은 dPER 가 결합 하지 못하지만 CYC 단백질의 결합에는 영향이 없다.···21 B.S2 세포에서dCLK(△657-707)은dPER와의 결합을 하지못한 다.···25 C.dCLK(△657-707) 을 발현하는 형질전환 초파리는 정상적인 생체리듬을 가지지 못한다.···29 Ⅳ. 고찰 ···38 Ⅴ. 결론 ···40 참고문헌···42 ABSTRACT ···46
그림 차례
Fig. 1.Schematic diagram of internally deleted amino acid region on dCLK protein···23
Fig. 2.Interaction between PERIOD with CLOCK internal deletion mutants···24
Fig. 3.Interaction between CYCLE with CLOCK internal deletion mutants···25
Fig. 4.Amino acid 657-707 is necessary for PERIOD binding to CLOCK···27
Fig. 5.CLOCK△657-707 does not have transcriptional
activity···28
Fig. 6.Amino acid 657-707 is not necessary for CYCLE binding to CLOCK···29
Fig. 7.Daily activity patterns of Clk-WT; ClkOUT
and Clk△657-707; ClkOUT···33
Fig. 8.Amino acid 657-707 is necessary for PERIOD binding to CLOCK at in vivo···34
Fig. 9.Molecular rhythms of Clk-WT; ClkOUT andClk△657-707;ClkOUT
during Light/Dark Cycle···35
Fig. 10.Molecular rhythmsofClk-WT; ClkOUT andClk△657-707;ClkOUT
during Light/Dark Cycle···36
Fig. 11.Daily levels of tim and clk mRNAsClk-WT; ClkOUT
표 차례
Table 1. Circadian behavior of Clk-WT; ClkOUT andClk△657-707;ClkOUT
I. 서
론
지구상에 존재하는 모든 생명체는 지구의 자전으로 변화하는 환경을 미리 예측할 수 있다. 예를 들면 포유류의 경우 뇌의 시상하부에 위치한 SCN(Suprachiasmatic Nuclei) 이 낮과 밤을 예측하고 각 장기의 다양한 세포로 필요한 정보를 보내어 하루 중 대사작용, 체온, 호르몬분비, 수면 등을 조절한다. 이로 인해 하루를 주기로 생명체의 여러 생리작용과 행동 및 대사 패턴을 변화시키는 생체리듬이 나타나게 된다. 생체시계와 질병과의 상관관계는 암, 수면장애, 여러가지 형태의 우울증, 신경퇴행성 질환, 대사증후군, 약물중독 등 무수히 많은 예로부터 중요성이 입증되고 있다(Bunney 와 Bunney,2000; Cardinali, 2000; Fu 와 Lee, 2003; McClung, 2007; Rudic 등 , 2004). 따라서 생체시계의 작용기전을 밝혀 생체리듬을 일으키는 원인과 비정상적 생체리듬이 발생함으로 인해 어떠한 질병이 나타나며, 예방과 치료를 위한 수단과 방법을 탐색하는데 중요한 단서가 될 것이다. 생체시계는 미생물에서부터 포유류까지 지구상의 모든 생물이 가지고 있는데 기원전부터 과학자들은 생물내부에 자체적인 시계를 가지고 있을 것이라고 생각하고 있었으며, 이에 관한 연구는 초파리를 동물모델로 이용하면서 활발히 진행되었다.생체시계의 핵심기전은 “interlocked transcriptional-translational feedback loop” 으로써 두 개의 loop 이 맞물려 생체시계의 핵심 유전자들의 발현이 하루를 주기로 진동할 수 있도록 한다. 초파리에서는 dperiod(dper) / timeless (tim) loop 이 두개의 feedback loops 을 이루고promotor 부분의 E-box 에 붙어 전사를 촉진하고 dPER/TIM 단백질은 이형이합체를 이루어서 자신의 유전자를 만들게 하는 dCLK/CYC 단백질의 전사활성을 저해하는 되먹임 고리를 만들게 된다. 또한 dCLK/CYC 단백질의 이형이합체는 vrille (vri )과 PAR domain protein 1ε (PDP1 ε) 유전자의
E-box 에 결합하여 이들의 발현을 촉진하는데, 이 때 VRILLE (VRI) 은 dClk 의
발현을 억제하고 반면 PDP1 ε 은 dClk 의 발현을 촉진하는 또 다른 feedback loop 를 이루게 된다(Cyran 등 , 2003).
생체시계의 “interlocked transcriptional-translational feedback loop” 에서 dPER 와 TIM 의 이형이합체가 dCLK 과 CYC 이형이합체의 활성을 억제한다는 것은 잘 알려져 있다. 이때 dPER-TIM 이형이합체 단백질이 전사인자인 dCLK-CYC 이형이합체 단백질을 억제하기 위해 직접적인 결합이 필요할 것이며 (Lee 등 , 1998; Lee 등, 1999; Sato 등 , 2006), Chang 등은 dPER 가 단독으로도 dCLK-CYC 의 활성을 저해할 수 있음을 보고하여
dCLK-CYC 이합체의 활성을 저해하는 주요 단백질은 dPER 임을
밝혔다(Chang 과 Reppert, 2003). Chang 등은 이 연구에서 dPER 에서 dCLK 의 활성을 억제하는데 필요한 최소 도메인을 탐색하였고, 그 결과 dPER 에서 아미노산 764-1034 가 내부적으로 제거 되었을 때 dCLK 의 전사활성이 억제되지 않았고 이로부터 Chang 등은 dPER 의 아미노산 764-1034 를 dCLK:CYCInhibition Domain(CCID)라 명명하였다(Chang 과 Reppert, 2003).
여러 초파리 종으로부터 CCID domain 의 단백질 서열을 비교 분석해 보면 종간 보존성이 높은 부위 (Conserved region) 와 종간 보존성이 상대적으로 높지 않은 부위 (Non-conserved region)로 나뉘는 것을 알 수 있다. 특히 dPER 에서 종간 보존성이 높은 부위인 C3 부위(아미노산 768-842)는 DBT 와
결합하는 부위라는 것을 지도교수의 이전 연구에 의해 밝혀 졌고 C4 부위(아미노산 926-977)도 선임연구자에 의해 dCLK 과 결합하는 부위라는 것이 밝혀졌다.(Kim 등,2007 ; Sun 등,2010)
따라서 본 연구에서는 생체시계의 “interlocked transcriptional-translational feedback loop” 에서 dCLK 의 전사활성을 저해하기 위해 dPER 단백질이 dCLK 에 결합하는 부위를찾고 이 도메인의 기능을 분자적 • 생화학적 관점에서 탐구하고자 한다.
Ⅱ. 재료 및 방법
A. plasmid
1. S2에서 발현시키기 위한 plasmid 들
(1) plasmid 종류
pMT(A)-dClk△DBD-V5,pMT(A)-dClk△HLH-V5,pMT(A)- dClk△63-95-V5,pMT(A)-dClk△PAS-A-V5,pMT(A)-dClk△145- 263-V5,pMT(A)-dClk△PAS-B-V5,pMT(A)-dClk△310-378-V5,pMT(A)-dClk△379-479-V5, pMT(A)-dClk△480-578-V5, pMT(A)-dClk△579-656-V5,pMT-dClk△579-778,pMT-dClk△657-707,pMT-dClk△708-778, pMT-dClk△784-834,pMT-dClk△837-945(2) Site-directed mutagenesis를 위한 primer
(A) clk△DBD
Forward :5’- GAC AAG GAT GAT ACA TTC AAC TCG CTG GTC -3’ Reverse :5’- GAC CAG CGA GTT GAA TGT ATC ATC CTT GTC -3’ (B) clk△HLH
Forward :5’- AAG CGA CGA GAT CAG AAA AAT CAT AAT GAG -3’ Reverse :5’- CTC ATT ATG ATT TTT CTG ATC TCG TCG CTT -3’ (C) clk△63-95
Forward :5’- ACG ATA GCC TTC CTG CTG GAG TCT CTC GAT -3’ Reverse :5’- ATC GAG AGA CTC CAG CAG GAA GGC TAT CGT -3’ (D) clk△PAS-A
Forward :5’- TAC ACT CAC CTC ATG GAG GCG CTG CTG AAT -3’ Reverse :5’- ATT CAG CAG CGC CTC CAT GAG GTG AGT GTA -3’ (E) clk△145-263
Forward :5’- TAT GAG ATG GAC CAT AGT AAT GAG TTC ACT -3’ Reverse :5’- AGT GAA CTC ATT ACT ATG GTC CAT CTC ATA -3’ (F) clk△PAS-B
Forward :5’- ATT ATT GAT CCC ACA GAC AGT ATT GTG GCG -3’ Reverse :5’- CGC CAC AAT ACT GTC TGT GGG ATC AAT AAT -3’ (G) clk△310-378
Forward :5’- CAC TTT GAT GAT CTG GGC CAA AAA TCG GGA -3’ Reverse :5’- TCC CGA TTT TTG GCC CAG ATC ATC AAA GTG -3’ (H) clk△379-479
Forward :5’- GAT AGT CGA AAG GAG AAT ATC AGC TCC ACA -3’ Reverse :5’- TGT GGA GCT GAT ATT CTC CTT TCG ACT ATC -3’ (I) clk△480-578
Forward :5’- GCT TCG AGT TAT GGC ACT CCG AAA ATG GTG -3’ Reverse :5’- CAC CAT TTT CGG AGT GCC ATA ACT CGA AGC -3’ (AJ) clk△579-778
Reverse :5’- GAG ATT GTG CTG GTG CAC TGT ATG TTG CAG -3’ (K) clk△579-656
Forward :5’- CTG CAA CAT ACA GTG CC GAT ACG GTG GTT -3’ Reverse :5’- AAC CAC CGT ATC GGG CAC TGT ATG TTG CAG -3’ (L) clk△657-707
Forward :5’- CAA ACG GAT ATG CTG CCC ATG ATG TCG ATG -3’ Reverse :5’- CAT CGA CAT CAT GGG CAG CAT ATC CGT TTG -3’ (M) clk△708-778
Forward :5’- TAC ACA TAT CTG CAG CAC CAG CAC AAT CTC -3’ Reverse :5’- GAG ATT GTG CTG GTG CTG CAG ATA TGT GTA -3’ (N) clk△784-834
Forward :5’- CAC CAG CAC AAT CTC CAA AAT GAT ATT TTA -3’ Reverse :5’- TAA AAT ATC ATT TTG GAG ATT GTG CTG GTG -3’ (O) clk△837-945
Forward :5’- CAA AAT GAT ATT TTA AAT CTG GTG CAG CAG -3’ Reverse :5’- CTG CTG CAC CAG ATT TAA AAT ATC ATT TTG -3’
(3)Site-directed mutagenesis
QuickChange site-directed mutagenesis kit (Stratagene Co., La Jolla, CA,U.S.A) 을 사용하여 site directed mutagenesis를 수행하였다. 이 때 pfuTurbo DNA polymerase (Stratagene Co., La Jolla, CA, U.S.A)를 이용
(Stratagene Co., La Jolla, CA, U.S.A) 가 제시하는 방법을 사용하였고 PCR
반응 혼합액 구성요소는 100 ng 의 pMT-HAdClk-V5, 5 ㎕ 의 10X cloned
Pfu reaction buffer , 2 ㎕의 10mM dNTP, 각각 150 ng 의 primer (F)(R),
1㎕ 의 Pfu Turbo DNA polymerase (Strata gene Co., La Jolla, CA,U.S.A)
그리고 멸균수로 최종반응 양을 50 ㎕로 맞추어 조제하였다. PCR은 핵산증폭기
(MJ Research, Inc., Waltham, MA, U.S.A)를 이용하였다. PCR 산물 중
mutation되지 않은 주형 DNA를 제거 하기 위해 제한효소 Dpn I 1 ㎕를 처리한
후 37℃에서 1 시간 반응 후 , XL10-Gold Ultracompetent cells (Stratagene Co., La Jolla, CA,U.S.A) 에 42 ℃로 45초간 heatshock을 주고 난 후, 1 시간 30 분 동안 shaking incubator에서 배양하여 transformation 하였다. 재조합 plasmid를 가진 콜로니 임을 확인하기 위해 sequencing을 보내 염기서열을 확인 하였다.
2. 형질전환 초파리 제작을 위한 plasmid
AttbpAcman-dClk-V5이plasmid의NcoI-SphI단편을 pSP72
vector(promega Co., Madison, WI, U.S.A) 에subcloning하였다.이 plasmid 를 pSP72-dClk (NcoI/SphI) 로 명명하였다. site-direct mutagenesis를수행 후 pSP72-dClk△657-707(NcoI/SphI)를 제작하였다. 아미노산 657-707 의
제거는 DNA sequencing
analysis를통해확인하였다.pSP72-dClk△657-707(NcoI/SphI)에서 NcoI 과 SphI 제한효소를 이용하여 dClk△657-707을 잘라낸 후 pSP72-dClk(NheI-SphI)에subclonning 하여 pSP72-dClk△657-707(NheI/SphI)를 제작하였다. 이렇게 준비된 plasmid를 attbpAcman-dClk
vector 에NheI-SphI을 이용하여 subcloning 함으로써 최종적으로
B. Schneider 2(S2) 세포의 배양과 transfection
S2 세포는 10% Fetal Bovine Serum, Certified(FBS) (Invitrogen Co., Carlsbad, CA, U.S.A) 와 0.5% penicillin streptomycine (Sigma-Aldrich Co., Carlsbad, CA, U.S.A)를 사용하였고 23℃ 항온기에서 배양하였다. Transfection은 Effectene Transfection reagent (Qiagen Co., Hilden, Germany)를 사용하였고 제조사가 제시한 실험 방법에 따라 실험하였다. 각 transfection 은 각각 다른 0.4 ㎍ 의 pMT-HAdClk 을 포함한 plasmid 와 0.6 ㎍ 의 pAct-per 또는 각각 다른 0.5 ㎍ 의 pMT-HAdClk 을 포함한 plasmid 와 0.5 ㎍ 의 CYC-3F 를 사용하였다. pAct-per와 pMT-cyc-3F는 transfection 후 최종 500 uM CuSO4 를 넣어 induction 함으로써
C. 형질전환 초파리 (Transgenic Drosophila melanogaster)
이 pAcman-dClkD657-707 plasmid를 BestGeneInc (BestGene Inc., Chino Hills, CA, U.S.A)에 보내 형질전환 초파리 제작을 의뢰하였다. 이 때 pAcman-dClkD657-707 plasmid를 주입하는 초파리 라인은 VK00018을 이용하였다. 그 결과 4개의 독립적 라인을 얻었고 각각의 초파리를 Clk 을 발현하지 않은 ClkOUT 초파리와 교배하여 유전적 배경에서 ClkD657-707 을 발현하는 초파리 라인을 확립하였다. 초파리의 유전형은 다음과 같다. W; ClkD657-707,4M;ClkOUT W; ClkD657-707,3M;ClkOUT W; ClkD657-707,2M;ClkOUT W; ClkD657-707,1M;ClkOUT
D. Locomoter activity assays
초파리 각각의 locomotor activity 는 Drosophila Activity System (DAM)을 이용하여 측정하였다. (Waltham MA) (Hemblen-Coyle 등, 1992). 측정을 위해 초파리는 어린 수컷 성체를 사용하였다. 25℃ 항온기에서 12시간 간격으로 빛과 어둠속에 총 4일 간 노출하였고 (12:12LD, zeitgeber time zero [ZT0] sms light phase 가 시작되는 시점으로 정의를 내린다.), Constant-darkness condition (DD)을 12:12LD 이후 연속해서 7일간 유지하였다. 각 초파리의 Locomotor activity 는 Dr. F. Rouyer. (France)로부터 받은 Faax software (MacOSX 를 위한 Fly Activity Analysis Suite) 를 이용하여 분석하였다. Period 는 초파리 각각을 chi-square periodogram analysis 하여 , 각 유전형의 그룹별 평균을 산출하여 그 값을 획득하였다. power는 DD동안 rhythm의 strength의 정도이다. 각각의 초파리들은 power 가 10 이상의 값을 가지고 width 값 (periodogram 95% confidence line 보다 30분 위에 peak된 수를 표시 ) 이 2 이상의 rhythm을 나타낸다.
E. Immunoblot analysis
전기영동용 Gel은 6% polyacrylamide gel (Resolving gel mix; 1.2 ㎖ 의 30% polyacrylamide/Bis solution,37.5:1(BIO-RAD),2.325 ㎖의 DW, 2.4 ㎖ 의 1M Tris-Hcl pH 8.8, 60㎕ 의 10% SDS, 25 ㎕ 의 25% APS, 6㎕ 의 TEMED) (Stacking gel ; 375 ㎕ 의 30% Acrylamide Bis,37.5:1,BIO-RAD 1.85 ㎖ DW, 250 ㎕ 의 1M Tris-Hcl pH 6.8, 25 ㎕ 의 10% SDS, 10 ㎕ 의 25% APS, 3㎕ 의 TEMED)을 제작하여 사용했다. 시료의 전 처리를 위해 S2 세포를 거두어 들인 뒤 PBS로 한 번 씻어 준 후 150 ㎕ 의 Modified RIPA(50mM Tris-Hcl pH 7.5, 1% NP-40, 0.25% deoxycholate, 50mM Nacl, 5% glycerol) 에 신선한 1 mM EDTA, 100X Xpert protease inhibitor cocktail
solution(GenDEPOTCo., U.S.A), 2.5M 의 NaF , 200mM Na3VO4, 0.5mM의
PMSF 을 추가하여 세포를 용해 하였다. 용해 방법은 얼음 위에서 5 분 마다 10초씩 세 번 voltexing 하여 총 15분간 용해한 후 4℃ 에서 13000 rpm 으로 15분간 원심 분리하여 상층액 만을 회수하였다. 회수한 각 세포 용해액은 ELIZA leader (BioTex Inc., Houston, TX, U.S.A) 를 이용하여 595 nm 로 농도를 측정하여 OD3과 OD6 으로 정량한 후 단백질을 변성시키기 위해 4X SDS-PAGE sample buffer(2.5㎖ 1M Tris-Hcl pH 6.8, 4㎖ 의
Glycerol(100%) , 0.8㎖ β-mercaptoethano, 0.8g SDS, 2.5㎖ Autoclaved
water )를 추가하여95℃ 이상의 온도에서 3 분간끓인후spindown하여 sample 들을 loading 하였다.준비된 sample들은 SDS-PAGE 전기영동법 (Sodium Dodecyl Sulfate – Polyacrylamide gel Electrophoresis, Laemmli법) 을
수행하였다. Mini-PROTEIN Tetra Cell (BIO-RAD Laboratories, Inc., Hercures, CA, U.S.A) 을 이용하였고, 전기영동용 buffer (1 L ; 3.025 g Tris, 14.4 g Glycine, DW, 1% SDS)를 채운 뒤 전기영동 하였다. Stacking gel에서는 130 V 로 Resolving gel에서는 190 V 로 설정 하였고 PRO-STAIN Prestained protein marker (Intron Biotechnology Inc., Korea) 을 함께 loading 하여 marker의 90.5 kDa 밴드가 gel의 가장 하단에 위치할 때까지 전기영동 하였다. 전기영동 완료된 gel은 resolving gel 만을 잘라낸 후 transfer buffer 에 충분히 적신 Gel Blot Paper (Whatman Inc., Springfield
Mill, Kent, U.K) 와 Immobilon-P Transfer Membrane(Millipore
Inc.,U.S.A)를 함께 dry blot sandwich를 만들어 Trans-Blot SD semi-dry electrophoretic transfer cell (BIO-RAD Laboratories Inc., Hercures, CA, U.S.A)에 함께 장착한 다음 240mA 에서 45분간 transfer 하였고, Transfer 된 nitrocellulose transfer membrane 은 0.1% Ponceau S(0.1% Ponceau S, 5% acetic acid) 로 확인 하였다. Orbital shaker 위에서 5% Blocking 하였고, 1차 항체가 포함된 5% Blocking 용액에 넣어 냉장실에서 하룻동안 반응 시킨 뒤 , 2차 항체가 포함된 5% Blocking 용액에 넣어 상온에서 세 시간 동안 반응하였다. 이 때 Anti-V5(Invitrogen Co., Carlsbad, CA, U.S.A) 는 1:10000, Anti-HA(3F10)(Roche Diagonostics Co., Mannheim,
Germany) 은 1:4000으로 희석하여 사용하였고, Goat anti-mouse
IgG(H+L)horseradish peroxidase(molecular Probes Inc., Eugene, OR, U.S.A)는 1:10000,Goat anti–rat IgG (H+L), (molecular Probes Inc., Eugene,
OR, U.S.A)는 1:5000으로 희석하여 사용하였다. 항체 반응이 끝난 뒤에는 항상 15분 간격으로 4번 TBST(4M NaCl, 1M Tris-HCl pH 7.5, 0.05% Tween-20, DW)로 세척하였다. 항체반응이 끝나면 Immobilon Western chemiluminescent HRP substrate(Millipore Co., Billerica, MA, U.S.A)를 nitrocellulose transfer membrane 에 적절히 묻힌 후 CP-BU NEW medical X-ray film blue (Agfa HealthyCare Co., Soptestraat, Mortsel,Belgium) 에 적정시간 동안 노출시켜 현상하였다.
F. Immunoprecipitation(IP)
pMT-HAdClk또는 pMT-HAdClk(△) series 0.4㎍ 과 함께 pAct-dper-V5/His(0.6㎍) ,pMT-HAdClk또는 pMT-HAdClk(△) series 0.5㎍ 과 함께 pMT-cyc-3F(0.5㎍)을 S2 세포에 Effectene Transfection reagent (Qiagen Co., Hilden, Germany) 을 사용하여 제조사의 설명에 따라 transient
transfection 하였다. 36 시간 동안 배양한 뒤 최종 500 uM CuSO4 를 넣어
dClk의 발현을 유도한 후 24 시간 동안 배양하였다. 세포를 수확하고 PBS로 한 번 씻어준 후 Modified RIPA (50mM Tris-Hcl pH 7.5, 1% NP-40, 0.25% deoxycholate, 50 mMNaCl, 5% glycerol) 에 신선한 0.5mM PMSF, , 100X Xpert protease inhibitor cocktail solution(GenDEPOT Co., U.S.A), 2.5M 의
NaF , 200mM Na3VO4,1 mM EDTA 를 추가한 lysis buffer 를 이용하여
세포를 용해하였다. 용해 방법은 얼음 위에서 5 분마다 10 초씩 세 번 voltexing 하여 총 15 분간 용해한 후 13000 rpm으로 15 분간 4℃ 원심 분리 하여 상층액만을 회수하였다. 회수한 세포용해액은 ELIZA leader (BioTex Inc., Houston, Tx, U.S.A) 를 이용하여 595nm 에서 O.D값을 측정한 후 단백질 양을 동일하게 맞추고 각각 dPER에 대한 anti-V5 항체 (Invitrogen Co., Carlsbad, CA, U.S.A) 또는 dCLK 에 대한 Anti-HA (12CA5) 항체 (Roche Diagonostics Co., Mannheim, Germany)를 3㎕ 씩을 넣어준 다음 4℃ 냉장실에서 하루 동안 lotation 하였다. 다음날 GammaBind G sepharose (GE Health care Bio-Sciences AB.,Bjorkgatan, Uppsala, Sweden) bead 를 사용할 만큼 미리 준비하여 세포용해시 사용했던 동일한 buffer (Modified-RIPA) 로 약하게 세 번 세척 한 후. 각 sample 마다 준비한 bead를 25㎕
씩넣고 4℃ 에서 세 시간 동안 lotation 하였다. Sample 들은 4℃ 에서 2분 동안 원심 분리한 후 상층액은 따로 모아두고, bead를 다시 4℃에서 5분간 lotation 하여 buffer(Modified-RIPA) 로 세척하는 과정을 세 번 반복한 후 buffer (Modified-RIPA) 를 최대한 제거한다. 1.2X Sample buffer 를 각 sample 마다 30㎕ 씩 넣고 95℃ 에서 5 분간 끓인 후 15 ㎕ 씩 loading 하여 immunoblottung 을 수행하였다. Immunoblotting 에는 6% polyacrylamide gel 을 사용하였고, IP 수행 시 3F10(Roche Diagonostics Co., Mannheim, Germany) 로 dCLK을 검출한 sample 에는 dPER를 찾기 위해 1 차 항체로 Anti-HA(12CA5)(Roche Diagonostics Co., Mannheim, Germany) 로 dCLK 을 검출한 sample 에는 dPER를 찾기 위해 Anti-V5 (incitrogen Co., Carlsbad, CA, U.S.A) 항체를 사용하였다. 또한 원래의 세포 용해액은dCLK과 dPER 둘 다 O.D 10 으로정량하여 함께 loading 하여 줌으로써 세포 내 존재하는 각 단백질 양과 dPER 또는 dCLK 의 각각의 결합도를 비교하였다.
이것과 함께 dClk과 CYC는EZ view tmRed ANTI-FLAG M2 Affinity
gel( sigma-Aldrich, Inc., Missouri 63103 U.S.A)bead 를 각 sample 마다 25㎕ 씩 넣고 4℃ 냉장실에서 하루 동안 lotation 하였다.Sample 들은 4℃ 에서 2분 동안 원심 분리한 후 상층액은 따로 모아두고, bead만 있는 곳에 pH 2.7 의 0.1M Glycine-Hcl 을 27㎕ 을 넣고 10분동안 room temperature 에서 incubation 한다. 10분간의 incubation 동안 2분마다 tapping 을 한 다음 4℃ 에서 2분 동안 원심 분리한 후 상층액은 새로운 e.tube 에 옮긴다. 이때 옮기는 e.tube 에는 3㎕의 pH 8.8 tris-Hcl 을 미리 넣어 둔다. 총 30 ㎕ 의 sample 이 들어있는 e.tube에 10㎕의 4X sample buffer 를 넣고 95℃ 에서 3
수행하였다.Immunoblotting 에는 8% , 10% polyacrylamide gel 을 사용하였고, IP 수행 시 Anti-FLAG( sigma-Aldrich, Inc., Missouri 63103 U.S.A)으로 CYC 를 검출한 sample 에는 dCLK 을 찾기 위해 1 차 항체로 Anti-HA(12CA5)(Roche Diagonostics Co., Mannheim, Germany) 로 dCLK 을 검출한 sample 에는 CYC를 찾기 위해 Anti-FLAG(Monoclonal ANTI-FLAG M2) ( sigma-Aldrich, Inc., Missouri 63103 U.S.A) 항체를 사용하였다. 또한 원래의 세포 용해액은dCLK 은 O.D 8, CYC 는 O.D 3 으로 정량하여 함께 loading 하여 줌으로써 세포 내 존재하는 각 단백질 양과 CYC 또는 dCLK 의 각각의 결합도를 비교하였다.
G. Luciferase Reporter Assay 와 β-Galactosidase Enzyme
Assay System
Assay를 위해서 먼저 세포 용해액을 준비해야 한다. 24well plate에 S2 세포를 배양한 후 dclkE-Luc, pMT-dclk-V5/His 와 pMT-dclk△ 657-707 - V5/His 를 Effectene Transfection reagent (Qiagen Co., Hilden, Germany ) 을 사용하여 제조사의 설명에 따라서 transfection 하였고, 모든 sample의 transfection 효율을 동일화 시키기 위해 pMTB로 보정하였으며 오차범위를 줄 이기 위해 triplecation 하였다. 36 시간 후 최종 500mM CuSO4로 induction 한 다음 12시간 후 well을 PBS로 두 번 세척하였다. 5X RepoterLysis Buffer (RLB)(Promega Co., Madison, WI, U.S.A) 를 1X로 희석하여 각 well의 표면 을 충분히 덮을 수 있을 정도로 150㎕ 를 분주 하였고 , 4℃ shaker 로 강하게 흔들면서 20 분 가량 incubation 하면서 세포를 최대한 용해 시켰다. 세포 용 해액은 4℃에서 13000rpm 으로 5 분간 원심분리하여상층액만을 회수하였다.
Luciferase repoter assay 를 위해서 상온에서 Luciferase Assay
Reagent(LAR) )(Promega Co., Madison, WI, U.S.A) 를 녹인 다음 1.5 ㎖ microcentrifuge tube 에 100㎕ 의 LAR 와 세포용해액을 넣어 잘 섞어서 delay 는 3초 , integration time 은 15초, replication 은 1로 설정된 Luminometer (Turner Designs Inc., Sunnyvale, CA, U.S.A) 에서 측정하였다.
이 때 세포용해액 중 dClk△ 657-707 이 아닌 dClk을 2 ~ 10 ㎕ 을 넣어 측
정하였을 경우 측정값이 약 3000 이 나올 때 적절하다. β-Galactosidase Enzyme Assay 를 위해서 96 well plate 에 각 sample 20 ㎕ , 1X RLB 30
㎖ ONPG) 50 ㎕ 를 넣고 37℃ 에서 노란색으로 변할 때까지 약 30 분 정도 반응시킨 후 150 ㎕ 의 1 M Sodium Carbornate 를 넣어 반응을 중단시키고 즉시 ELIZA leader (BioTex Inc., Houston, TX, U.S.A) 를 이용해 420 nm 의 파장으로 측정하였다. β-Galactosidase Enzyme Assay 값을 이용하여 Luciferase Reporter Assay 값을 보정하여 graph를 작성하였다.
H. Quantitative Real-time RT-PCR
초파리 머리를 TRI reagent(Molecular Research center, Inc)로 용해하여 RNA를 획득한다.이렇게 얻은 총 500ng 의 RNA 를 oligo-dT primer 와 amfiRivert reverse transcriptase (GenDEPOT)를 이용하여 cDNA를 합성한다. cDNA를 주형으로 Quantitect SYBR Green PCR kit (Qiagen)을 이용하여 Corbett Rotor Gene 6000 instrument (Corbett Life Science) 에서real-time
RT-PCR을 수행하였다.primer sequence는 dClkforward : 5´
CAGTTCAACTCGCTGGTCAA-3´; dClk reverse : 5´
TCAGGAAGGCTATCGTGGAC-3´timforward: 5´-CCCT
ATACCCGAGGTGGAT-3´;timreverse:5´-TGATCGAGTTGCAGTGCTTC-3´.여기에 mRNA가 cycling 하지 않는 CBP20 를 첨가하여 값을 보정하였다(Majercak et al. 2004). CBP20 의
sequence 는 다음과 같다.CBP20 forward:
5´-GTCTGATTCGTGTGGACTGG-3´; CBP20 reverse:
5´-CAACAGTTTGCCATAACCCC-3´. 수행 후 나온 결과를Roter Gene 6000
software 를 이용하여분석하였다.그리고 상대적인 mRNA level을 2-△△Ct방법을
III. 결과
A. S2 세포에서 dCLK(△579-778) 은 dPER 가 결합 하지 못
하지만 CYC 단백질의 결합에는 영향이 없다.
최근 우리 연구실에서는, dCLK 의 단백질에서 전사 억제제인 dPER가 결합 하는 부위인 아미노산 579-778을 dCLK에서 제거하게 되면 dPER 단백질이 결합하지 못한다는 연구결과를 얻게 되었다.(Fig.1) 먼저 dCLK(△579-778) 단백질이 partner 단백질로 알려진 CYC 단백질과의 결합이 되지 않기 때문일 가능성을 먼저 조사해보았다. dCLK full length domain을 12가지 종류로 부분적 으로 아미노산을 내부적으로 삭제를 시켜 dCLK 단백질에 dPER 단백질이 결 합하지 못하는 부위를 찾아 냈다. 그리고 dPER 단백질이 결합하지 못하는 부위 에 partner 단백질인 CYC 단백질과의 결합에는 어떤 영향을 미치는 지를 immunoprecipitation(IP)를 통해 알아보았다. dCLK(△579-778)는 dPER 단 백질이 결합하지 않았지만 dCLK 단백질과 이형접합체를 형성하는 partner 단백 질인 CYC 의 경우dPER 단백질과는 다르게 결합하는데 아무 문제가 없었다. 이 와 같은 실험을 진행하면서 얻게된 또다른 실험결과는 dCLK 과 CYC 가 결합 하는 부위는 PAS-A 가 중요한 역할을 할 것이라는 실험 결과를 얻게 되었다. 또 dCLK 단백질에서 아미노산 PAS-A, 145-263, PAS-B, 310-378 부위가 삭제가 되면 dCLK 의 발현량에도 영향을 미치는 것으로 확인이 되었다.Fig.1.Schematic diagram of internally deleted amino acid region on dCLK protein
bHLH, basic helix loop helix; A, PAS-A; B, PAS-B; Q-rich, glutamine rich; Poly Q, Poly glutamine; (1), DBD;(2). HLH;(3). 63-95;(4). PAS-A;(5), 145-263;(6). PAS-B;(7), 310-378;(8), 380-478;(9), 479-578;(10), 579-778;(11), 784-834;(12), 837-945
Fig.2.Interaction between PERIODwith CLOCK internal deletion mutants S2 cells were co-transected with 400 ng of each variant form of Clkplasmids as indicated on top (numbers as same as in Fig. 1) in combination with 600ng pAct-dper. At 36 h after transfection, cells were incubated in media
containing 500 uM CuSO4(final) to induce ectopic expression of target
proteins and harvested 24 h later. Extracts were prepared and either subjected to immunoprecipitation (IP) or analyzed directly (input). Immune complexes were recovered with anti-dPER (GP339) or anti-HA(12CA5)
Fig.3.Interaction between CYCLEwithCLOCK internal deletion mutants
S2 cells were co-transected with 500 ng of each variant form of Clk
plasmids as indicated on top (numbers as same as in Fig. 1) in combination with 500 ngpMT-cyc -3F. At 36 h after transfection, cells were incubated
in media containing 500 uM CuSO4 (final) to induce ectopic expression of
target proteins and harvested 24 h later. Extracts were prepared and either subjected to immunoprecipitation (IP) or analyzed directly (input). Immune complexes were recovered with anti-HA(12CA5) or anti-CYC (anti-FLAG)
B. S2 세포에서 dCLK(△657-707)은 dPER과의 결합을 하지
못한다.
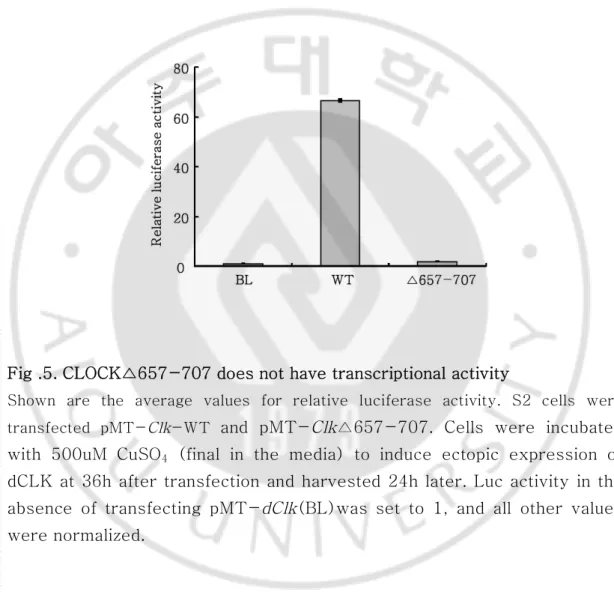
이전의 결과에서 dCLK(△579-778)이 dPER와의 결합을 하지 못하지만 또 다른 중요 단백질인CYC 와의 결합에는 문제가 없음을 immunoprecipitation(IP) assay 를 통해 확인하였다. 앞서 확인한 아미노산 579-778 부위를 더 좁혀 아미노산 657-707 부위가 삭제가 되면 dPER 단백 질이 dCLK에 결합하는데 중요한 최소 domain인 것을 앞서 실험한 방법과 같이 immunoprecipitat ion(IP) 를 통해서 알아 내었다. 이때도 앞선 결과와 마찬가 지로 CYC의 결합은 문제가 없었다.dCLK wild-type 과 dCLK(△657-707) 의 전사활성을 luciferase assay로 측정해 보았다. 그 결과 dCLK (△657-707) 는 전사활성이 wild type에 비해 사라져 버린 것을 확인할 수 있었다. 아미노산 657-707이 삭제되면 dPER 단백질이 결합하지 못하고 dCLK 자체의 전사활성 도 없어지게 된다.Fig .4. Amino acid 657-707 is necessary for PERIOD binding to CLOCK S2 cells were transected with 400 ngpMT-dClk-WT ,pMT-dClk(△657-707) in combination with 600ng pAct-dper.at 36 h after transfection, cells were
incubated in media containing 500 uM CuSO4(final) to induce ectopic
expression of target proteins and harvested 24 h later. Extracts were prepared and either subjected to immunoprecipitation (IP) or analyzed directly (input). Immune complexes were recovered with anti-dPER
(GP339) or anti-HA(12CA5) S2 cells were transected with pMT-dClk-WT or
Fig .5. CLOCK△657-707 does not have transcriptional activity
Shown are the average values for relative luciferase activity. S2 cells were transfected pMT-Clk-WT and pMT-Clk△657-707. Cells were incubated
with 500uM CuSO4 (final in the media) to induce ectopic expression of
dCLK at 36h after transfection and harvested 24h later. Luc activity in the absence of transfecting pMT-dClk(BL)was set to 1, and all other values were normalized.
Fig.6.Amino acid 657-707 is not necessary for CYCLE binding to CLOCK. S2 cells were transected with 500 ngpMT-dClk-WT ,pMT-dClk(△657-707) in
combination with 500ngpMT-cyc -3F.at 36 h after transfection, cells were
incubated in media containing 500 uM CuSO4(final) to induce ectopic
expression of target proteins and harvested 24 h later. Extracts were prepared and either subjected to immunoprecipitation (IP) or analyzed directly (input). Immune complexes were recovered with anti-HA(12CA5)
or anti-CYC (anti-FLAG).S2 cells were transected with pMT-dClk-WT or
C. dCLK(△657-707) 을 발현하는 형질전환 초파리는 정상적
인 생체리듬을 가지지 못한다.
dCLK 의 아미노산 657-707 의 in vivo 에서의 역할을 탐색하기 위하여 아미 노산 657-707 부위가 제거된 dCLK 을 발현하는 초파리를 제작하였다..dCLK (△657-707) 을 발현하는 독립적인 2개의 형질전환 초파리 라인 (3M, 4M)을 확립하였고 CLKOUT background 의 초파리와 교배 후 이를 dClk(△657-707) 초파리라 명명하였다. 정상적인 dClk을 발현하는 초파리 역시 CLKOUT background 의 초파리와 교배 후 이를 dClk(WT)이라 명명하고 대조군으로 사용하였다. 생체리듬은 DAM(Drosophila Activity Monitoring)system을 이용하
여 분석하였다. (Table. 1) 그 결과 dClk(WT) 초파리는 homozygote 형태로
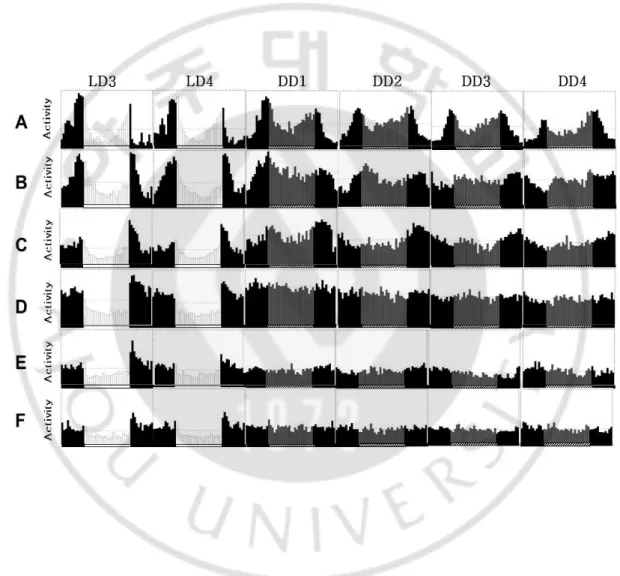
존재 할 경우 24.4시간의 주기를 가지고 heterozygote의 상태가 되면 27.3 시 간의 주기를 가지게 된다.이처럼 transgene 을 넣어준 경우에는 dose effect 가 존재하기 때문에 WT gene이 1copy (heterozygote) 가 들어가면 2 copy (homozygote) 가 들어갔을 때에 비해 rhythmicity가 확연히 떨어지는 것을 확 인할 수 있다.dClk(△657-707) 의 경우에는 WT 과는 확연히 다른 결과를 보 여 주었다. Homozygote 와 heterozygote의 형태에 상관없이 주기를 가지는 초 파리가 없다는 실험 결과가 나왔다. 같은 mutant이지만 line 에 따라 다양성을 나타내는데 이는 같은 유전자를 가지고 발현을 하더라도 모두 같은 행동 양상을 나타내는 것이 아니라는 것을 알려주는데 3M 과 4M mutant line 같은 경우
activity 양의 차이는 있으나 이들의 행동 양상은 비슷하게 나타나는 것을 볼수 있었다. 이는 dClk(△657-707) 을 발현하는 초파리의 경우 주기를 형성하는데 중요한 역할을 하는dPER 단백질이 dCLK 단백질에 결합하지 못해서 앞선 in vitro 실험에서도 보았듯이 dCLK의 전사활성이 사라지는 것에 대한 결과인 것 으로 말해준다. 그렇다면 생체리듬 주기가 사라져 버린 dClk(△657-707) 형 질전환 초파리의 생체 내에서 dPER 는 어떠한 변화가 있는지 조사하기 위해 dClk(△657-707) 형질전환 초파리를 12:12 LD cycle 에 entrainment 시킨 후 4 시간 간격으로 초파리를 수확하여, 머리로부터 단백질 액을 얻어 western blot 을 수행하였다. 정상의 dCLK단백질은 시간에 따라 그 인산화 정도가 순환 한다. 늦은 저녁과 이른 아침에는 과도하게 인산화된 (hyperphosphorylation) isoform 들이 많이 존재하고 하루의 중간에는 비교적 적게 인산화된 (hypophosphorylation) isoform들이 많이 존재한다 (Fig 8). 반면에 dCLK(△ 657-707 단백질은 하룻 동안 인산화 isoform의 변화가 없이 일정한 것을 관찰 할 수 있었고, 더욱이 그 양이 정상의 dCLK 단백질에 비해 매우 높은 것을 관
찰하였다PER 와 TIM 단백질은 dClk△657-707;ClkOUT 초파리에서 그 양은
dClk;ClkOUT
초파리보다 낮지만 시간에 따라 변화하는 것을 관찰하였다. 그리고 24 시간 동안 constant - dark (DD) 상태일 때는 WT은 LD와 비슷하게 rhythmic한 양상을 나타내는데 비해 mutant의 TIM과 PER는daily activity patterns 결과와 동일하게 peak이 뒤로 점차 밀리는 것을 확인할 수 있었
다.dCLK△657-707 단백질의 전사활성을 측정하기 위해
dClk△657-707;ClkOUT 에서 하룻 동안 tim mRNA 양을 측정하였다. CLK의 mRNA 경 우 LD와 DD 에 mutant는 비슷한 graph를 나타내는데 dCLK의 level은 mutant의 경우 더 높은 것을 확인할 수 있었다. 하지만 dCLK의 repressor인
TIM의 mRNA의 변화량은ZT에서 WT에 비해 mRNA의 양이 떨어져 있음을 확 인할 수 있었다. 이것은 S2 cell 에서와 같이 dCLK△657-707 단백질이 전사 활성을 완전히 잃은 것은 아니고 어느 정도는 가지고 있는 것을 의미한다.DD에 서는 mRNA의 peak이 뒤쪽으로 밀리는 것을 볼 수 있었다. 이 결과는 앞선 행 동분석 결과 그리고 molecular rhythm의 변화에 일치하는 결과를 보여주고 있 다.
a Measure of the strength or amplitude of the rhythm.
b
Percentage of flies with activity rhythms having a power value of 10 and a width value of 2.
cTotal number of flies that survived until the end of the testing period.
d AR = arrhythmic Period ± SEM(h) Power a Rhythmicity (%)b Number c ClkOUT AR AR AR 11 w;Clk-wt,1A;ClkOUT 24.4±0.11 77.6 100 11 w;Clk-wt,1A/CyO;ClkOUT 27.3±1.79 59.7 46.2 15 w;Clk△657-707-3M;ClkOUT 26±0.43 81.2 67.9 28 w;Clk△657-707-3M/CyO;ClkOUT 24.3±0.17 47.8 15 20 w;Clk△657-707-4M;ClkOUT AR AR AR 10 w;Clk△657-707-4M/CyO;ClkOUT AR AR AR 8
Fig. 7.Daily activity patterns of Clk-WT; ClkOUT and Clk△657-707; ClkOUT
Flygenotypes are A(w;Clk-WT,1A;Clkout)
,B(w;Clk-WT,1A/CyO;Clkout) ,C(w;Clk△657-707-3M;Clkout)
,D(w;Clk△657-707-3M/CyO;Clkout) ,E(w;Clk△657-707-4M;Clkout)
Fig.8. Amino acid 657-707 is necessary for PERIOD binding to CLOCK atin vivo
Adult flies of each genotype (indicated above the panel; WT,
w;Clk-WT;ClkOUT; △657-707, w;Clk△657-707;ClkOUT) were collected at the
indicated ZT. Fly headextracts were preparedand either subjected to immunoprecipitation (IP) with anti-V5 antibody or analyzed directly (input). Immune complexes were recovered with dPER (Rb1) or anti-dCLK(Rb-V5).
Fig.9.Molecular rhythms of Clk-WT; ClkOUT andClk△657-707;ClkOUT
during Light/Dark Cycle
Adult flies of each genotype (indicated above the panel; WT, w
;Clk△657-707) were collected at the indicated ZT . Head extracts were prepared and analyzed by immunoblotting. Anti-CLK(V5) or TIM(TR-3) and anti-PER(Rb1) antibodies were used to visualize dPER or TIM, respectively
Fig.10.Molecular rhythmsofClk-WT; ClkOUT andClk△657-707;ClkOUT
during Light/Dark Cycle
Adult flies of each genotype (indicated above the panel) were collected at the indicated CT. Head extracts were prepared and analyzed by immunoblotting. Anti-CLK(V5) or anti-TIM(TR-3) and anti-PER(Rb1) antibodies were used to visualize dPER or TIM, respectively
Fig.11.. Daily levels of tim and clk mRNAs Clk-WT; ClkOUT
andClk△657-707;ClkOUT flies
cycles in dclkor timRNA levels. Adult flies of the indicated genotypes (as
shown in Fig.8.9 were collected at the indicated ZT and CT. RNA was extracted from fly heads and quantitative real-time RT-PCR used to
measure the relative levels of total dclk or tim mRNA. RNA levels from
control flies (A,C =clk-wt )(B,D = clk△657-707) at ZT4 were set to 1 and all other values were normalized. Shown are the average values from three independent experiments. Error bars denote S.E.M.
IV. 고 찰
모든 생물은 생체시계를 가지며 이것을 이용하여 하루 중 생체리듬을 조절한다. 그 중에서도 분자생물학적 유전학적 연구가치가 뛰어나며 인간과 상동성이 높은 초파리 동물모델 생체시계의 transcriptional-translational feedback loop 에서 중요한 역할을 수행하는 dCLK 의 분자생물학적• 생화학 적 역할을 규명하는 것은 생체시계 연구 분야에서 매우 중요한 연구이다. dCLK 의 서열 중 dPER binding domain (아미노산 657-707) 의 역할을 찾고자 하였다. 그 결과 우리는 아미노산 579-778 중에서도 중요한 역할을 하는부위를 아미노산 657-707 로 줄일 수 있었고 이 부위가 제거 되었을 경우 전사 억제제 역할을 하는 dPER 의 결합이 되지 않는 것을 확인할 수 있었으며, dCLK 의 전사활성마저 잃어 버리는 것을 발견하였다. 한 편 in vitro 뿐만 아니라 in vivo 결과에서도 중요한 의미를 가진다. dCLK-WT gene 을 발현하는 형질전환 초파리는 약 24.4 시간의 주기를 가지며 rhythmic 한 반면 dCLK(△657-707) 을 발현하는 형질전환 초파리의 독립적인 라인에서 모두 주기가 없어졌고 정상적인 생체리듬을 가지지 못하는 것을 알 수 있었다. 초파리 세포배양에서 확인한 바에 의하면, dCLK (△657-707) 은 dPER 와 결합할 수 있는 능력을 잃어버려 dCLK 의 전사 활성을 억제하지 못하였다. dCLK (△657-707) 은 일부분만이 삭제가 되었음에도 전사 억제제 역할을 하는 dPER 가 결합할 수 있는 능력도 잃어버릴 뿐만 아니라 dCLK 의 자체적인 전사활성 능력마저도 잃어 버린다는 사실은 매우 흥미롭다. 이러한 dClk (△657-707)을 발현시키는 형질전환초파리는 주기도 나타나지 않을 뿐 아니라 arrhythmic 한 현상을 나타낸다. in vitro 에서는 dCLK (△657-707)이 전사활성을 잃어 버리지만 in vivo 에서는 dPER 와 TIM 이 시간에 따라 thythmic 하게 생성되는 것은 매우
흥미롭다. 이는 dCLK (△657-707) 단백질의 분자적 생화학적 결함이 in vivo 에서는 다를 수 있다는 것을 시사하는 것으로 이에 대한 연구는 앞으로 더욱 진행되어야 할 것이다. 이러한 dCLK 의 아미노산 657-707 의 역할을 현재 기술한 논문에서 전사활성, 단백질의 변화, 그리고 행동학적 측면에서 관찰 하였는데 앞으로의 또 다른 접근의 연구가 필요할 것이라고 생각된다. 그래서 우리는 첫 째, 형질전환 초파리의 생체 내 gene 들의 RNA level 을 확인할 것이다. 둘 째,
dCLK 과 dPER 가 생채 내에서도 결합할 수 있는지
Immunoprecipitation(IP)을 통해 조사 할 것이다. in vitro 의 결과와
동일하다면 생체 내에서도 결합하지 못할 것이다. 현재 진행중에 있는 이러한 연구들이 결실을 맺게 된다면 생체시계 연구분야의 생화학적• 분자생물학적 기전을 이해하는데 매우 중요한 역할을 할 것이라 생각된다.
V. 결 론
모든 생명체는 생체시계를 가지고 있으며, 생체시계의 핵심기전은
“interlocked transcriptional-translational feedback loop” 으로써 두 개의 loop 이 서로 맞물려 생체시계의 핵심 유전자들의 발현이 하루 24 시간을 주기로 진동할 수 있도록 한다. 앞서 발표된 논문에서 dPER 의 아미노산 926-977 의 역할이 dCLK 이 결합하는 부위로 밝혀졌고 (Sun 등 2010), 우리는 dCLK 의 아미노산 657-707 의 역할을 밝혀내기 위해 연구를 진행하였다. 먼저 dCLK 에서 아미노산 657-707 이 제거된 plasmid 를 만들었고, 이것을 이용하여 S2 세포에서 immunoprecipitation(IP)을 통해 확인 결과 dPER 가 결합하지 못하였다. 뿐만 아니라 dPER 의 전사활성 억제가 일어나지 않음에도 dCLK 에서 아미노산 657-707 이 제거된 plasmid 는 전사활성 자체가 없어지는 것을 관찰할 수 있었다. 이 현상이 dCLK 과 이형 이합체를 형성하는 CYC 가 결합하지 못해서 일어나는 현상이 아니라는 것을 Immunoprecipitation (IP)를 수행함으로써 알 수 있었다. 초파리 수준에서 dCLK 의 아미노산 657-707 의 역할을 연구하기 위해 아미노산 657-707 이 제거된 dCLK 을 발현하는 초파리를 제작하였다. 제작한 형질전환 초파리를 이용하여 하루 중 생체리듬을 측정한 결과 정상적인 dCLK 을 발현하는 초파리는 정상적인 rhythm 을 나타내는데 비해 dClk(△657-707)을 발현하는 초파리는 두 개의 독립적인 라인에서 activity 에서는 차이가 보이긴 했으나 모두 arrhythmic 해지는 것을 알 수
있었다.그리고 dClk(△657-707)을 발현하는 초파리의 molecular rhythm
에서는 in vitro 의 결과에서 예상치 못했던 PER 와 TIM 단백질의 생성이
WT 에 비해 level 이 낮지만 rhythmic 하게 만들어 진다는 사실을 알게 되었다. 이 것을 좀더 자세히 알아보기 위해 12:12 LD 상태에 둔 초파리와 하루 동안 어둠 상태에 둔 초파리의 mRNA 의 변화량을 확인한 결과 dClk 의 경우 WT 에 비해 PER 에 의한 저해를 받지 않기 때문에 level 이 높은 것을 확인할 수 있었지만 시간에 따른 변화량은 WT 과 비슷한 주기를 가지고 있었다. 하지만 tim 의 경우에는 12:12 LD 상태에 둔 초파리의 하루 동안의 변화량은 WT 에 비해 level 이 낮으며 거의 주기를 가지지 않는 것으로 확인이 되었다. 이 결과는 우리가 만든 dClk(△657-707)을 발현하는 초파리가 전사활성을 잃어 버리는 것에 의해 예측 가능한 결과였다.하루 동안에 어둠 상태에 놓였던 초파리의 dClk 의 mRNA 변화량은 WT 과 크게
차이가 없으며 level 이 더 높은 것을 볼수 있는데 tim 의 mRNA
변화량은 WT 에 비해 전사활성이 더욱 떨어진 것을 확인할 수 있었다.tim mRNA 가 시간에 따른 주기를 가지지 못하면서 늦은 저녁인 CT 20 에서 peak 을 나타내는데 mRNA 에서도 시간에 따른 변화가 molecular level 의 변화와 일치하는 현상을 확인 할 수 있었다.
참고문헌
1.Panda S, Antoch MP, Miller BH, Su AI, Schook AB, Straume M,
Schultz
PG,
Kay
SA,
Takahashi
JS,
Hogenesch
JB.
2002.Coordinated transcription of key pathways in the mouse by
the circadian clock. Cell. 109(3):307-20.
2.Bunney WE, Bunney BG. 2000. Molecular clock genes in man and
lower animals: possible implications for circadian abnormalities in
depression. Neuropsychopharmacology. 22(4):335-345.
3.Cardinali DP. 2000. The human body circadian: How the biologic
clock influences sleep and emotion. NeuroEndocrinolLett.
21(1):9-15.
4.McClung CA. 2007. Role for the Clock gene in bipolar disorder.Cold
Spring HarbSymp Quant Biol. 72:637-44.
5.Rudic RD, McNamara P, Curtis AM, Boston RC, Panda S, Hogenesch
JB, Fitzgerald GA. 2004. BMAL1 and CLOCK, two essential
components of the circadian clock, are involved in glucose
homeostasis.PLoS Biol. 2(11):e377
suppressor. Nat Rev Cancer. 3(5):350-61.
7.Bae K, Lee C, Sidote D, Chuang KY, Edery I. 1998. Circadian
regulation of a Drosophila homolog of the mammalian Clock gene:
PER and TIM function as positive regulators. Mol Cell Biol.
18(10):6142-51.
8. Cyran SA, Buchsbaum AM, Reddy KL, Lin MC, GlossopNR,Hardin
PE, Young MW, Storti RV, Blau J. 2003. vrille, Pdp1, and dClock
form a second feedback loop in the Drosophila circadian clock.
Cell.112(3):329-41.
9.Myers MP, Wager-Smith K, Rothenfluh-Hilfiker A, Young MW.
1996. Light-induced degradation of TIMELESS and entrainment
of the Drosophila circadian clock.Science. 271(5256):1736-40.
10. Cyran SA, Yiannoulos G, Buchsbaum AM, Saez L, Young MW,
Blau J. 2005. The double-time protein kinase regulates the
subcellular localization of the Drosophila clock protein period. J
Neurosci. 25(22):5430-7.
11. Chang DC, Reppert SM. 2003. A novel C-terminal domain of
drosophila
PERIOD
inhibits
dCLOCK:CYCLE-mediated
transcription. curr.biol .758-62
DOUBLETIME kinase binding domain on the Drosophila PERIOD
protein is essential for its hyperphosphorylation, transcriptional
repression, and circadian clock function. Mol Cell Biol.
27(13):5014-28.
13. Sun WC, Jeong EH, Jeong HJ, Ko HW, Edery I, Kim EY. 2010.
Two distinct modes of PERIOD recruitment onto dCLOCK reveal
a novel role for TIMELESS in circadian transcription. J Neurosci.
30(43):14458-69.
14. Darlington TK, Wager-Smith K, Ceriani MF, Staknis D, Gekakis
N, Steeves TD, Weitz CJ, Takahashi JS, Kay SA. 1998. Closing
the circadian loop: CLOCK-induced transcription of its own
inhibitors per and tim.Science. 280(5369):1599-603.
15. Hardin PE. 2005. The circadian timekeeping system of
Drosophila.Curr Biol. 15(17):R714-22.
16. Cyran SA, Yiannoulos G, Buchsbaum AM, Saez L, Young MW,
Blau J. 2005. The double-time protein kinase regulates the
subcellular localization of the Drosophila clock protein period. J
Neurosci. 25(22):5430-7.
17. Glossop NR, Houl JH, Zheng H, Ng FS, Dudek SM, Hardin PE.
2003.VRILLE feeds back to control circadian transcription of
Clock in the Drosophila circadian oscillator. Neuron.
37(2):249-61.
18. Markstein M, Zinzen R, Markstein P, Yee KP, Erives A,
Stathopoulos A, Levine M. 2008. A regulatory code for
neurogenic
gene
expression
in
the
Drosophila
embryo.Development. 131(10):2387-94.
-Abstract-
Role of PERIOD binding domain for CLOCK in
Drosophila
circadian clock
Doo-Hyun Kang
Department of Medical Sciences (Neuroscience) The Graduate School, Ajou University
(Supervised by Associate Professor Eun-Young Kim)
Negative transcriptional feedback loops are a core feature of eukaryotic circadian clocks and are based on rhythmic interactions between clock-specific repressors and transcription factors. In Drosophila, the repression of CLOCK (CLK)-CYCLE (CYC) transcriptional activity by PERIOD (PER) is critical for driving circadian gene expression. To better understand how PER inhibits CLK-CYC transcriptional activity, we sought to identify PER binding domain of CLK. To this end, we made several versions of mutant CLK which is internally deleted with 100~ 200 amino
acids.We evaluated the interaction between each CLK internal deletion mutants and PER via immunoprecipitation. CLK internally deleted with aa 657-707, which is similar to 19 region of mammalian CLK, could not interact well with PER in S2 cells. To investigate the role for amino acid
657-707 in vivo,we made transgenic flies expressing Clk△657-707 while
clk-WT rescued the arrhythmicity of clkOUT
flies, CLK△657-707 could not restore arrhythmic locomotorbehavior.This result suggest that as in the case of S2 tissue culture, amono acid 657-707 might play critical roles for maintaining circadian clock system. Further study will be necessary to
investigate the role of amino acid 657-707 in vivo using
Clk△657-707 ;ClkOUT flies
Keywords: Drosophila, Circadian clock, dPERIOD, dCLOCK, per binding