저작자표시-비영리-변경금지 2.0 대한민국 이용자는 아래의 조건을 따르는 경우에 한하여 자유롭게 l 이 저작물을 복제, 배포, 전송, 전시, 공연 및 방송할 수 있습니다. 다음과 같은 조건을 따라야 합니다: l 귀하는, 이 저작물의 재이용이나 배포의 경우, 이 저작물에 적용된 이용허락조건 을 명확하게 나타내어야 합니다. l 저작권자로부터 별도의 허가를 받으면 이러한 조건들은 적용되지 않습니다. 저작권법에 따른 이용자의 권리는 위의 내용에 의하여 영향을 받지 않습니다. 이것은 이용허락규약(Legal Code)을 이해하기 쉽게 요약한 것입니다. Disclaimer 저작자표시. 귀하는 원저작자를 표시하여야 합니다. 비영리. 귀하는 이 저작물을 영리 목적으로 이용할 수 없습니다. 변경금지. 귀하는 이 저작물을 개작, 변형 또는 가공할 수 없습니다.
Master’s Thesis in Molecular Medicine
Effects of Loganin on osteoblast and
osteoclast differentiation
Ajou University Graduate School
Major in Molecular Medicine
Department of Biomedical Sciences
Effects of Loganin on osteoblast and
osteoclast differentiation
Seon-Yong Jeong, Advisor
I submit this thesis as the
Master’s thesis in Molecular Medicine.
February 2018
Ajou University Graduate School
Major in Molecular Medicine
Department of Biomedical Sciences
i
- Abstract -
Effects of Loganin on osteoblast and osteoclast differentiation
The maintenance of bone mass involves both bone formation and bone resorption, and imbalances between these processes can result in bone loss and, eventually, osteoporosis. We demonstrated that an ethanol extract of Cornus
officinalis increased the differentiation of osteoblasts from pre-osteoblast cells and
decreased the differentiation of osteoclasts from primary-cultured monocyte cells. The goals of the present study were to isolate the bioactive compound(s) responsible for the anti-osteoporotic effects of the Cornus officinalis extract and to
investigate the anti-osteoporotic effects of the isolated compound(s) in vitro and in
vivo. A single compound, Loganin, was isolated as the putative candidate bioactive component. Furthermore, Loganin significantly increased the activity of alkaline phosphatase (ALP) and expression of bone remodeling markers (e.g., Alp and
Runx2 in) pre-osteoblast MC3T3-E1 cells and increased the formation of
mineralized nodules in MC3T3-E1 cells. On the other hand, Loganin reduced the activity of tartrate-resistant acid phosphatase (TRAP) in the osteoclasts of primary-cultured monocytes from mouse bone marrow. Similarly, Loganin also increased
ii
the ALP activity and decreased the TRAP activity of co-cultured osteoblasts and osteoclasts. In vivo, Loganin improved the bone mineral density, serum OPG/RANKL ratio, and micro CT and whole-body bone scan results in Loganin-treated ovariectomized (OVX) mice.
Together, these results demonstrate that Loganin effectively enhances bone formation and reduces bone resorption during bone remodeling and suggests that Loganin could be used to treat osteoporosis.
_________________________________________________________________________
Keywords: natural extract, Cornus officinalis, Loganin, osteoporosis, osteoblast,
iii
TABLE OF CONTENTS
ABSTRACT ··· i
TABLE OF CONTENTS ··· iii
LIST OF FIGURES ··· v
LIST OF TABLES ··· vii
I. INTRODUCTION ··· 1
II. MATERIALS AND METHODS ··· 4
A. Fractionation, isolation, and identification of bioactive component(s)· 4 B. Reagents and cell culture materials ··· 7
C. Water-soluble tetrazolium salt (WST) assay in osteoblast cells ··· 8
D. Alkaline phosphatase (ALP) activity assay and staining in osteoblast cells ··· 8
E. Mineralized nodule formation in osteoblast cells··· 9
F. Quantitative reverse-transcription PCR (qRT-PCR) ··· 10
G. Osteoclastogenesis of primary cultured monocytes ··· 11
H. Tartrate-resistant acid phosphatase (TRAP) activity assay and staining in monocytes ··· 12
I. Co-culture of MC3T3-E1 cells and primary monocytes ··· 12
J. Ovariectomized model ··· 13
iv
L. Microcomputed tomography (Micro CT) and SPECT scan ··· 14
M. Blood sampling and serum OPG and RANKL measurement ··· 15
N. Statistical analysis ··· 16
III. RESULTS ··· 17
A. Cellular differentiation of osteoblastic ME3T3-E1 cells ··· 17
B. Bioactive component of the Cornus officinalis extract ··· 19
C. Effect of Loganin on osteoblast differentiation and mineralized nodule formation ··· 21
D. Effect of Loganin on the expression of bone remodeling markers ··· 23
E. Effect of Loganin on osteoclast differentiation ··· 25
F. Effect of Loganin on osteoblast and osteoclast differentiation ··· 27
G. Effect of Loganin on OVX-induced bone loss in an osteoporotic mouse model ··· 29
H. Effect of Loganin on serum OPG and RANKL levels ··· 35
IV. DISCUSSION ··· 37
V. CONCLUSION ··· 41
REFERENCES ··· 42
v
LIST OF FIGURES
Figure 1. Bone remodeling ··· 3
Figure 2. Isolation of Loganin through the fractionation of a 70% ethanol extract of Cornus officinalis ··· 5
Figure 3. Nuclear magnetic resonance spectra of the bioactive (D2) fraction ··· 6
Figure 4. Chemical structure of Loganin ··· 20
Figure 5. Effect of Loganin on the alkaline phosphatase (ALP) activity, ALP staining, and Alizarin Red S staining of MC3T3-E1 cells ··· 22
Figure 6. Effect of Loganin on the expression of bone remodeling markers ·· 24
Figure 7. Effects of Loganin on osteoclast differentiation in primary-cultured monocytes ··· 26
vi
Figure 8. Effect of Loganin on the alkaline phosphatase (ALP) and tartrate-resistant acid phosphatase (TRAP) activity of co-cultured cells MC3T3-E1 and monocytes ··· 28
Figure 9. Effects of Loganin on the bone mineral density (BMD) of ovariectomized (OVX) mice ··· 31
Figure 10. Anti-osteoporosis effect of Loganin on the bones of ovariectomized (OVX) mice ··· 32
Figure 11. Anti-osteoporosis effect of Loganin on the femurs of ovariectomized (OVX) mice ··· 33
Figure 12. Anti-osteoporosis effect of Loganin in ovariectomized (OVX) mice ··· 34
Figure 13. Anti-osteoporosis effect of Loganin on serum levels of OPG and RANKL in ovariectomized (OVX) mice ··· 36
vii
LIST OF TABLES
Table 1. Alkaline phosphatase (ALP) activity of MC3T3-E1 cells treated with extracts from 66 plant species native to Korea ··· 18
- 1 -
I. INTRODUCTION
Bone is one of the hardest tissues in the human skeleton and is maintained through the processes of bone formation and bone resorption (Hadjidakis and Androulakis, 2006), during which osteoclasts and osteoblasts continuously remove old and damaged bone and regenerate the removed tissue, respectively (Bernhardt et al., 2017). Osteoblasts differentiate from mesenchymal stem cells, whereas osteoclast cells differentiate from the monocyte/macrophage lineage, the cells differentiated according to Receptor activator of nuclear factor kappa-B ligand (RANKL) (Weilbaecher et al., 2011) (Figure 1).
Imbalances between bone resorption and formation can result in inappropriate bone remodeling and, eventually, osteoporosis and osteopenia (Zheng et al., 2014). Osteoporosis characterized by the systemic reduction of bone mass and microarchitecture (Moreira et al., 2014). Osteoporosis patients in elderly or postmenopausal women tends to aggressive bone resorption compared to bone formation, leading to an enhanced risk of bone fragility and susceptibility to fractures (Bernhardt et al., 2017). Currently used osteoporosis treatment agents include bisphosphonates, which inhibit bone resorption, and vitamin D and calcium supplements (Tella and Gallagher, 2014). However, these agents are inappropriate for long-term use, owing to their side effects (Ishtiaq et al., 2015). As a result, many
- 2 -
studies have aimed to identify therapeutic agents that can be used for long periods of time with fewer side effects (Park et al., 2016; Liu et al., 2017), and the alternative use of natural products as therapeutic agents has attracted increasing interest, especially because they typically have fewer side effects.
A variety of plant-derived products have been used in traditional medicine to treat diseases since ancient times. The present study assessed the effectiveness of 66 plant extracts in treating osteoporosis and selected the extract of Cornus
officinalis as a putative osteoporosis remedy. Studies have reported that Cornus
officinalis extract imparts a variety of beneficial effects, such as the inhibition of
diabetic diseases, inflammation, oxidative stress, and osteoclast differentiation (Ma et al., 2014; Huang et al., 2017)), that the Cornus officinalis extract increases the differentiation of osteoblasts from pre-osteoblast MC3T3-E1 cells and decreases the differentiation of osteoclasts from pre-osteoclast monocyte cells. However, despite the positive effect of Cornus officinalis extracts on osteoporosis, the specific bioactive compounds responsible for the extract’s activity have yet to be reported. Therefore, the goals of the present study were to isolate the bioactive compound(s) responsible for the anti-osteoporotic effects of Cornus officinalis and to investigate
- 3 -
Figure 1. Bone remodeling (Krane, 2005). Hematopoietic progenitor-derived
osteoclasts resorb bone (releasing growth factors), and mesenchymal progenitor-derived osteoblasts replace the gaps with new bone. Pre-osteoblast express receptor activator of the receptor activator of nuclear factor kappa-B ligand (RANKL), a molecule that binds to the receptor activator of nuclear factor kappa-B on preosteoclast cells and stimulates osteoclast differentiation (Krane, 2005).
- 4 -
II. MATERIALS AND METHODS
A. Fractionation, isolation, and identification of bioactive component(s)
A 70% ethanol extract of Cornus officinalis was evaporated, suspended in H2O, and then partitioned successively using hexane (1.6 g), dichloromethane (1.9
g), ethyl acetate (5.2 g), and butanol (40 g; Figure 2). The effect of each fraction (A– D, respectively) on osteoblast differentiation was evaluated. Fraction D was then subject to chromatography using a Diaion HP-20 gel column and a gradient H2O-methanol solvent system (0, 20, 40, 60, 80, and 100% MeOH). Of the resulting subfractions (D1-D6), subfraction D2 was subject to preparative HPLC, eluting with MeOH-H2O to give compound. The structure of the compound in the bioactive
fraction was determined using proton nuclear magnetic resonance (1H-NMR; Figure
3. A), carbon-13 nuclear magnetic resonance (13C-NMR; Figure 3. B), and mass
- 5 -
Figure 2. Isolation of Loganin through the fractionation of a 70% ethanol extract of Cornus officinalis. Abbreviations: Fr., fraction; CC, column
- 6 -
Figure 3. Nuclear magnetic resonance spectra of the bioactive (D2) fraction.
Proton nuclear magnetic resonance (1H-NMR) (A), carbon-13 nuclear magnetic resonance (13C-NMR) (B), and mass spectrum (C) analyses of the D2 fraction.
- 7 -
B. Reagents and cell culture materials
For the osteoblastic differentiation analysis, pre-osteoblast cell line, MC3T3-E1 (RIKEN Cell Bank, Tsukuba, Japan), was used in this study (Li et al., 2010; Addison et al., 2015). MC3T3-E1 cells were cultured in α-modified minimal essential medium (α-MEM) that was supplemented with 10% FBS, penicillin (100 U/ml), and streptomycin (100 μg/ml). After allowing 24 h for cell adhesion, osteoblast differentiation was induced by the addition of osteogenic medium that contained ascorbic acid (50 μg/ml) and β-glycerophosphate (10 mM). All the cells were cultured in a humidified atmosphere at 37 °C and 5% CO2, and the medium of
the cultures was changed every 3 d.
Mouse-derived monocytes were cultured in α-MEM culture medium that was supplemented with 10% FBS and macrophage colony-stimulating factor (M-CSF; 100 ng/ml). After allowing 24 h for cell adhesion, osteoclast differentiation was induced by the addition of osteogenic medium that contained RANKL (100 ng/ml). All the cells were cultured in a humidified atmosphere at 37 °C and 5% CO2, and the
- 8 -
C. Water-soluble tetrazolium salt (WST) assay in osteoblast cells
Pre-osteoblast MC3T3-E1 cells were incubated in a 96-well plate (5×103
cells/well) for 24 h, treated with different concentrations of the Loganin (D2) fraction (1, 5, or 10 μg/ml), and incubated for an additional 48 h, following the methods of other similar studies (e.g., Zeng et al., 2015; Guo et al., 2016). To determine cell viability, WST solution (20 μl, 5 mg/ml in phosphate-buffered saline) was added to each well, and after another 2 h of incubation, the absorbance of each well was measured at 450 nm and 595 nm using a microplate reader (BIO-RAD; Hercules, CA, USA).
D. Alkaline phosphatase (ALP) activity assay and staining in osteoblast cells
Pre-osteoblast MC3T3-E1 cells were incubated for 24 h in a 48-well plate (2×104 cells/well) for adhesion to occur. Osteoblast differentiation was then induced
by the addition of media that contained ascorbic acid (50 μg/ml) and β-glycerophosphate (10 mM), and after different concentrations of Loganin (1, 5, or 10 μg/ml) were added, the cells were incubated for another 3 d. To measure ALP activity, the cells were homogenized with lysis buffer [0.5 M Tris-HCl (pH 9.0), 0.9% NaCl, 1% Triton X-100, 200 mM EDTA] and reacted with alkaline phosphatase yellow (pNPP) solution (Sigma-Aldrich) at 37 °C. Then, after 24 h, the absorbance
- 9 -
of each well was measured at 405 nm using a microplate reader (BIO-RAD) (Bhargavan et al., 2009). For staining, the medium was carefully removed from the wells, and the cells washed were with PBS, fixed by incubation in 4% paraformaldehyde for 15 min at room temperature, rinsed with triple-distilled water (3DW), and stained using BCIP/NBT (Sigma-Aldrich) solution. Positive ALP staining was determined using a light microscope.
E. Mineralized nodule formation in osteoblast cells
Pre-osteoblast MC3T3-E1 cells were incubated for 24 h in a 48-well plate (2×104 cells/well) in order for adhesion to occur. Osteoblasts were incubated in
differentiation media that contained ascorbic acid (50 μg/ml) and β-glycerophosphate (10 mM) and treated with Loganin (1 μg/ml) for 21 d, with the medium being changed every 2 or 3 d. Matrix mineralization and nodule formation were determined using Alizarin Red S (Sigma-Aldrich). For staining, the medium was carefully removed from the wells, and the cells were washed with PBS, fixed by incubation in 4% paraformaldehyde for 15 min at room temperature, rinsed with 3DW, and stained in 40 mM Alizarin Red S. Positive Alizarin Red S staining was determined using a light microscope.
- 10 -
F. Quantitative reverse-transcription PCR (qRT-PCR)
RNA was extracted from cultured cells using TRIzol reagent (Invitrogen; Carlsbad, CA, USA), and the quality of the isolated RNA was measured using the ratio of absorbances at 260 and 280 nm. The RNA was subsequently reverse transcribed using a RevertAid First Strand cDNA Synthesis Kit (Thermo Fisher Scientific; Waltham, MA, USA), with oligo(dT)18 as a random primer, and real-time
reverse transcription polymerase chain reaction (qRT-PCR) was performed using the ABI Prism 7000 Sequence Detection System (Applied Biosystems; Foster City, CA, USA), a SYBR Green qPCR Kit (TaKaRa; Shiga, Japan), and 10-μl reaction volumes that contained 100 ng cDNA.
The following primers were used to amplify osteoblast markers: 5′-TCC CAC GTT TTC ACA TTC GG-3′ and 5′-GGC CAT CCT ATA TGG TAA CGG G-3′ for mouse Alp (GenBank: NM_007431.3; 117 bp), 5′-TAA AGT GAC AGT GGA CGG TCC C-3′ and 5′-CCT CAG TGA TTT AGG GCG CA-3′ for mouse Runx2 (GenBank: NM_009820.5; 104bp), and 5′-TGA CCA CAG TCC ATG CCA TC-3′ and 5′-GAC GGA CAC ATT GGG GGT AG-3′ for mouse Gapdh (GenBank: NM_001289726.1; 203 bp). The qRT-PCR conditions were as follows: denaturation at 95 °C for 5 min; 40 cycles of amplification at 95 °C for 5 sec, 60 °C for 30 sec, and 72 °C for 30 sec; and a terminal melting step of 72 to 95 °C for 2 min (5 sec per degree). By normalizing to Gapdh, the relative quantification of gene expression was
- 11 -
performed using the comparative threshold (Ct) method previously described (Livak and Schmittgen, 2001)
G. Osteoclastogenesis of primary cultured monocytes
To prepare primary-cultured monocytes, the bone marrow of femoral bones from 7-week-old BALB/c mice was removed by flushing with a fine-bore syringe and incubated in α-MEM medium that contained 100 ng/ml of macrophage colony-stimulating factor (M-CSF; PeproTech, Rocky Hill, NJ, USA) for 3 d (Kikuta & Ishii, 2013; Kim & Kim, 2016). The identity of the isolated monocytes was validated by immunophenotypic analysis, using CD11b antibody (BioLegend; San Diego, CA, USA), the FACS Aria Ⅲ Cell Sorter (BD Biosciences; San Jose, CA, USA), and FACS Diva (BD Biosciences). The animal research procedures were approved by the Animal Care and Use Committee of the Ajou University School of Medicine (IACUC No. 2014-0066), and all experiments were conducted in accordance with the institutional guidelines established by the committee. All efforts were made to minimize animal suffering and the number of mice used.
The monocytes were incubated in α-MEM medium that contained 10% FBS and M-CSF (100 ng/ml) for 3 d, and thought sub-culture, the monocyte cells were incubated in a 96-well plate (2×104 cells/well) into α-MEM medium with 10% FBS
- 12 -
and M-CSF (100 ng/ml), receptor activator of nuclear factor kappa-B ligand (RANKL; 100 ng/ml; PeproTech; Rocky Hill, NJ, USA) for 5 d. The medium of the cultures was changed every 3 d.
H. Tartrate-resistant acid phosphatase (TRAP) activity assay and staining in monocytes
The monocyte cultures were homogenized with lysis buffer [0.5M Tris-HCl (pH 9.0), 0.9% NaCl, 1% Triton X-100, 200 mM EDTA] and reacted at 37 °C using the TRACP & ALP Assay Kit (TaKaRa; Shiga Prefecture, Japan), and then, after 24 h, the absorbance of each well was measured at 405 nm using a microplate reader (BIO-RAD). For staining, the medium was carefully removed from the wells, and the cells were washed with PBS, fixed by incubation in 4% paraformaldehyde for 15 min at room temperature, rinsed with 3DW, and then stained using an Acid-Phosphatase Kit (100 μl/well; Sigma-Aldrich). The cells were rinsed again with 3DW, and positive TRAP staining was determined using a light microscope.
I. Co-culture of MC3T3-E1 cells and primary monocytes
Pre-osteoblast MC3T3-E1 cells were incubated in a 48-well plate (2×104
- 13 -
MC3T3-E1 cells were incubated for another 24 h. The MC3T3-E1 cells and primary-cultured monocyte cells were differentiated by the addition of differentiation media that contained ascorbic acid (50 μg/ml) and β-glycerophosphate (10 mM), treated with different concentrations of Loganin (1, 5, or 10 μg/ml), and then incubated for an additional 5 d. The medium of the co-cultures was changed every 3 d.
J. Ovariectomized model
Eight-week-old ovariectomized (OVX, n=24) and sham-operated (Sham, n=8) female ddY mice (Shizuoka Laboratory Center Inc., Hamamatsu, Japan) were maintained on a diet of Formula-M07 (5.0 g/d; Feedlab Co., Ltd., Hanam, Korea) and tap water (15 ml/day) and were individually housed in transparent plastic cages with controlled temperature (23 ± 2° C), humidity (55 ± 5%), and illumination (12-h p(12-hotoperiod). T(12-he mice were treated wit(12-h different concentrations of Loganin (2, 10, or 50 mg/kg/d) through oral administration using a needle for 12 weeks, and bone mineral density (BMD) was measured before and after treatment. The animal research protocol was approved by the Animal Care and Use Committee of the Ajou University School of Medicine, and all experiments were conducted in accordance with the institutional guidelines established by the Committee.
- 14 -
K. Measurement of bone mineral density (BMD)
After anesthetization the mice using tiletamine/zolazepam (Zoletil; Virbac Laboratories, Carros, France) and placing the mice on specimen trays, all in the same position, the BMD and BMC of one femur from each animal was measured using a mus bone densitometer (GE Lunar, Madison, WI, USA) and on-board PIXI-mus software for small animals.
L. Microcomputed tomography (Micro CT) and SPECT scan
Transverse CT images of the sample were obtained using a Micro-CT scanner in the GSTEP (Inveon, Siemens, Germany), with a voltage of 60 kV, current of 400 μA, exposure time of 400 ms, magnification of M-H, CCD readout of 1280×1280, and rotation steps of 360. Reconstruction was performed using Inveon Research Workplace and COBRA Exxim (Siemens). Two-dimensional axial and 3-D images were reconstructed for qualitative and quantitative analyses. For region of interest (ROI) measurement and analyses, axial images were displayed using Inveon Research Workplace (Siemens). Axial reformats were performed to allow slice-by-slice, manual tracing of the contours of trabecular bone. For cross-sectional analysis of trabecular bone, a region that was ~300 μm below the growth plate was analyzed, and a variety of histomorphometric parameters, including relative bone volume
- 15 -
(BV/TV, %) and trabecular number (Tb.N, mm), thickness (Tb.Th, mm), and spacing (Tb.Sp, mm), were measured. For the SEPCT scan, mice were anesthetized using isoflurane/N2O/O2, and Tc-99m HDP (Mallinckrodt Medical B.V, Netherlands) was
injected into the tail vein of each specimen. In vivo planar images were obtained using an Inveon SPECT scanner (Siemens Preclinical Solutions, Malvern, PA, USA) that was equipped with a low-energy, all-purpose collimator. Images were collected until 100,000 were acquired for each specimen.
M. Blood sampling and serum OPG and RANKL measurement
Immediately after the anesthetized mice were euthanized, blood samples were collected from the left ventricle of each mouse using syringes. Each blood sample was transferred to an individual heparinized tube, and the serum was separated by centrifugation (1200 ×g for 15 min at 4 °C) and then stored at -20 °C until further analysis. The serum levels of OPG and RANKL were assessed using a Luminex multiplex assay (Merck Millpore, USA), according to the manufacturer’s instructions. The ratio of OPG and RANKL (OPG/RANKL) was used as a measure of bone density maintenance.
- 16 -
N. Statistical analysis
For each experimental condition, at least three separate experiments were performed by triplicate. Data are expressed as the mean ± standard deviation. Statistical analysis was performed using ABI Prism 7000 Sequence Detection System Statistics. Statistical differences were analyzed using Student’s t-test. A probability value (p) less than 0.05 (p<0.05) was considered statistically significant. Comparisons of multiple groups were done with a one-way analysis of variance (ANOVA), followed by Tukey’s HSD (honest significant difference) post hoc test for correction of multiple comparisons.
- 17 -
III. RESULTS
A. Cellular differentiation of osteoblastic ME3T3-E1 cells
Sixty-six plant extracts were screened for identifying a natural product that could effectively induce bone formation using pre-osteoblast cell line, MC3T3-E1, and the murine mesenchymal stem cell line, C3H10T½ (Table 1). Eight of the plant extracts increased ALP activity, which is responsible for effective bone formation, and the extract of Cornus officinalis induced the highest level of ALP activity without toxic effect. Therefore, the present study investigated the identity and effectiveness of the bioactive compounds in Cornus officinalis extracts.
- 18 -
Table 1. Alkaline phosphatase (ALP) activity of MC3T3-E1 cells treated with extracts from 66 plant species native to Korea.
- 19 -
B. Bioactive component of the Cornus officinalis extract
We reported that an extract of Cornus officinalis enhanced the differentiation of osteoblasts from pre-osteoblast MC3T3-E1 cells and inhibited the differentiation of osteoclasts from primary monocytes. Therefore, the goal of the present study was to further investigate the bone growth-promoting compounds found in Cornus
officinalis extracts (Figure 2). All fractionation was accessed by ALP activity assay
in the osteoblast cells at different concentrations (10, 50, and 100 μg/ml). The final bio-active sub-fraction constituent was analyzed by proton nuclear magnetic resonance(1H-NMR), carbon-13 nuclear magnetic resonance (13C-NMR), and mass
spectrometry analyses (Figure 3). The IUPAC name of the bioactive compound, (1S,4aS,6S,7R,7aS)-6-Hydroxy-7-methyl-1-[(2S,3R,4S,5S,6R)-3,4,5-trihydroxy-6-
- 20 -
- 21 -
C. Effect of Loganin on osteoblast differentiation and mineralized nodule formation
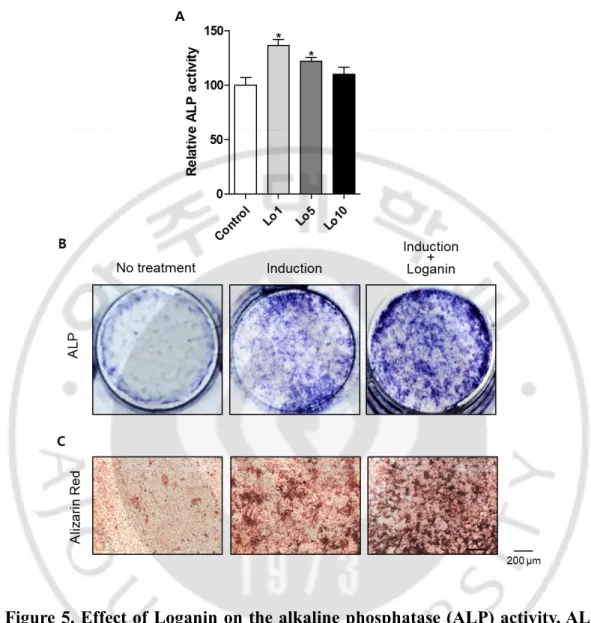
To investigate the effect of Loganin on osteoblast differentiation, MC3T3-E1 cells were incubated with different concentrations of Loganin (1, 5, or 10 μg/ml) for 3 d, and bone formation was assessed using an ALP activity assay and ALP staining. Loganin treatment significantly increased the ALP activity of the MC3T3-E1 cells, and the high- Loganin (1 μg/ml) treatment significantly increased ALP staining. To examine the effect of Loganin on osteoblast mineralization, pre-osteoblast cells were incubated with Loganin for 21 d, after which the mineralized nodules were measured using Alizarin Red S reagent, which is commonly used to measure calcium deposits in mineralized osteoblast cells (Mori et al., 1997). Loganin treatment increased Alizarin Red S staining, thereby indicating that Loganin enhances the formation of mineralized nodules in osteoblast cells (Figure 5C). These results indicate that Loganin induces osteoblast differentiation and mineralization in MC3T3-E1 cells.
- 22 -
Figure 5. Effect of Loganin on the alkaline phosphatase (ALP) activity, ALP staining, and Alizarin Red S staining of MC3T3-E1 cells. (A) ALP activity,
assessed after the induction of osteoblast differentiation. *: p < 0.05 vs. control. (B) ALP staining (with BCIP/NBT), performed after the induction of osteoblast differentiation. (C) Alizarin Red S staining of mineralized nodule formation cells.
- 23 -
D. Effect of Loganin on the expression of bone remodeling markers
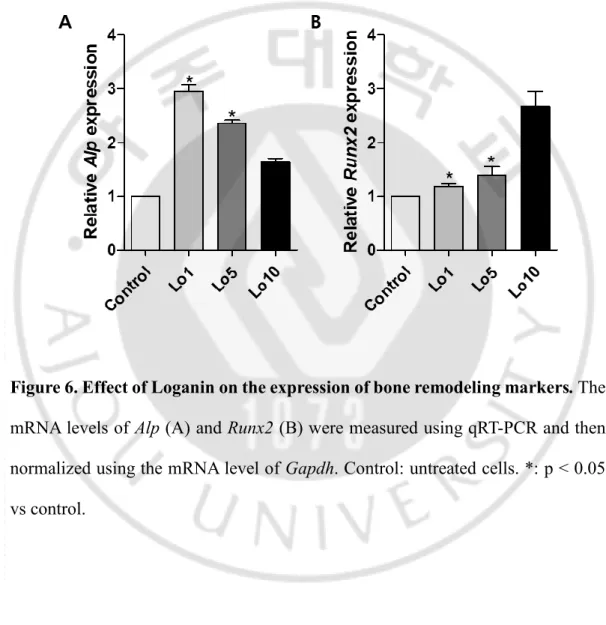
To measure the effect of Loganin on bone remodeling markers, MC3T3-E1 cells were incubated with Loganin for 3 d, and then the expression levels of Alp and
Runx2 were measured using quantitative RT-PCR (qRT-PCR), as previously
suggested (Balint et al., 2001; Komori, 2011; Neve et al., 2013). Loganin significantly increased the expression of Alp and Runx2 (Figure 6A and B), which suggests that Loganin promotes osteoblast differentiation and mineralized nodule formation by up-regulating bone-remodeling markers (Figure 6B).
- 24 -
Figure 6. Effect of Loganin on the expression of bone remodeling markers. The
mRNA levels of Alp (A) and Runx2 (B) were measured using qRT-PCR and then normalized using the mRNA level of Gapdh. Control: untreated cells. *: p < 0.05 vs control.
- 25 -
E. Effect of Loganin on osteoclast differentiation
To investigate the effect of Loganin on bone resorption, the differentiation of osteoclasts from mouse-derived monocytes was measured using a TRAP activity assay and TRAP staining. Monocytes, which are monocytic precursors, differentiate into osteoclasts (Nakagawa et al., 2015). The successful isolation of monocytes was confirmed by FACS, using a monocyte-specific antibody (anti-CD11b antibody) (Wagner et al., 2014) (Figure 7A). Loganin decreased TRAP activity and staining in a dose-dependent manner (Figure 7B and C), which suggests that Loganin inhibits the differentiation of osteoclasts from monocytes.
- 26 -
Figure 7. Effects of Loganin on osteoclast differentiation in primary-cultured monocytes. (A) Verification of identity for monocytes from isolated from
7-week-old mouse bone marrow, using fluorescence-activated cell sorting analysis. (B, C) Tartrate-resistant acid phosphatase (TRAP) activity and staining in monocytes. *: p < 0.05 vs. control.
- 27 -
F. Effect of Loganin on osteoblast and osteoclast differentiation
To investigate the effect of Loganin on bone remodeling in physiological conditions where osteoblasts and osteoclasts coexist, mouse-derived monocytes and MC3T3-E1 cells were co-cultured for 24, 48, 72, and 120 h. The ALP activity of Loganin-treated cells increased earlier (at 24 h) than that of the control cells (at 72 h; Figure 8A). The TRAP activity of the control cells increased in a time-dependent manner, whereas that of the Loganin -treated cells decreased (Figure 8B). Therefore, Loganin increased the differentiation of osteoblasts and decreased the differentiation of osteoclasts.
- 28 -
Figure 8. Effect of Loganin on the alkaline phosphatase (ALP) and tartrate-resistant acid phosphatase (TRAP) activity of co-cultured cells MC3T3-E1 and monocytes. Alkaline phosphatase (ALP) activity (A). Tartrate-resistant acid
phosphatase (TRAP) activity (B). Control: untreated control of Loganin. *: p < 0.05 vs control.
- 29 -
G. Effect of Loganin on OVX-induced bone loss in an osteoporotic mouse model
The in vitro results confirmed the anti-osteoporotic effect of Loganin in OVX mice. BMD was measured using a PIXI-mus bone densitometer at 0, 6, and 12 weeks. The BMD of the OVX group was significantly lower than that of the Sham group at both 6 and 12 weeks. However, Loganin inhibited OVX-induced BMD loss in a density-dependent manner (Figure 9). For further analysis of bone loss, Micro CT and SPECT scan analysis were performed. For cross-sectional analysis of trabecular bone, a region that was ~300 μm below the growth plate was analyzed using micro CT analysis, and a variety of histomorphometric parameters, including relative bone volume (BV/TV, %) and trabecular number (Tb.N, 1/mm), thickness (Tb.Th, mm), and spacing (Tb.Sp, mm) were measured (Trivedi et al., 2009). SrCl2
was used as a positive control (Fernandez et al., 2014). The bone structure of the OVX group was impaired, as indicated by relatively low BV/TV, Tb.N, and Tb.Th values and relatively high Tb.Sp values, when compared to the Sham group. However, both the positive control (SrCl2) (10 mg/kg/day) and Loganin (50
mg/kg/day) treatments improved the BV/TV, Tb.N, Tb.Th, and Tb.Sp values. Thus, Micro CT revealed that Loganin inhibits the osteoporotic bone loss and impaired bone structural properties of OVX mice (Figure 10). The femoral bone scan images confirmed that the bone loss of the OVX group was greater than that of the Sham group but less than that of the SrCl2- and Loganin -treated groups (Figure 11).
- 30 -
Technetium Tc-99m HDP and an Inveon SPECT scanner that was equipped with a low-energy, all-purpose collimator were used to scan the mouse skeletons. Images were collected until 100,000 were acquired for each specimen. Technetium Tc-99m HDP is absorbed into bone and fluoresces blue when excited. High radioactive signals detected in the bladder are the radioisotopes that are not interacted with bone and thus gathered in the bladder. The Tc-99m HDP level of the OVX group was lower than that of the Sham group, and the Tc-99m HDP level of the Loganin-treated group was greater than that of the OVX group (Figure 12), which suggests that Loganin prevents osteoporotic bone loss.
- 31 -
Figure 9. Effects of Loganin on the bone mineral density (BMD) of ovariectomized (OVX) mice. (A) BMD after 6 and 12 weeks. (B) BMD after 6
and 12 weeks. (C) BMD after 12 weeks. *: p < 0.05, **: p < 0.005, ***: p < 0.001 vs OVX.
- 32 -
Figure 10. Anti-osteoporosis effect of Loganin on the bones of ovariectomized (OVX) mice. (A) Relative bone volume (BV/TV). (B) Trabecular thickness (Tb.Th).
- 33 -
Figure 11. Anti-osteoporosis effect of Loganin on the femurs of ovariectomized (OVX) mice. Vertical and horizontal cross-sections.
- 34 -
Figure 12. Anti-osteoporosis effect of Loganin in ovariectomized (OVX) mice.
The anti-osteoporotic effect of Loganin was visualized using Inveon SPECT scanning.
- 35 -
H. Effect of Loganin on serum OPG and RANKL levels
To investigate the effect of Loganin on metabolic bone markers in mice sera, serum levels of OPG and RANKL were measured using ELISA. The OVX group yielded greater serum OPG and RANKL levels than the Sham group, and Loganin increased and decreased serum OPG and RANKL, respectively (Figure 13. A, B). Therefore, Loganin increased the OPG/RANKL ratio, which indicates that Loganin inhibits osteoclastogenesis (Figure 13. C) and suggests that Loganin prevents osteoporotic bone loss.
- 36 -
Figure 13. Anti-osteoporosis effect of Loganin on serum levels of OPG and RANKL in ovariectomized (OVX) mice. (A) Serum levels of OPG. (B) Serum
- 37 -
IV. DISCUSSION
We demonstrated that an ethanol extract of Cornus officinalis increased the differentiation of osteoblasts from pre-osteoblast cells and decreased the differentiation of osteoclasts from primary-cultured monocyte cells. The goals of the present study were to isolate the bioactive compound(s) responsible for the anti-osteoporotic effects of the Cornus officinalis extract and to investigate the
anti-osteoporotic effects of the isolated compound(s) in vitro and in vivo. Loganin, which is known to prevents diabetes and related symptoms and to improve brain function (Ahn et al., 2017), was isolated as the only candidate bioactive component. Loganin stimulated the maturation and differentiation of osteoblast cells from early to terminal stages in the whole differentiation process (Li et al., 2010). ALP that glycoprotein found on the osteoblasts surface, increase bone formation activity (Sardiwal et al., 2013). In addition, mineralization involves the accumulation of calcium and minerals to form bones (Gautam et al., 2011; Bonucci, 2012).
In the present study, Loganin significantly increased the ALP activity, ALP staining, and mineralized nodule formation of MC3T3-E1 cells. ALP increases during active bone formation, along with the induction of osteoblast activity, which suggests that ALP plays an important role in the mineralization of newly formed bone (Watts, 1999). Runx2 is a key transcription factor related to osteoblast
- 38 -
differentiation (Gilbert et al., 2002). Loganin increased the expression of osteoclast markers (Alp and Runx2) by MC3T3-E1 cells, which suggests that Loganin promotes osteoblast differentiation and mineralization by up-regulating osteoblast markers during new bone formation.
Bone remodeling involves a balance between bone formation and bone resorption. Bone resorption is caused by the differentiation of osteoclast cells, the osteoclasts conduct through degradation and removal of both the inorganic mineral and organic matrix (Kikuta and Ishii, 2013). Tartrate-resistant acid phosphatase (TRAP) is considered a histochemical osteoclast marker (Agrawal et al., 2010; Kim and Kim, 2016; Salamanna et al., 2016). The differentiation of osteoclasts, which was measured indirectly by measuring TRAP activity, was suppressed by Loganin in both primary cultured monocyte cells and in the co-culture of pre-osteoblasts and osteoclast precursors. These data indicate that Loganin enhances osteoblast differentiation in pre-osteoblast cells and inhibits osteoclast differentiation in monocyte cells. Therefore, Loganin could enhance bone formation and suppress resorptive bone loss.
To confirm the in vitro results, an osteoporotic mouse model was used to investigate the anti-osteoporotic effects of Loganin in vivo. The OVX mouse model is well known as human postmenopausal women and is characterized by low bone mass and BMD (Wu et al., 2013). BMD is often used as an index to quantify
- 39 -
osteoporosis and, in the present study, was measured using PIXI-mus (Vasikaran et al., 2011). As expected, the BMD of the OVX mice was lower than that of the Sham-operated mice. However, the BMD of the Loganin-treated group indicated that Loganin inhibits OVX-induced bone loss. The main histomorphometric parameters, which include relative bone volume (BV/TV, %), trabecular number (Tb.N, 1/mm), trabecular thickness (Tb.Th, mm), and trabecular spacing (Tb.Sp, mm), were used to assess the degree of osteoporosis (Zhang et al., 2010). SrCl2, an anti-osteoporotic
agent that inhibits bone loss (Pemmer et al., 2011), was used as a positive control. Loganin improved all four of the histomorphometric parameters in OVX mice, thereby confirming the anti-osteoporosis effect of Loganin in vivo. In addition, the OVX group exhibited osteoporosis that presented bone loss in femoral bone Micro CT. However, the positive control (SrCl2) and Loganin treatments inhibited
OVX-induced bone loss. The bone loss observed in the OVX mice was also evaluated using Technetium Tc-99m HDP. After technetium Tc-99m HDP was performed intravenous injection in mice tail of Sham group and OVX group, Loganin oral-injection group, Inveon SPECT scan performed. As a result, the absorption of technetium Tc-99m HDP by the OVX group was lower than that of both the Sham group and Loganin -treated groups, which indicates that Loganin inhibits OVX-induced bone loss. Next, the serum levels of OPG and RANKL were measured (Wasilewska et al., 2010). OPG is an inhibitor of osteoclast differentiation, whereas RANKL is an osteoclast differentiation activity factor, and the degree of
- 40 -
osteoclastogenesis can be assessed using the OPG/RANKL ratio as an indicator (Hamed et al., 2014). In the present study, the SrCl2 and Loganin treatments
increased serum levels of OPG but decreased serum levels of RANKL, which suggests that Loganin promotes bone formation by enhancing the expression of bone markers. In addition, all in vivo experiments, the results of Loganin treatment were similar to those of the positive control, SrCl2 treatment. Taken together, the in vivo
results demonstrate that Loganin prevents osteoporotic bone loss and improves metabolic bone markers in OVX mice.
In summary, the in vitro results of the present study demonstrate that Loganin increases osteoblast differentiation, which is associated with bone formation, by up-regulating osteoblast-inducing markers but inhibits osteoclast differentiation, which is associated with bone resorption. Similarly, the in vivo results also demonstrate that Loganin inhibits osteoporotic bone loss. Therefore, Loganin is a potential therapeutic agent for treating osteoporosis.
- 41 -
V. Conclusion
The present study demonstrates that Loganin isolated from Cornus
officinalis increases the differentiation and mineralization of pre-osteoblast cells and
decreases the differentiation of pre-osteoclastic monocytes. Even in co-cultures of pre-osteoblast MC3T3-E1 cells and osteoclast-precursor monocytes, Loganin effectively enhanced osteoblast differentiation and inhibited osteoclast differentiation. In vivo experiments also demonstrated that Loganin inhibits osteoporotic bone loss.
In conclusion, Loganin apparently plays a significant role in enhancing bone formation and preventing osteoporotic bone loss and, thereby, represents a natural therapeutic agent that could potentially be used to treat osteoporosis.
- 42 -
REFERENCES
1. Addison WN, Nelea V, Chicatun F, Chien YC, Tran-Khanh N, Buschmann MD, Nazhat SN, Kaartinen MT, Vali H, Tecklenburg MM, Franceschi RT, McKee MD: Extracellular matrix mineralization in murine MC3T3-E1 osteoblast cultures: an ultrastructural, compositional and comparative analysis with mouse bone. Bone 71: 244-256, 2015
2. Agrawal A, Buckley KA, Bowers K, Furber M, Gallagher JA, Gartland A: The effects of P2X7 receptor antagonists on the formation and function of human osteoclasts in vitro. Purinergic Signal 6: 307-315, 2010
3. Ahn JH, Mo EJ, Jo YH, Kim SB, Hwang BY, Lee MK: Variation of loganin content in Cornus officinalis fruits at different extraction conditions and maturation stages. Biosci Biotechnol Biochem 81: 1973-1977, 2017
4. Balint E, Szabo P, Marshall CF, Sprague SM: Glucose-induced inhibition of in vitro bone mineralization. Bone 28: 21-28, 2001
5. Bernhardt A, Schamel M, Gbureck U, Gelinsky M: Osteoclastic differentiation and resorption is modulated by bioactive metal ions Co2+, Cu2+ and Cr3+ incorporated into calcium phosphate bone cements. PLoS One 12: e0182109, 2017
- 43 -
6. Bhargavan B, Gautam AK, Singh D, Kumar A, Chaurasia S, Tyagi AM, Yadav DK, Mishra JS, Singh AB, Sanyal S, Goel A, Maurya R, Chattopadhyay N: Methoxylated isoflavones, cajanin and isoformononetin, have non-estrogenic bone forming effect via differential mitogen activated protein kinase (MAPK) signaling. J Cell Biochem 108: 388-399, 2009
7. Bonucci E: Bone mineralization. Front Biosci (Landmark Ed) 17: 100-128, 2012 8. Fernandez JM, Molinuevo MS, McCarthy AD, Cortizo AM: Strontium ranelate stimulates the activity of bone-specific alkaline phosphatase: interaction with Zn(2+) and Mg (2+). Biometals 27: 601-607, 2014
9. Gautam AK, Bhargavan B, Tyagi AM, Srivastava K, Yadav DK, Kumar M, Singh A, Mishra JS, Singh AB, Sanyal S, Maurya R, Manickavasagam L, Singh SP, Wahajuddin W, Jain GK, Chattopadhyay N, Singh D: Differential effects of formononetin and cladrin on osteoblast function, peak bone mass achievement and bioavailability in rats. J Nutr Biochem 22: 318-327, 2011
10. Gilbert L, He X, Farmer P, Rubin J, Drissi H, van Wijnen AJ, Lian JB, Stein GS, Nanes MS: Expression of the osteoblast differentiation factor RUNX2 (Cbfa1/AML3/Pebp2alpha A) is inhibited by tumor necrosis factor-alpha. J Biol
Chem 277: 2695-2701, 2002
11. Hadjidakis DJ, Androulakis, II: Bone remodeling. Ann N Y Acad Sci 1092: 385-396, 2006
- 44 -
12. Hamed SA, Moussa EM, Youssef AH, Abd ElHameed MA, NasrEldin E: Bone status in patients with epilepsy: relationship to markers of bone remodeling.
Front Neurol 5: 142, 2014
13. Huang J, Zhang Y, Dong L, Gao Q, Yin L, Quan H, Chen R, Fu X, Lin D: Ethnopharmacology, phytochemistry, and pharmacology of Cornus officinalis Sieb. et Zucc. J Ethnopharmacol 213: 280-301, 2017
14. Ishtiaq S, Fogelman I, Hampson G: Treatment of post-menopausal osteoporosis: beyond bisphosphonates. J Endocrinol Invest 38: 13-29, 2015
15. Kikuta J, Ishii M: Osteoclast migration, differentiation and function: novel therapeutic targets for rheumatic diseases. Rheumatology (Oxford) 52: 226-234, 2013
16. Kim JH, Kim N: Signaling Pathways in Osteoclast Differentiation. Chonnam
Med J 52: 12-17, 2016
17. Komori T: Signaling networks in RUNX2-dependent bone development. J Cell
Biochem 112: 750-755, 2011
18. Krane SM: Identifying genes that regulate bone remodeling as potential therapeutic targets. J Exp Med 201: 841-843, 2005
19. Li M, Wang W, Wang P, Yang K, Sun H, Wang X: The pharmacological effects of morroniside and loganin isolated from Liuweidihuang Wan, on MC3T3-E1 cells. Molecules 15: 7403-7414, 2010
- 45 -
20. Liu W, Mao L, Ji F, Chen F, Wang S, Xie Y: Icariside II activates EGFR-Akt-Nrf2 signaling and protects osteoblasts from dexamethasone. Oncotarget 8: 2594-2603, 2017
21. Livak KJ, Schmittgen TD: Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 25: 402-408, 2001
22. Ma W, Wang KJ, Cheng CS, Yan GQ, Lu WL, Ge JF, Cheng YX, Li N: Bioactive compounds from Cornus officinalis fruits and their effects on diabetic nephropathy. J Ethnopharmacol 153: 840-845, 2014
23. Moreira LD, Oliveira ML, Lirani-Galvao AP, Marin-Mio RV, Santos RN, Lazaretti-Castro M: Physical exercise and osteoporosis: effects of different types of exercises on bone and physical function of postmenopausal women. Arq Bras
Endocrinol Metabol 58: 514-522, 2014
24. Mori S, Harruff R, Ambrosius W, Burr DB: Trabecular bone volume and microdamage accumulation in the femoral heads of women with and without femoral neck fractures. Bone 21: 521-526, 1997
25. Nakagawa T, Ohta K, Kubozono K, Ishida Y, Naruse T, Takechi M, Kamata N: Zoledronate inhibits receptor activator of nuclear factor kappa-B ligand-induced osteoclast differentiation via suppression of expression of nuclear factor of activated T-cell c1 and carbonic anhydrase 2. Arch Oral Biol 60: 557-565, 2015
- 46 -
26. Neve A, Corrado A, Cantatore FP: Osteocalcin: skeletal and extra-skeletal effects. J Cell Physiol 228: 1149-1153, 2013
27. Park E, Kim MC, Choi CW, Kim J, Jin HS, Lee R, Lee JW, Park JH, Huh D, Jeong SY: Effects of Dihydrophaseic Acid 3'-O-beta-d-Glucopyranoside Isolated from Lycii radicis Cortex on Osteoblast Differentiation. Molecules 21, 2016
28. Pemmer B, Hofstaetter JG, Meirer F, Smolek S, Wobrauschek P, Simon R, Fuchs RK, Allen MR, Condon KW, Reinwald S, Phipps RJ, Burr DB, Paschalis EP, Klaushofer K, Streli C, Roschger P: Increased strontium uptake in trabecular bone of ovariectomized calcium-deficient rats treated with strontium ranelate or strontium chloride. J Synchrotron Radiat 18: 835-841, 2011
29. Salamanna F, Pagani S, Maglio M, Borsari V, Giavaresi G, Martelli AM, Buontempo F, Fini M: Estrogen-deficient osteoporosis enhances the recruitment and activity of osteoclasts by breast cancer cells. Histol Histopathol 31: 83-93, 2016
30. Sardiwal S, Magnusson P, Goldsmith DJ, Lamb EJ: Bone alkaline phosphatase in CKD-mineral bone disorder. Am J Kidney Dis 62: 810-822, 2013
31. Tella SH, Gallagher JC: Prevention and treatment of postmenopausal osteoporosis. J Steroid Biochem Mol Biol 142: 155-170, 2014
32. Trivedi R, Kumar A, Gupta V, Kumar S, Nagar GK, Romero JR, Dwivedi AK, Chattopadhyay N: Effects of Egb 761 on bone mineral density, bone
- 47 -
microstructure, and osteoblast function: Possible roles of quercetin and kaempferol. Mol Cell Endocrinol 302: 86-91, 2009
33. Vasikaran S, Eastell R, Bruyere O, Foldes AJ, Garnero P, Griesmacher A, McClung M, Morris HA, Silverman S, Trenti T, Wahl DA, Cooper C, Kanis JA, Group I-IBMSW: Markers of bone turnover for the prediction of fracture risk and monitoring of osteoporosis treatment: a need for international reference standards. Osteoporos Int 22: 391-420, 2011
34. Wagner M, Koester H, Deffge C, Weinert S, Lauf J, Francke A, Lee J, Braun-Dullaeus RC, Herold J: Isolation and intravenous injection of murine bone marrow derived monocytes. J Vis Exp, 2014
35. Wasilewska A, Rybi-Szuminska A, Zoch-Zwierz W: Serum RANKL, osteoprotegerin (OPG), and RANKL/OPG ratio in nephrotic children. Pediatr
Nephrol 25: 2067-2075, 2010
36. Watts NB: Clinical utility of biochemical markers of bone remodeling. Clin
Chem 45: 1359-1368, 1999
37. Weilbaecher KN, Guise TA, McCauley LK: Cancer to bone: a fatal attraction.
Nat Rev Cancer 11: 411-425, 2011
38. Wu K, Lin TH, Liou HC, Lu DH, Chen YR, Fu WM, Yang RS: Dextromethorphan inhibits osteoclast differentiation by suppressing RANKL-induced nuclear factor-kappaB activation. Osteoporos Int 24: 2201-2214, 2013
- 48 -
39. Zhang ZM, Li ZC, Jiang LS, Jiang SD, Dai LY: Micro-CT and mechanical evaluation of subchondral trabecular bone structure between postmenopausal women with osteoarthritis and osteoporosis. Osteoporos Int 21: 1383-1390, 2010
40. Zheng M, Ge Y, Li H, Yan M, Zhou J, Zhang Y: Bergapten prevents lipopolysaccharide mediated osteoclast formation, bone resorption and osteoclast survival. Int Orthop 38: 627-634, 2014
- 49 - - 국문요약 -
Lonanin 의 조골세포 분화 촉진 및 파골세포
분화 억제 기전 규명
아주대학교 대학원 의생명과학과 여 수 빈 (지도교수: 정 선 용) 골다공증은 세계보건기구(WHO)에서 ‘골량의 감소와 미세구조의 이상을 특징으로 하는 전신적인 골격계 질환으로, 결과적으로 뼈가 약해져서 부러지기 쉬운 상태가 되는 질환’으로 정의한다. 골다공증은 호르몬의 변화, 노화, 약물복용 등의 환경 및 유전 인자들의 상호작용에 의해 발생하는 다인자성 복합 질환이며, 최근 고령화의 진전에 따른 골다공증 환자수의 급격한 증가 현상이 나타나고 있으며, 세계보건기구의 자료를 인용하면 인구 전반의 고령화 진전의 영향으로 2050 년에 이르면 골다공증 유병률이 지금의 4 배 정도까지 증가하게 될 것으로 예상하고 있다. 골다공증 치료를 위해서는 장기적인 약물 치료가 필요한데, 현재 많이 사용되는 골다공증 치료제로는 비스포스포네이트, 호르몬제 등이 있으나, 부작용이 있어 장기 투여가 어려우며 새로운 뼈- 50 - 생성에 의한 골밀도 증가에는 큰 역할을 하지 않는다는 단점이 있다. 이에 부작용이 적어 장기간의 사용이 가능한 천연물 유래의 약물 치료제 개발에 대한 필용성이 있어왔다. 선행연구에서 66 종의 약용식물 유래 에탄올 추출물을 대상으로 조골세포주(MC3T3-E1)의 분화능 촉진 효능을 탐색하여, 가장 효과가 좋은 산수유 추출물(Cornus officinalis)을 발굴하였다. 본 연구에서는 산수유 추출물에서 조골세포의 분화를 촉진하는 효능성분(bioactive compound)을 분리 ∙ 동정하고 그 효과를 in vitro 와 in vivo 실험을 통해 조사하고자 하였다.
산수유 에탄올 추출물을 다양한 용매를 사용한 분획(fractionation)과 질량스펙트럼 방법을 통해 분석한 결과, 효능성분으로 Loganin 을 동정하였다. Loganin 이 골 형성(bone formation)에 관여하는 조골세포(osteoblast)의 분화와 골 흡수(bone resorption)에 관여하는 파골세포(osteoclast)의 분화에 미치는 영향을 각각 연구하였으며, 실제 생체 내와 유사한 환경인 조골세포와 파골세포의 공배양(co-culture) 조건에서 Loganin 이 이들 세포의 분화에 미치는 영향을 연구하였다. 조골세포의 전구세포인 조골모세포주 MC3T3-E1 에 Loganin 을 처리하여 조골세포 분화시에 나타나는 alkaline phosphatase(ALP) 활성을 측정한 결과, 조골세포의 분화 증가의 효과가 있었으며, 석회화 결절형성(mineralized nodule formation)eh 유의하게 증가하였다. 또한, Quantitative reverse-transcription PCR (qRT-PCR)를 통해
- 51 -
조골세포 분화 마커인 Alp, Runx2 의 mRNA 발현량을 측정한 결과, Loganin 을 처리한 군에서 대조군에 비해 모두 유의하게 증가되었다. 마우스 골수에서 분리 ∙ 배양된 초대 배양 단핵세포(primary-cultured monocyte)에 Loganin 을 처리하여 파골세포 분화시에 나타나는 Tartrate-resistant acid phosphatase(TRAP) 활성을 분석한 결과, 파골세포 분화 억제에 유의한 효과가 있었다. 또한, 공배양 조건에서는 Loganin 의 처리에 의해 조골세포의 분화 증가와 파골세포의 분화 억제가 관찰되었다.
마지막으로, 난소를 절제한 골다공증 유도 마우스(ovariectomized, OVX) 실험을 통해 Loganin 의 in vivo 에서의 효과를 조사하였다. Loganin 을 3 가지 농도(2, 10 and 50 mg/kg/days)로 12 주 동안 구강주입(oral injection) 실험을 통해, Loganin 10 mg/kg/days 의 농도에서 골밀도(bone mineral density), 혈액에서의 OPG/RANKL 비율, micro-CT 와 뼈 스캔(SPECT scan)에서의 뼈의 미세구조 및 뼈 굵기 등의 전반적인 골다공증 평가 자료에서 유의한 개선 효과가 있음을 증명하였다. 결론적으로, 산수유 에탄올 추출물의 효능성분인 Loganin 은 조골세포의 분화 증진을 통한 뼈 형성 촉진과 파골세포의 분화억제를 통한 뼈 흡수 억제에 효능이 있음을 증명하였다. 또한, 동물실험에서 Loganin 이 난소절제에 의해 나타나는 골다공증의 개선 또는 치료에 좋은 효과가 있음을 밝혔다. 본 연구결과가 향후 Loganin 를 이용한 골다공증 치료제 개발에 활용될 수 있기를 기대한다.
- 52 -
__________________________________________________________________ 핵심어: 천연물 추출물, 효능성분, 골다공증, Loganin, 조골세포,