153 http://dx.doi.org/10.4196/kjpp.2012.16.3.153
ABBREVIATIONS: MTT, 3-(4,5-Dimethylthizol-2-yl)-2,5-diphenylte- trazolium bromide; BCA, bicinchoninic acid; FACS, fluorescence activated cell sorter; PARP, Poly (ADP-ribose) polymerase; PMSF, phenylmethanesulfonylfluoride or phenylmethylsulfonyl fluoride;
PBS, phosphate buffered saline.
Received November 17, 2011, Revised May 1, 2012, Accepted May 11, 2012
Corresponding to: Hoon Yoo, Department of Pharmacology and Dental Therapeutics, School of Dentistry, Chosun University, 375, Seosuk-dong, Dong-gu, Gwangju 501-759, Korea. (Tel) 82-62-230- 6894, (Fax) 82-62-230-6894, (E-mail) [email protected]
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://
creativecommons.org/licenses/by-nc/3.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
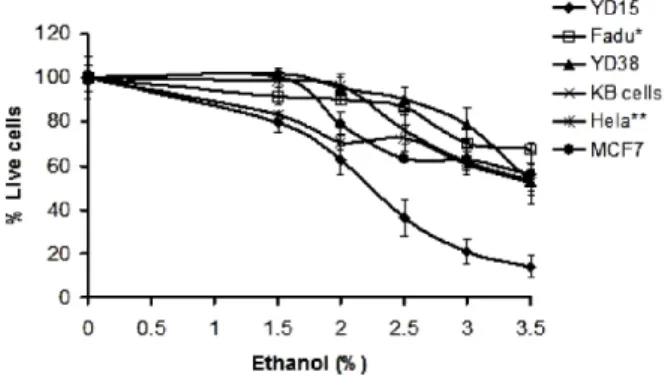
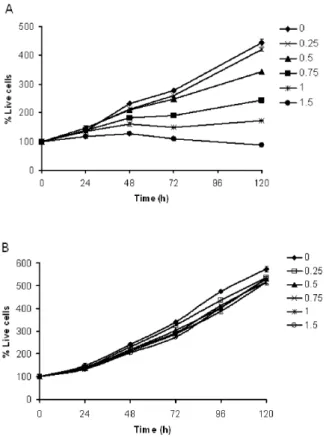
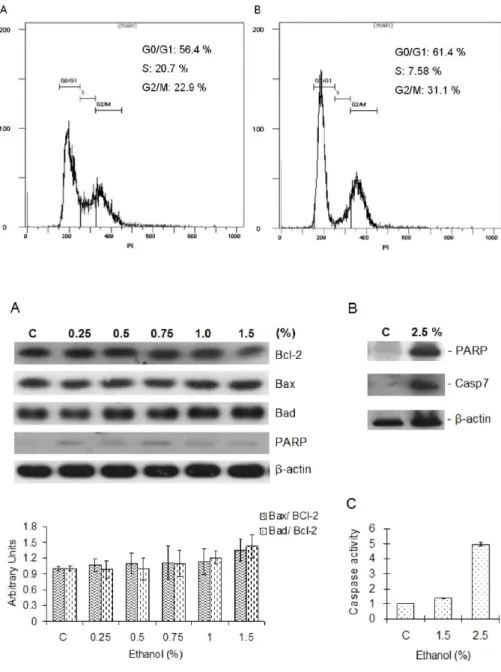
Ethanol Elicits Inhibitory Effect on the Growth and Proliferation of Tongue Carcinoma Cells by Inducing Cell Cycle Arrest
Thanh-Do Le
1, Thi Anh Thu Do
1, Rina Yu
2, and Hoon Yoo
11