*Corresponding Author : Sang-Ki Choi([email protected]) Received March 13, 2018
Accepted May 20, 2018
Revised April 26, 2018 Published May 28, 2018
알코올성 간 손상 조직에서 TGF-β
1와 c-Myc , Erb-B2 , Thymosin-β
4유전자 발현 융합 연구
김진수1, 최상기2*
1대전보건대학교 임상병리과, 2국립순천대학교 생물학과
A study of Expression of TGF-β1, c-Myc, Erb-B2 and Thymosin-β4 Gene in Alcoholic Liver Damage Tissue.
Jean–Soo Kim1, Sang-Ki Choi2*
1Department of Clinical Laboratory Science, Daejeon Health Institute of Technology
2Department of Biology, Sunchon National University
요 약 본 연구는 25% 에탄올에 손상된 간조직에서 TGF-β1와 c-Myc, Erb-B2, Thymosin-β4유전자의 발현을 알아 보고 자 실시 하였다. 실험군은 2군으로 나누어 25% 에탄올로 간 손상을 유발한 실험군과 정제수를 투여한 대조군으로 나누어 실험하였다. 검사 결과는 25% 에탄올를 투여 했던 실험군은 대조군에 비하여 TGF-β1, c-Myc 및 Thymosin-β4유전자의 발현 증가를 알 수 있었으며 Erb-B2 유전자는 뚜렷한 발현을 알 수 없었다. 또한 손상된 간 조직에서 헤마톡실린 에오진 염색을 통한 세포 손상을 관찰 할 수 있었다. 결론적으로 기존 임상에서 간 기능 관련 질병 예방과 질환 판정 시 혈청학적, 조직학적 검사 외에 TGF-β1, c-Myc 및 Thymosin-β4의 분자 진단 기법에 의한 유전자 발현 상태를 융합 검사함으로써 간 질환 판정의 보조 자료로 활용 될 수 있을 것으로 사료 된다.
주제어 : 헤마톡실린 에오진 염색, TGF-β1, c-Myc, Erb-B2, Thymosin-β4, 융합
Abstract This study has been conducted to see the expression of TGF-β1, c-Myc, Erb-B2 and Thymosin-β4 genes in ethanol - damaged liver tissues. Experimental groups were divided into 2 groups, one where damaged liver was caused by 25% ethanol and normal group administered with purified water. Results of test showed the expression of TGF-β1, c-Myc, and Thymosin-β4genes was higher in the experimental group treated with 25% ethanol than in the normal group. Erb-B2 gene was not expressed clearly. Thus, it is considered that we can expect to utilize TGF-β1, c-Myc 및 Thymosin-β4as auxiliary data and find clinical meanings of diagnosis on hepatic diseases, In addition to serologic and histological examination by convergence examining the gene expression status by molecular diagnostic techniques in liver-related disease prevention and diagnosis through results of this study.
Key Words : Hematoxylin, Eosin, TGF-β1, c-Myc, Erb-B2, Thymosin-β4, convergence
1. 서론
최근 의료 기관에서 조직검사 관련 질병 진단 시 HER2 (위암, 유방암) 유전자, BRCA (유방암, 난소암), ALK (폐암), BRAF 돌연변이 유전자검사 (갑상선 유두
암), 그리고 P53 (암 억제유전자) 검사 등 FISH, SISH 분 자 진단 기법을 이용한 유전자 검사를 많이 시행하고 있 다. 그러나 간장은 ALT, AST, AFP등 혈액검사와 간 초 음파검사, CT검사 등을 통하여 진단이 주로 이루어지고 있다. 간장은 일반적으로 알코올 대사 과정 중 생성된 아
세트 알데히드나 반응성 강한 라디칼 등[1] 으로 혈중에 중성지방 상승에 따른 고지혈증, 간 경변 및 지방간 질환 을 유발한다[2]. 알코올은 보통 섭취 후 30분 안에 위에서 25%정도 흡수되며 나머지는 소장에서 흡수되어 간에서 주로 대사된다[3]. 대사 과정은 cytochrome P450 2E1, alcohol dehydrogenase (ADH), catalase, Microsomal Ethanol Oxidizing System (MEOS)에 의하여 acetaldehyde로 대 사 된다 [4-9]. 이때 생성된 acetaldehyde는 acetaldehyde dehydro-genase (ALDH)에 의해 acetic acid로 전환되는 데 이때 장애가 발생한다면 acetaldehyde 축적을 통한 변 이원성 및 발암을 유발한다[10]. 따라서 우리 몸은 catalase, superoxide dismutase (SOD), glutathione S-transferase (GST), glutathione peroxidase (GSH-px) 등 항산화 효소나 polyphenol, 비타민 같은 항산화제를 통해 세포 산화적인 손상을 감소 시킨다 [11,12].
Thymosin-β4는 송아지 흉선의 성장인자로 처음 알려 졌으며 [13-16] 크기는 4.9 kDa 이고 [17] ATP-G액틴과 1:1 비율로 결합함으로서 [15] G액틴 분리과정에 관여하 고 [18,19] 세포 운동성 [20,21], 혈관 형성과정 [22,23] 특 히 신생혈관 생성 초기 과정에 관여하며 [24] 배아세포 분화 및 발달 중인 뇌 [25-27], 모낭이 만들어지는 과정 [28], 암세포 [24], 그리고 암전이하는 과정 중 [29,30]에 발현 되는 것으로 알려져 있다. 특히 같은 암이라도 흑색 종 전이 [31]와 섬유육종 전이 [32]에서 높게 발현되는 것 으로 보고 되고 있다.
c-Myc (c- Myelocytomatosis oncogene) 유전자는 8 번 염색체 q24부분에 위치하고 Telomerase 발현과 세포 증식에 관여하는 유전자들에게 영향력을 끼치는 전사인 자로써 백혈병, 위암, 폐암 등에서 증폭되는 것으로 보고 되고 있다 [33].
Erb-B2 (erythroblastic leuemia virtual oncogene homolog 2) 유전자는 17번염색체 q11.2-12에 위치하며 난소암과 유방암의 25%에서 증폭이 일어나며 생존률 예 후가 나쁜 것으로 알려져 있다[33].
Transforming growth factor-β1 (TGF-β1) 유전자는 세포의 증식, 세포의 성장, 그리고 세포의 사멸과 분화 조 절에 관여 하는 유전자로 특히 만성 및 급성의 간세포 손 상으로 인한 1형 콜라겐 발현 시 동반되는 것으로 알려져 있으며 이는 TGF-β1증가가 간 섬유화의 시작을 의미한 다고 하겠다[34-40].
따라서 본 연구는 간 손상 시 조직에서의 혈액학적인
변화와 전자현미경적 구조 및 광학 현미경의 조직학적인 변화를 관찰하고 손상된 간 조직 내의 Thymosin-β4, c-Myc, Erb-B2, TGF-β1의 mRNA 발현의 차이를 관찰 함으로써 임상에 진단과 관련한 참고 자료로 활용하고자 하였다.
2. 재료 및 방법
2.1 간 손상 모델
실험동물은 평균체중 20~25g의 10주령 된 수컷 BALB/c 마우스를 노터스 회사로 부터 공급받아 1주일 동안 환경에 적응시켰으며, 알코올성 간 손상을 유발시 킬 목적으로 25% Ethanol 단독투여군 (n=5)과 대조군 (n=5)으로 분리 후 실험모델 이치호 방법 [41]을 채택하 여 실험군은 무수 Ethanol (99.9%, 덕산과학)을 희석한 25% Ethanol (5ml/kg)를 매일 1회, 8주 동안 구강 위관 영양법으로 투여하였으며. 8주 후 Ether로 마취하고 희 생시켰다.
2.2 혈청학적 검사
혈청학적 검사는 실험동물을 Ether로 마취하고 희생 시킨 후 주사기를 이용하여 700㎕ 이상을 심장에서 채취 하였으며 10분간 3,000×g, 4℃에서 원심 분리 후 ALP, ALT, AST 분석에 사용하였다.
2.3 Hematoxylin & Eosin 염색
실험동물에서 적출한 일부 간 조직을 10% 포르말린에 24시간 고정 하고 탈수 과정, 투명과정, 침투과정, 포매 과정을 거친 후, 생성된 블록을 microtome으로 4~6 μm 박절 하였다. 박절 시 나온 리본을 탈 파라핀 과정과 함 수 처리 및 염색을 실시 하였다. 핵 염색은 Harris- Hematoxylin (YD Diagnostics, korea), 세포질은 에오신 Y (Showa, Japan)로 염색을 하였다
2.4 전자현미경적 조직 관찰
실험동물을 Ether로 마취하고 희생시킨 후 간 조직 일 부를 세절한 후 Glutaraldehyde에 고정하여 전자현미경 용 표본을 제작하였다. 즉 Glutaraldehyde에 하루 정도 고정한 후 사산화 오스뮴에 2차 고정을 2시간 정도 실시 하였으며 수세 후 알코올을 이용한 저 농도에서 고 농도
(70% ~ 100%)로 탈수를 하였고, 산화프로필렌을 이용 한 치환 과정, 에폰812를 이용한 침투과정, 포매 틀을 이용 한 블록제작, 그리고 중합과정을 거친 후 Ultramicrotome 을 이용하여 박절 하였으며 초산 우라닐로 전자염색을 한 후 투과전자현미경을 이용하여 조직 관찰을 하였다
2.5 유전자 검사
실험동물에서 적출한 간 조직 일부를 세절한 후 -70℃
에서 냉동 보관하였다가 유전자 mRNA 발현 검사를 위 한 Real time PCR에 이용하였으며 유전자 결과 분석방 법은 케니스 방법 [42]을 적용하였다. RNA추출은 Homogenizer를 이용하여 조직을 파쇄한 후 B사 RNA Prep Kit를 이용하였고 역전사과정은 RNA 20 μl을 사용 하여 42°C에서 60 분간 반응시켰으며 전환된 cDNA는 Accupower Greenstar qPCR preMix Kit를 이용하여 Real time PCR 장비로 증폭하였다. 관련 과정은 cDNA, Primers Table 1, qPCR premix를 포함한 총량 50μl를 95°C에서 20초, 56°C에서 45초 과정으로 40번 증폭하였다.
Table 1. Primers used for the detection of gene expression
Target gene sequence (5´→3´) bp
Thymosin-β4
F TCT TTT CCC TCT CTG CT R AGA TGC AAA GAG GTT GG 111
c-Myc F CAC ATC AAT TTC TTC CT
R GTG CTG CAT GAG GAG AC 84
Erb-B2 F CAA CGT CAA TGA GTA AG
R AGG AGA TCA CAG GTT AC 131
TGF-β1
F ACG TCA CTG GAG TTG TA R CGT GGA GTT TGT TAT CT 141
2.6 통계분석
실험결과는 평균 ± 표준편차를 구하였으며 SPSS/
Windows 19.0을 이용하여 통계 처리하였다. 집단 간의 평균값 유의성 차이는 t 검정을 실시 하였으며 통계적인 유의성은 p = 0.05 수준에서 실시하였다.
3. 결과
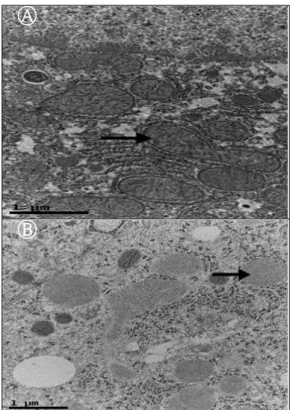
3.1 전자현미경적 조직관찰
정상 간 조직에서 미토콘드리아 등 세포질 내 소기관 은 정상적으로 분포하고 있었으며 특이할 만할 소견을
찾아볼 수 없었지만 25% Ethanol을 처리한 조직에서는 중심 정맥 주위에 용해소체들이 분포하고 있었다. 동시 에 미토콘드리아 상태는 속 사립체막(→)의 내부로 돌출 된 능선이 뚜렷하지 못하는 등 전체적으로 많은 손상이 있었다. Fig. 1
3.2 혈청 검사
ALT는 대조군에 비해 25% Ethanol 단독 투여 군에서 42.0 U/l로 높게 나타났으며 AST는 대조군에 비해 25%
Ethanol 단독 투여군에서 177.8 U/l로 높은 값을 보였다.
또한 ALP는 대조군에 비해 25% Ethanol 투여군 값이 97.4 U/l로 높다는 것을 알 수 있었다. 각 평균값에 대한 t 검정은 P < 0.05로 유의성이 있다는 것을 알 수 있었다.
Table 2
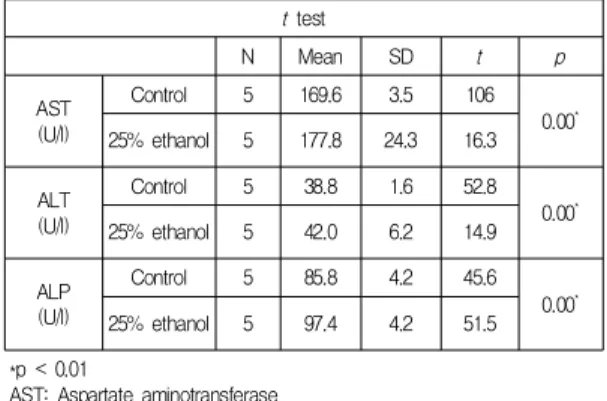
Table 2. Blood analysis to identify liver function t test
N Mean SD t p
AST (U/l)
Control 5 169.6 3.5 106
0.00* 25% ethanol 5 177.8 24.3 16.3
ALT (U/l)
Control 5 38.8 1.6 52.8
0.00*
25% ethanol 5 42.0 6.2 14.9
ALP (U/l)
Control 5 85.8 4.2 45.6
0.00*
25% ethanol 5 97.4 4.2 51.5
*p < 0.01
AST: Aspartate aminotransferase ALT: Alanine aminotransferase ALP: Alkaline phosphatase
3.3 Hematoxylin & Eosin 염색
H & E염색 결과는 대조군에 비해 25% Ethanol 단독 투여군에서 중심정맥 주위에 조직변성에 의한 핵농축 (↓)과 저 염색성 (→)을 일으켰으며 핵이 전체적으로 커 지는 반응세포핵 (←) 등 이상 세포 등을 볼 수 있었다.
Fig. 2
3.4 유전자 검사
Real time PCR을 통한 발현 연구 시 Ct값이 보통 18~32정도에서 이루어지는데 Erb-B2 유전자는 대체로 이 값을 벗어났으므로 대조군이나 25% Ethanol 투여군 에서 유전자 발현이 많지 않은 것을 알 수 있었다. Table
3 또한 Real time PCR 결과 분석방법인 케니스 방법 [42]
을 이용하여 housekeeping gene인β-actin 유전자 대조 군 발현정도를 1 기준으로 하였을때 25% Ethanol 투여군 발현양은 TGF-β1가 2.46, c-Myc 10.17, Thymosin-β4 2.46으로 유전자 발현이 증가 된 것을 알수 있었다. Table 4
Table 3. Cq values of gene expression in groups administered with 25% ethanol
Cq values = Ct
β-actin Erb-B2 TGF-β1 c-Myc Thymosin-β4
Control 20.18 33.58 27.96 29.88 23.60
25%
ethanol 19.24 32.52 25.72 25.59 21.36
Table 4. Relative expression of β-actin, Erb-B2, TGF- β1, c-Myc and Thymosin-β4gene expression
β-actin Erb-B2 ΔCt ΔΔCt 2-ΔΔCt
Control 20.18 33.58 13.40 0 1
25%
ethanol 19.24 32.52 13.28 -0.12 1.09
β-actin TGF-β1 ΔCt ΔΔCt 2-ΔΔCt
Control 20.18 27.96 7.78 0 1
25%
ethanol 19.24 25.72 6.48 -1.30 2.46
β-actin c-Myc ΔCt ΔΔCt 2-ΔΔCt
Control 20.18 29.88 9.70 0 1
25%
ethanol 19.24 25.59 6.35 -3.35 10.17
β-actin Thymosin-β4 ΔCt ΔΔCt 2-ΔΔCt
Control 20.18 23.60 3.42 0 1
ethanol25% 19.24 21.36 2.12 -1.30 2.46
4. 고찰 및 결론
일반적으로 Thymosin-β4 유전자는 면역반응이나 혈 관 생성, 암전이 등에 관여 하며 c-Myc, Erb-B2 유전자 는 종양 관련 유전자이고 TGF-β1는 조직 섬유화 관련 유전자로 알려져 있다. 따라서 본 연구는 손상된 간조직 에서 TGF-β1, 및 c-Myc, Erb-B2, Thymosin-β4 발현 정도를 알아보고자 실시하였다.
결과에 대하여 살펴보면 알코올성 간 손상 조직에서 전자현미경적 조직변화는 정상 간 조직에서 미토콘드리 아 등 세포질 내 소기관은 정상적으로 분포하고 있었으 며 특이할 만할 소견을 찾아볼 수 없었지만 25% Ethanol
을 처리한 조직에서는 중심 정맥 주위에 용해소체들이 분포하고 있었다.
Ⓐ
Ⓑ
Fig. 1. Shapes of liver tissue in the Transmission Electron Microscope.
Ⓐ : Shapes of the control liver tissue in the Transmission Electron Microscope,
Ⓑ : Shapes of the liver tissue in the group administered with 25%
ethanol in the Transmission Electron Mi -croscope. This picture is not shows the structure of the mitochondria (→) and shows the structure of fat and lysosome.
동시에 미토콘드리아 상태는 속 사립체막의 내부로 돌출된 능선이 뚜렷하지 못하는 등 전체적으로 많은 손 상이 있었다. 혈청학적 검사는 ALT는 대조군에 비해 25% Ethanol 단독 투여 군에서 42.0 U/l로 높게 나타났으 며 AST는 대조군에 비해 25% Ethanol 단독 투여군에서 177.8 U/l로 높은 값을 보였다. 또한 ALP는 대조군에 비 해 25% Ethanol 단독 투여군 값이 97.4 U/l로 높다는 것 을 알 수 있었다. 각 평균값에 대한 t 검정은 P < 0.05로 유의성이 있다는 것을 알 수 있었다. H & E염색 결과는 대조군에 비해 25% Ethanol 단독 투여군에서 중심정맥 주위에 조직변성에 의한 핵농축과 저 염색성을 일으켰으 며 핵이 전체적으로 커지는 반응세포핵 등 이상 세포 등
을 볼 수 있었다. Real time PCR을 통한 발현 연구 시 Ct 값이 보통 18~32정도에서 이루어지는데 Erb-B2 유전 자는 대체로 이 값을 벗어났으므로 대조군이나 25%
Ethanol 투여군에서 유전자 발현이 많지 않은 것을 알 수 있었다.
Ⓐ
Ⓑ
Fig. 2. H & E stain of liver tissue.
Ⓐ : A is a H & E stain of control liver tissue,
Ⓑ : B is a H & E stain f liver tissue adm- inistered with 25%
ethanol. It is shown Pycnosis (↓), size change(←) of Nuclear and hypochromasia(→).
또한 Real time PCR 결과 분석방법인 케니스 방법 [42]을 이용하여 housekeeping gene인 β-actin 유전자 대조군 발현정도를 1 기준으로 하였을때 25% Ethanol 투 여군 발현양은 TGF-β1가 2.46, c-Myc 10.17, Thymosin-β42.46으로 유전자 발현이 증가 된 것을 알수 있었다. 위 결과를 보면 ALT, AST, ALP의 혈청학적 관 련 효소의 증가를 알 수 있었는데 이는 알코올에 의한 간 조직 손상을 의미하는 것으로 알코올에 의한 간의 독성 효과를 의미 한다 [43]는 논문과 알코올 투여시 혈청관련 효소 증가를 보고한 [44]연구 결과와 일치함을 알 수 있 었다. 즉 세포막에서 불포화지방산의 과산화반응으로 인 하여 막 구조나 기능을 파괴함으로써 간의 단백질 합성 을 저해하며 간의 글리코겐(glycogen)양을 감소시키고,
혈중으로 ALT 와 AST 등의 혈청 관련효소 유출의 증가 를 나타낸 것이다 [45]. 따라서 간 손상 영향 정도를 보여 주는 각종 혈청학적, 조직학적 결과와 간 조직 손상에 따 른 유전자 발현 정도를 분석하여 본 바 손상 받은 간 조 직에서는 TGF-β1, c-Myc 및 Thymosin-β4유전자의 발 현 증가를 알 수 있었으며 Erb-B2 유전자는 뚜렷한 발 현을 알 수 없었다. 결론적으로 기존 임상에서 간 기능 관련 질병 예방과 질환 판정 시 혈청학적, 조직학적 검사 외에 TGF-β1, c-Myc 및 Thymosin-β4의 분자 진단 기 법에 의한 유전자 발현 상태를 검사함으로써 간 질환 판 정의 보조 자료로 활용 될 수 있을 것으로 사료 된다
REFERENCES
[1] V. A. Ramchandani, W. F. Bosron, T. K. Li. (2001).
Research advances in ethanol metabolism. Pathol. Biol, 49, 676-682. .
[2] H. Rouach, M. Clement, M. T. Ofanelli, B. Janvier, J.
Nordmann, R. Nordmann. (1983). Hepatic lipid peroxidation and mitochondrial susceptibility to peroxidative attacks during ethanol inhalation and withdrawal. Biochem. Biophys, Acta, 753, 439-444.
[3] C. S. Lieber. (1994). Alcohol and liver :update.
Gastroenterology, 106, 1085-1090.
[4] S. Zakhari. (2006). Overview: how is alcohol metabolized by the body? Alcohol Res & Health, 29, 245-254.
[5] C. S. Lieber. (1970). New pathway of ethanol metabolism in the liver. Gastroenterology, 59, 930-937.
[6] H. L. Bleich, E. S. Boro. (1977). Metabolic and hepatic effects of alcohol. N Engl J Med, 296, 612-616.
[7] A. Gramenzi, F. Caputo, M. Biselli, F. Kuria, E. Loggi, P. Andreone, M. Bernardi. (2006). Review article:
alcoholic liver disease-pathophysiological aspects and risk factors. Aliment Pharmacol Ther, 24, 1151-1161.
[8] S. K. Das, D. M. Vasudevan. (2007). Alcohol-induced oxidative stress. Life Sci, 81, 177-187.
[9] K. Nagata, H. Suzuki, S. Sakaguchi. (2007). Common pathogenic mechanism in development progression of liver injury caused by non-alcoholic or alcoholic steatohepatitis. J Toxicol Sci, 32, 453-468.
[10] C. S. Lieber. (2004). Milestones in liver disease. J Hepatol, 40, 198-202.
[11] T. Byers, G. Perry. (1992). Dietary carotenes, vitamin C, and vitamin E as protective antioxidants in human cancers. Ann Rev Nutr, 12, 135-159.
[12] R. A. Lawrence, R. F. Burk. (1976). Glutathione peroxidase activity in selenium-deficient rat liver.
Biochem Biophys Res Commun, 71, 952- 958.
[13] T. L. Low, A. L. Goldstein. (1982). Chemical characterization of thymosin beta-4. J. Biol. Chem, 257, 1000-1006.
[14] M. J. Kim, (2017). A study on Expression of Thymosin β4in Cervical Cancer induced Human Papilloma Virus, konyang university Graduate school. 7-8.
[15] K. C. Hong, (2015). Overexpression and characterization of thymosin beta-4 in Escherichia coli, Dongwee univrsity Graduate school, 10-11.
[16] K. O. Hong, (2016). Thymosin b4 induces proliferation, invasion, and epithelial-to-mesenchymal transition of oral squamous cell carcinoma, seoul univrsity Graduate school, 1-2.
[17] S. H. Yoon. (2008.). Expression of Thymosin-β4 in odontoblast differentiation of the mouse. Josun Univ.
Graduate school.
[18] D. Safer, M. Elzinga, V. T. Nachmias. (1991).
Thymosin beta4 and Fx, an actin-sequestering peptide, are indistinguishable. J. Biol. Chem, 266, 4029-4032.
[19] D. Safer, R. Golla, V. T. Nachmias. (1990). Isolation of a 5-kilodalton actin-sequestering peptide from human blood platelets. Proc. Natl. Acad. Sci. USA, 87, 2536-2540.
[20] D. Pantaloni, M. F. Carlier. (1993). How profiling promotes actin filament assembly in the presence of thymosin b4. Cell, 75, 1007-1014.
[21] F. Kang, D. L. Purich, F. S. Southwick. (1999). Profilin promotes barbed- end actin filament assembly without lowering the critical concentration. J Biol Chem, 274, 36963–36972.
[22] D. S. Grant, J. L. Kinsella, M. C. Kibbey, S. LaFlamme, P. D. Burbelo, A. L. Goldstein, H. K. andKleinman.
(1995). Matrigelinducesthymosin beta4 gene in differentiating endothelial cells. J Cell Sci, 108, 3685-3694.
[23] D. S. Grant, W. Rose, C. Yaen, A. Goldstein, J.
Martinez, H. Kleinman. (1999). Thymosin-β4 enhances endothelial cell differentiation and angio genesis. Angio genesis, 3, 125–135.
[24] T. Huff, C. S. Muller, A. M. Otto, R. Netzker, E.
Hannappel. (2001). beta-Thymosins, small acidic peptides with multiple functions. Int J Biochem Cell Biol, 33, 205-209.
[25] J. Go´mez-Ma´rquez, J. I. Pedrares, A. Otero, R.
Anado´n. (1993). Prominent expression of the
actin-sequestering peptide Fx gene in the hippo campal region of ratbrain. Neuroscience Letters, 152, 41–44.
[26] J. Go´mez-Ma´rquez, F. F. delAmo, P. Carpintero, R.
Anado´n. (1996). High levels of mouse thymosin b4 mRNA in differentiating P19 embryonic cells and during development of cardiovascular tissues. Biochim BiophysActa, 1306, 187–193.
[27] R. Anadon, I. R. Moldes, P. Carpintero, G. Evangelatos, E. Livianou, L. Leondiadis, I. Quintela, M. C. Cervino, J. Go´mez-Ma´rquez. (2001). Differential expression of thymosin b4 and b10 during rat cerebellum postnatal development. Brain Res, 894, 255-265.
[28] D. Philp, A. L. Goldstein, H. K. Kleinman. (2004).
Thymosin β4 promotes angiogenesis, wound healing, and hair follicle development. Mech Ageing Dev, 125, 113-115.
[29] W. S. Wang, P. M. Chen, H. L. Hsiao, S. Y. Ju, Y.
Su. (2003). Over expression of the thymosin beta-4 gene is associated with malignant progression of SW480 colon cancer cells. Oncogene, 22, 3297-3306.
[30] K. M. Malinda, A. L. Goldstein, H. K. Kleinmman.
(1997). Thymosin beta 4 stimulates directional migration of human umbilical vein endothelial cells. FASEG J, 11, 474-481.
[31] E. A. Clark, T. R. Golub, E. S. Lander, R. O. Hynes.
(2000). Genomics analysis of metastasis reveals an essectial role for RhoC. Nature, 406, 532-535.
[32] T. Kobayashi, F. Okada, N. Fujii, N. Tomita, S. Ito, H. Tazawa. (2002). Thymosin-beta 4 regulates motility and metastasis of malignant mouse fibrosarcoma cells.
Am J Pathol, 160, 869-82.
[33] K. H. Kim, K. J. Jo, Y. Go. (2011). Understanding and technique of genetics, Korea medical book, 203-205.
[34] M. Matsuoka, H. Tsukamoto. (1990). stimulation of hepatic lipocytie collagen pruduction by Kupffer cell-derived transforming growth factor B: implication for a pathogentic role in liver fibrogenesis. Hepatology, 11, 599-605.
[35] H. Nakatsukase, R. P. Evarts, C. C. Hsia, S, S.
Thorgeirsson. (1990). Transfoming growth factor-B1 and type I procollagen transscripts during regeneration and early fibrosis of rat liver. Lab Invest, 63, 171-180.
[36] D. A. Brenner. (1991). Transforming growth factor B and hepatic fibrosis: cause of effect?, Hepatology, 14, 740-742.
[37] A. Castilla, J. Prieto, N. Fausto. (1991). Transforming growth factor B1 and a in chronic liver disease. N Engl J Med, 324, 933-940.
[38] Z. Qi, N. Atsuchi, A. Ooshima, A. Takeshita, H. Ueno.
(1999). Blockade of type B transforming growth factor sigaling prevents liver fibrosis and dysfunction in the rat, Proc Natl Acad Sci, 96, 2345-2349.
[39] Y. S. Choi, (2017), The Regulatory Role of Dickkopf-1 as a Target of Oncogenic Transforming Growth Factor-β 1-Smad3 Pathway in Human Lung Cancer Cells, gangwon univrsity Graduate school, 1-2.
[40] J. H. Park, (2016), The association of single nucleotide polymorphisms in the TGF-β1 and RAGE genes with polycystic ovary syndrome, Cha university General Graduate Shool, 2-3.
[41] J. H. Lee , C. G, Yoon, S. I. Lee. (1993). Effects of dietary protein on the changes of lipoprotein fractions in carbon tetrachloride-treated rats. J Korean Soc Food Sci Nutr, 22, 127-131.
[42] J. L. Kenneth, D. S. Thomas. (2001). Analysis of relative gene expression data using real-time Quantitative PCR and the 2-DDCT method. Methods, 25, 402-408.
[43] Y. H. You, K. Y. Jung, Y. H. Lee, W. J. Jun, B. Y.
Lee. (2009). Hepatoprotective effects of hovenia dulcis fruit on ethanol-induced liver damage in vitro and in vivo. J Korean Soc Food Sci Nutr, 38, 154-159.
[44] Y. T. Ahn, J. S. Bae, Y. H. Kim, K. S. Lim, C. S.
Huh. (2005). Effects of fermented milk intake on hepatic antioxidative systems in alcohol treated rats. Korean J Food Sci Technol, 37, 631-635.
[45] P. Sher, (1997). Diagnostic effectiveness of biochemical liver-function tests, as evaluated by discriminant function analysis. Clin Chem, 23, 627- 630.
김 진 수(Jean-Soo kim) [정회원]
▪1997년 7월 ∼ 2001년 3월 : 삼성 서울병원
▪2001년 4월 ∼ 2012년 3월 : 건양 대학교병원
▪2012년 3월 ∼ 2014년 2월 : 광양 보건대학교 교수
▪2014년 3월 ∼ 현재 : 대전보건대학교 교수
▪관심분야 : 생물학, 조직학
▪E-Mail : [email protected]
최 상 기(Sang ki, Choi) [정회원]
▪1983년 2월 :서울대학교(학사)
▪1985년 2월 :서울대학교(석사)
▪1995년 2월 : 미국 Georgia 주립 대(박사)
▪2003년 2월 ∼ 현재 : 국립순천대 학교 교수
▪관심분야 : 생물학, 기능 유전체학
▪E-Mail : [email protected]