Effects of Pomace of Schizandra chinensis, Schizandrin, and Gomisin A on LPS-induced Inflammatory Responses in RAW264.7 Cells
Yu-Mi Seo
1, Hyun-Ji Kim
1, Eun-Joo Lee
1, Chungwook Chung
1, Hwa-Jung Sung
2, Ho-Yong Sohn
2, Jong-Yi Park
3and Jong-Sik Kim
1*
1Department of Biological Sciences, Andong National University, Andong 36729, Korea
2Department of Food and Nutrition, Andong National University, Andong 36729, Korea
3Gyeongbuk Institute for Bio Industry, Andong 36728, Korea
Received October 16, 2017 /Revised November 30, 2017 /Accepted December 4, 2017
Schizandra chinensis has been used as a traditional Chinese medicine and is known to have various bioactive components, including schizandrin and gomisin A. In the current study, we investigated the anti-inflammatory activities and their working mechanisms of ethanol extracts of pomace of Schizandra chinensis (PSC), schizandrin (SZ), and gomisin A (GA). First, we analyzed the effects of PSC on nitric oxide (NO) production and cell viabilities in lipopolysaccharide (LPS)-stimulated RAW264.7 cells. The results indicated that PSC dramatically reduced NO production in LPS-activated RAW264.7 cells in a dose-dependent manner without affecting cell viabilities. PSC also decreased the expression of pro-inflammatory genes iNOS and COX-2, whereas the expression of TNF-α was not affected by PSC.
In addition, PSC inhibited phosphorylation of p38, ERK1/2, and JNK but did not change the ex- pression of their total protein. The results indicate that PSC can regulate LPS-induced inflammatory responses by suppressing MAPK (mitogen-activated protein kinase) signaling. We also analyzed the effects of SZ and GA on NO production and cell viabilities in RAW264.7 cells. The results showed that SZ and GA also decreased NO production in a dose-dependent manner in LPS-activated RAW 264.7 cells without affecting cell viabilities. SZ reduced the expression of iNOS, whereas GA down- regulated iNOS and COX-2. Overall, these findings clarify the molecular mechanisms of the anti-in- flammatory effects mediated by PSC, SZ, and GA.
Key words : Anti-inflammation, Gomisin A, MAPK, Schizandra chinesis, Schizandrin
*Corresponding author
*Tel : +82-54-820-5798, Fax : +82-54-820-7705
*E-mail : [email protected]
This is an Open-Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Life Science 2018 Vol. 28. No. 3. 339~344 DOI : https://doi.org/10.5352/JLS.2018.28.3.339
서 론
오미자(Schizandra chinensis Baillon)는 목련과의 넝굴성 식 물인 오미자나무의 열매로, 단맛, 짠맛, 신맛, 쓴맛, 매운맛의 5가지 맛이 나며 약학적 활성이 뛰어나서 약용식물로 이용되 고 있다[6]. 2007년 세계보건기구(World Health Organization, WHO)의 보고에 의하면, 오미자의 주요 구성성분은 di- bennzo[a,c]cyclooctadiene 골격을 가진 약 30 종류의 리그난 이며, 대표적인 생리활성물질로는 schizandrin과 gomisin A 및 deoxyschizandrin 등이 있다[20]. 최근 오미자 및 오미자 추출물의 다양한 약리작용에 대한 연구가 활발히 진행되고 있으며, 항산화, 항균 및 항혈전 활성[6], 이상지질혈증과 항비 만 효과[5, 14], 간 보호 효과[1], 항염증 활성[4, 13] 그리고 항암
활성[11]에 대한 연구가 보고된 바 있다.
최근 식품의 품질 향상을 위해 오미자 및 오미자 추출액을 활용한 다양한 가공식품이 개발되고 있다[8, 15]. 하지만 활용 도가 높은 과즙에 비해 착즙 후 다량으로 생기는 가공 부산물 인 오미자 박(찌꺼기, pomace)은 별도의 활용 없이 대부분 폐 기되고 있는 실정이다. 오미자 박 에탄올 추출물의 항산화, 항균 및 혈액응고저해 활성이 오미자 씨와 열매보다 더 우수 하다고 보고된 바 있다[6]. 그러나, 아직까지 오미자 박 추출물 에 의한 항염증 활성과 작용기전 연구는 매우 미미한 수준이 다. 따라서, 본 연구에서는 오미자 박 추출물과 함께 오미자 유래의 순수물질인 schizandrin, gomisin A에 의한 항염증 활 성과 작용기전을 연구하고자 하였다.
염증이란 lipopolysaccharide (LPS)와 같은 다양한 외부 자 극에 대한 방어 기작으로[17], 면역을 담당하는 대식세포는 염 증을 매개하는 nitric oxide 및 다양한 pro-inflammatory cyto- kine의 방출을 통해 염증에 관여한다. 염증 반응이 심화되어 만성 염증으로 발전할 경우, 암의 발생과도 연관이 있다. 만성 염증인 염증성 장질환이 있는 경우 대장암으로 진행될 확률이 정상인에 비해 4~20 배 이상 높다고 보고된 바 있으며, 현재 염증반응 조절을 위해 단계별 간섭에 대한 연구가 진행되고
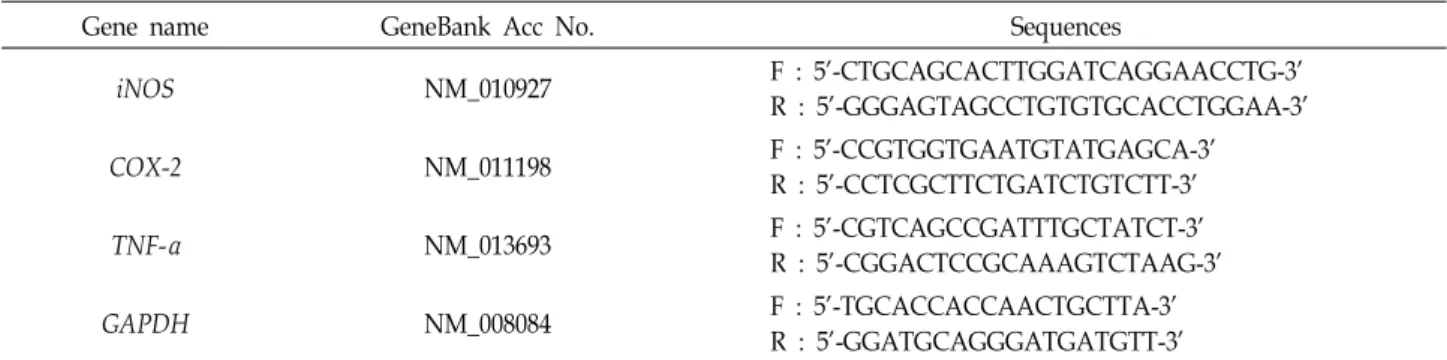
Table 1. Sequences of oligonucleotide primers used for reverse-transcription PCR
Gene name GeneBank Acc No. Sequences
iNOS NM_010927 F : 5'-CTGCAGCACTTGGATCAGGAACCTG-3'
R : 5'-GGGAGTAGCCTGTGTGCACCTGGAA-3'
COX-2 NM_011198 F : 5'-CCGTGGTGAATGTATGAGCA-3'
R : 5'-CCTCGCTTCTGATCTGTCTT-3'
TNF-α NM_013693 F : 5'-CGTCAGCCGATTTGCTATCT-3'
R : 5'-CGGACTCCGCAAAGTCTAAG-3'
GAPDH NM_008084 F : 5'-TGCACCACCAACTGCTTA-3'
R : 5'-GGATGCAGGGATGATGTT-3' 있다[18, 21].
세포막에 있는 톨-유사 수용체4(Toll-like receptor 4, TLR4) 와 MD-2 복합체가 LPS를 인식하면 세포 내 신호전달경로인 mitogen-activated protein kinase (MAPK)가 활성화되며, MAPK family에는 p38, extracellular signal-regulated kinase 1/2 (ERK1/2) 및 c-jun-N-terminal kinase (JNK)의 세 가지 단백질이 있다[12]. MAPK 경로의 활성화는 nuclear factor-κB (NF-κB)와 같은 전사조절인자의 발현 조절을 통해 NO와 다양 한 염증 매개 cytokine의 생성을 촉진한다. 이 과정에서 과다 생산된 NO와 cytokine은 심각한 조직 손상 및 패혈성 쇼크를 유발할 수 있어 염증 반응을 조절하는 것이 중요하다[2]. 오미 자의 유효 생리활성 물질인 schizandrin과 gomisin A가 NF-κ B와 MAPK 경로의 조절을 통해 항염증 효과를 나타낸다는 보고[3, 19]가 있으나, 이들이 포함된 오미자 박에 대한 LPS 유도 염증 모델에서의 연구는 보고된 바가 없다.
따라서, 본 연구에서는 오미자 박 에탄올 추출물과 오미자 유래 생리활성물질인 schizandrin 및 gomisin A에 의한 LPS 로 염증 유도된 마우스 대식세포 RAW264.7 세포에서 항염증 활성 및 그 작용기전에 대해 연구하였다.
재료 및 방법
오미자 박 에탄올 추출물의 제조 및 시약
본 실험의 오미자 박은 2016년 문경 농가에서 수확한 오미 자를 오미자 가공 공장에서 착즙기를 이용하여 착즙한 직후의 박(수분함량 60.5±3.3%)으로, 4℃의 냉장조건에서 실험실로 옮겨 사용하였다. 오미자 박 추출물 조제를 위해 오미자 박 15 kg에 20배의 95% ethanol을 가하여 상온에서 24시간 추출 하였으며, 상기 추출을 3회 반복하였다. 이후 추출액은 filter paper (Whatman No. 2)로 거른 후 감압 농축(Eyela Rotary evaporator N-1000, Tokyo Rikakikai Co., Ltd. Japan)하여 분 말로 조제하였다. 조제된 시료는 dimethyl sulfoxide (DMSO) 에 녹여 실험에 사용하였다. 순수물질인 schizandrin과 gomisin A는 Sigma사(St. Louis, MO, USA)에서 구입하여 사용하였다.
마우스 대식세포 RAW264.7 세포주 배양
마우스 대식세포 RAW264.7 세포주는 American Type Culture Collection (ATCC, Fredrick, MD, USA)에서 구입하 였다. 세포주 배양에는 10% Fetal Bovine Serum (FBS, Gibco, Grand Island, NY, USA), 1% penicillin 및 streptomycin (WelGene, Korea)이 첨가된 Dulbecco’s Modified Eagle Medium (DMEM, Gibco)을 사용하였다. 배양은 37℃, 5% CO2
조건의 배양기에서 실시하였다.
Nitric oxide (NO) 생성 분석
오미자 박 에탄올 추출물, schizandrin 및 gomisin A가 LPS 로 유도된 대식세포 RAW264.7 세포주의 nitric oxide 생성에 미치는 영향을 측정하기 위하여 nitric oxide 생성 분석을 실시 하였다. RAW264.7 세포주를 96 well plate의 각 well에 2×105 개 접종한 후 18시간동안 배양하였다. 그 후 LPS (Sigma)를 0.2 μg/ml의 농도로 1시간 동안 처리하고, 각 시료를 농도별로 처리하여 16시간 동안 배양하였다. 그 후 상등액 100 μl와 Griess reagent (Sigma) 100 μl를 혼합하여 15분 동안 상온에서 반응시키고 NanoQuant Plate™ (Tecan trading AG, Switzer- land)를 사용하여 540 nm에서 흡광도를 측정하였다. NO 생성 분석은 독립적인 5개의 well에서 수행되었으며, 통계 분석은 Sigma plot을 이용하여 mean ± SD 값으로 나타내었다.
세포생존율 분석
오미자 박 에탄올 추출물, schizandrin 및 gomisin A에 의한 세포독성을 측정하기 위해 CellTiter 96® AQueous One Solution Cell Proliferation Assay (MTS) kit를 이용하였다. 96 well에 2x105 개의 RAW264.7 세포를 접종하여 각 시료를 농도 별로 처리한 후 16시간 동안 반응시켰으며, 세포 배양액 100 μl 에 MTS 용액 20 μl를 첨가하였다. 그 후 37℃, 5% CO2 배양기에 4시간동안 반응시키고 NanoQuant Plate™ (Tecan trading AG, Switzerland)를 사용하여 580 nm에서 흡광도를 측정하였다. 세 포생존율 분석은 독립적인 5개의 well에서 수행하였다.
Reverse-transcription polymerase chain reaction Total RNA 추출은 RNeasy mini kit (Qiagen, Valencia, CA,
A B
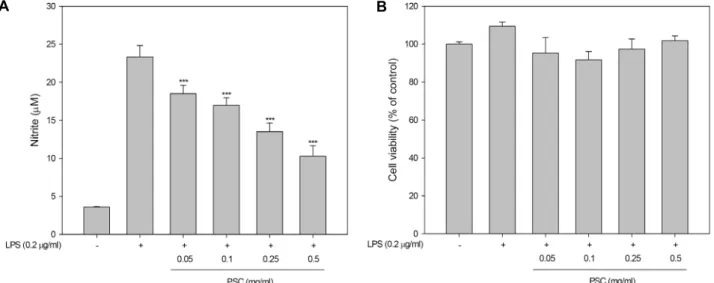
Fig. 1. Effect of ethanol extracts of pomace of Schizandra chinensis (PSC) on nitric oxide production and cell viabilities in LPS-stimulated RAW264.7 cells. RAW264.7 cells were 2x105 cells/well in a 96 well plate and incubated with PSC for 16 hr. And then, (A) nitric oxide production was measured by nitric oxide production assay and (B) cell viability was measured using MTS pro- liferation assay kit. Value indicate means ± SD (n=4). *p<0.05, **p<0.01, ***p<0.001.
USA)를 사용하였으며 제조사의 매뉴얼을 따라 수행하였다.
Total RNA는 NanoQuant Plate™를 이용하여 정량 한 뒤 -80℃
에 보관하며 사용하였다. RAW264.7 세포주로부터 추출한 to- tal RNA 2 μg을 주형으로 PrimeScript™ RT-PCR kit (TaKaRa, Japan)를 이용하여 제조사의 매뉴얼에 따라 cDNA를 합성하 였다. 합성된 cDNA에 RNase free water 40 μl를 첨가한 후 -20℃에 보관하며 사용하였다. PCR은 합성된 cDNA를 주형으 로 하였으며 유전자 특이적인 oligo primer를 이용하였다.
PCR에 사용된 primer는 Table 1과 같고, internal control 유전 자로 GAPDH를 사용하였으며, primer는 Bioneer 사(Korea)와 Macrogen 사(Korea)에서 구입하였다. PCR은 TaKaRa Ex Taq (TaKaRa)을 이용하여 수행하였으며, PCR 반응은 95℃에서 5 분간 denaturation하고 94℃에서 30초, 58℃에서 30초, 72℃에 서 30초의 cycle을 25회 반복한 후, 마지막으로 72℃에서 10분 간 extension하였다. PCR product는 1.5% agarose gel에서 전 기영동 하고 ethidium bromide (EtBr, Bioneer, Korea)로 염색 하여 gel image analysis system (Corebio, Korea)을 이용하여 사진 촬영하였다.
Western blot analysis
세포 접종 18시간 후 오미자 박 추출물, schizandrin 및 go- misin A를 serum free media에 혼합하여 4시간 동안 처리한 후, LPS를 1.0 μg/ml의 농도로 15분 동안 처리하였다. 처리 후 얼음을 이용하여 세포활동을 정지시킨 뒤 10X RIPA buffer (Cell signaling, Beverly, MA, USA)를 4X로 만들어 첨가한 후 세포를 수확하여 sonication 하였다. Sonication 후 cell ly- sate를 3,000 rpm에서 15분 동안 원심분리 한 뒤 상층액을 분 리하여 -20℃에 보관하며 실험에 사용하였다. 단백질의 정량
은 Bradford assay (Bio-Rad, Hercules, CA, USA) 방법을 이용 하였다. 본 실험에서는 p38, ERK1/2, JNK, p-p38, p-ERK1/2 및 p-JNK, ACTIN 1차 항체를 사용하였으며, 2차 항체는 HRP- conjugated rabbit antibody 및 HRP-conjugated mouse antibody를 사용하였다. ACTIN과 2차 항체는 Santa Cruz 사 (Santa Cruz, CA, USA)로부터 구입하였고, 그 외의 1차 항체 는 Cell signaling사로 부터 구입하여 사용하였다.
통계 분석
모든 실험은 최소 3회 이상 실시하였으며, 실험결과는 평균
± 표준편차로 나타내었고, 각 실험결과의 유의성 검토는 시료 가 포함되지 않은 대조구와 비교하여 Student’s t-test에 의해 판정하였으며 p 값이 0.05 미만일 때 유의성이 있다고 판단하 였다.
결과 및 고찰
오미자 박 에탄올 추출물 처리에 따른 NO 생성 및 세포생 존율 분석
오미자 박 에탄올 추출물(PSC, Pomace of Schizandra chi- nensis)이 LPS로 활성화된 RAW264.7 세포의 nitric oxide (NO) 생산과 세포생존에 미치는 영향을 연구하였다. Nitric oxide (NO)는 안정하고 독성을 가진 자유 라디칼 기체성분으로 대 식세포로 매개되는 숙주의 방어기작에 관여하는 염증반응의 주요지표로 여겨진다[9]. LPS로 활성화된 RAW264.7 세포주 에 0.05, 0.1, 0.25, 0.5 mg/ml의 PSC 시료를 각각 처리한 결과, PSC에 의해 농도의존적으로 NO의 생성이 감소하는 것을 확 인하였다(Fig. 1A). 그리고 세포생존율 측정 결과 PSC가 세포
Fig. 2. Down-regulation of expression of iNOS and COX-2 gene by PSC treatment. Mouse macrophage RAW264.7 cells were treated with 0.25 and 0.5 mg/ml PSC and total RNA was prepared. And then, reverse-transcription PCR was performed with iNOS, COX-2 and TNF-α gene spe- cific primers.
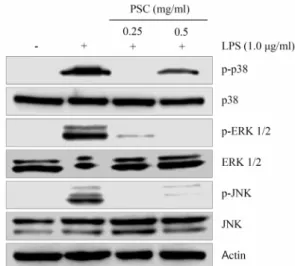
Fig. 3. Inhibition of phosphorylation of p-38, ERK1/2, and JNK by PSC. Mouse RAW264.7 cells were treated with 0.25 and 0.5 mg/ml PSC for 4 hr. And then cell lysates were prepared after LPS (1.0 μg/ml) treatment for 15 min.
Western blot analysis was carried out by using p38, phospho-p38, ERK1/2, phospho-ERK1/2 JNK, phos- pho-JNK, and ACTIN antibodies.
생존율에는 큰 영향을 주지 않은 것으로 측정되었다(Fig. 1B).
따라서, PSC는 LPS로 자극된 RAW264.7 세포에서 세포 생존 율에는 영향을 미치지 않으면서 NO의 생성을 처리한 PSC 농 도의존적으로 저해함을 확인하였다. 추가 연구는 PSC의 처리 농도를 0.25와 0.5 mg/ml 두 가지 농도로 선정하여 진행하였 다.
오미자 박 에탄올 추출물에 의한 염증 유전자 발현 분석 그람 음성세균의 세포벽 성분인 LPS와 같은 세포 외부 자극 은 iNOS (inducible nitric oxide synthase) 유전자의 발현을 유도하며, 대식세포가 활성화되면 iNOS가 높은 수준으로 발 현된다[7]. PSC에 의한 NO 생성 억제 기전을 연구하기 위하여 pro-inflammatory 유전자인 iNOS, COX-2 및 TNF-alpha의 유 전자 발현 분석을 실시하였다. LPS로 활성화된 RAW264.7 세 포주에 0.25 및 0.5 mg/ml의 PSC를 처리한 후 염증 유전자 발현을 분석하였다. 그 결과, iNOS와 COX-2 유전자의 발현이 PSC에 의해 농도의존적으로 감소하였지만 TNF-alpha의 발현 변화는 관찰되지 않았다(Fig. 2). 이러한 결과는 PSC가 iNOS 와 COX-2와 같은 pro-inflammatory 유전자의 발현을 억제함 으로써 항염증 활성을 나타냄을 시사한다.
오미자 박 에탄올 추출물에 의한 MAPK 신호전달 감소 MAPK는 MAPK kinase kinase (MAPKKK)와 MAPK kin- ase (MAPKK)의 인산화를 통해 활성화되는 세포 내 신호전달 경로이며, 전사조절인자 활성화 및 유전자 발현 촉진을 통해 cytokine 생산 및 세포 이주 등과 같은 다양한 세포 반응을 조절하는 것으로 알려져 있다[18]. PSC가 MAPK 경로를 통해 염증 반응을 조절하는지 확인하기 위해 MAPK 발현 분석을 수행하였다. PSC를 0.25와 0.5 mg/ml의 농도로 RAW264.7에 4시간 동안 처리한 후 LPS로 15 분 동안 자극하여 p38, ERK1/
2와 JNK 인산화의 변화를 측정하였다. Western blot 분석 결
과 0.25 mg/ml 농도의 PSC 처리군에서 p38, ERK 및 JNK의 총 단백질의 발현 변화 없이 인산화가 모두 억제되었으며, 0.5 mg/ml 처리군에서도 MAPK 인산화 수준이 감소된 것을 확 인하였다(Fig. 3). 이러한 결과를 통해 PSC가 MAPK 신호전달 경로를 억제함으로써, LPS로 유도된 염증반응을 억제한다는 것을 확인하였다.
Schizandrin 및 gomisin A 처리에 따른 NO 생성 및 세 포생존율 분석
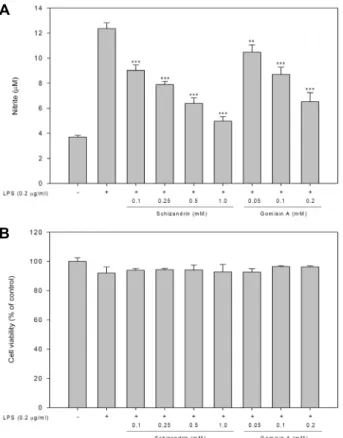
최근 오미자에서 유래된 schizandrin (SZ)과 gomisin A (GA)를 포함한 다양한 생리활성 성분과 약리학적 특성에 대한 총평이 보고되었다[16]. 본 연구에서는 오미자 유래의 생리활 성물질인 SZ과 GA가 LPS로 활성화된 RAW264.7 세포주에서 NO 생산 및 세포생존율에 미치는 영향을 확인하였다. Fig.
4에서 보는 바와 같이 SZ과 GA가 RAW264.7의 세포생존율에 영향을 미치지 않고 농도의존적으로 NO의 생성을 감소시킨 것을 확인하였다. 이 결과에 따라, 추가 연구를 위해 SZ은 0.5 mM로 GA는 0.2 mM로 처리농도를 선정하였다.
Schizandrin 및 gomisin A에 의한 염증 유전자 발현 감소 오미자 유래 순수물질인 SZ과 GA에 의한 항염증 활성 기전 을 이해하기 위하여 염증유발유전자인 iNOS, COX-2 및 TNF-alpha의 유전자 발현 분석을 실시하였다. LPS로 염증이 유도된 RAW264.7 세포주에 0.5 mM의 SZ과 0.2 mM의 GA를 처리한 후 RT-PCR 분석을 실시하였다. 그 결과, SZ를 처리한 경우는 iNOS 유전자의 발현만 감소된 반면, GA를 처리한 경
A
B
Fig. 4. Effect of schizandrin (SZ) and gomisin A (GA) on nitric oxide production and cell viabilities in LPS-activated RAW264.7 cells. RAW264.7 cells were 2x105 cells/well in a 96 well plate and incubated with SZ and GA for 16 hr. After treatment, (A) Nitric oxide production was measured by nitic oxide production assay and (B) cell viability was measured using MTS proliferation assay kit. Value indicate means ± SD (n=4). *p<0.05, **p<0.01,
***p<0.001.
Fig. 5. Down-regulation of iNOS and COX2 genes by SZ and GA. RAW264.7 cells were treated with 0.5 mM SZ or 0.2 mM GA and total RNA was prepared. And then, reverse-transcription PCR was performed with iNOS, COX-2 and TNF-α gene specific primers.
우는 iNOS와 COX-2 유전자 모두 발현이 감소됨을 확인하였 다(Fig. 5). 이러한 결과는 오미자 유래의 두 순수물질인 schi- zandrin과 gomisin A의 항염증 활성 작용기전이 다를 수 있음
을 시사한다. 하지만, 정확한 증명을 위해서는 MAPK path- way를 포함한 여러 pathway의 inhibitor를 이용한 실험이 요 구된다. 한편, NO 생산을 담당하는 iNOS와 더불어 cyclo- oxygenase-2 (COX-2) 또한 염증반응의 매개자로 작용하며, COX-2는 prostaglandin E₂(PGE)를 생성하여 염증반응을 악 화시킨다고 알려져 있다[10].
종합적으로, 오미자 박 에탄올 추출물(PSC), 오미자 유래 순수물질인 schizandrin (SZ)과 gomisin A (GA)는 LPS로 염 증이 유도된 RAW264.7 세포에서 세포 생존율에 영향을 주지 않으면서 다양한 기전을 통해 항염증 활성을 나타냄을 확인하 였다. 이러한 결과는 오미자에 의한 항염증 활성 및 작용기전 을 이해하는데 도움을 줄 수 있을 것으로 생각된다.
감사의 글
본 연구는 2016년도 산업통상자원부 바이오테라피산업기 반구축사업(과제번호 N0001805)에 의해 수행되었으며, 이에 감사드립니다.
References
1. Choo, B. K., Chung, K. H., Seo, Y. B. and Roh, S. S. 2013.
Antioxidant, antiinflammation and hepatoprotective activity of Schizandrae Fructus processed with differensiated steam- ing number. Kor. J. Herbology 28, 83-92.
2. Ci, X., Ren, R., Xu, K., Li, H., Yu, Q., Song, Y., Wang, D., Li, R. and Deng, X. 2010. Schisantherin A exhibits anti-in- flammatory properties by down-regulating NF-kappaB and MAPK signaling pathways in lipopolysaccharide-treated RAW 264.7 cells. Inflammation 33, 126-136.
3. Guo, L. Y., Hung, T. M., Bae, K. H., Shin, E. M., Zhou, H.
Y., Hong, Y. N., Kang, S. S., Kim, H. P. and Kim, Y. S. 2008.
Anti-inflammatory effects of schisandrin isolated from the fruit of Schisandra chinensis Baill. Eur. J. Pharmacol. 591, 293- 299.
4. Jang, J. Y. and Park, G. H. 2015. Anti-inflammatory effect of seed oil of Schisandra chinensis in the LPS-treated RAW 264.7 macrophages. Kor. J. Herbology 30, 77-82.
5. Jeong, E. J., Lee, W. J. and Kim, K. Y. 2009. Effects of Schizandra chinensis extract on the growth of intestinal bac- teria related with obesity. Kor. J. Food Sci. Technol. 41, 673-680.
6. Kim, M. S., Sung, H. J., Park, J. Y and Sohn, H. Y. 2017.
Evaluation of anti-oxidant, anti-microbial and anti-thrombo- sis activities of fruit, seed and pomace of Schizandra chinensis Baillon. J. Life Sci. 27, 131-138.
7. Korhonen, R., Lahti, A., Kankaanranta, H. and Moilanen, E. 2005. Nitric oxide production and signaling in inflamma- tion. Curr. Drug Targets Inflamm. Allergy 4, 471-479.
8. Lee, J. H. 2011. Quality of Sikhe incorporated with hot water extract of Omija (Schisandra chinensis Baillon) fruit. Food Eng.
Prog. 15, 80-84.
초록:오미자 박, schizandrin 및 gomisin A에 의한 RAW264.7 세포주에서 lipopolysaccharide 로 유도된 염증 반응의 억제
서유미1․김현지1․이은주1․정정욱1․성화정2․손호용2․박종이3․김종식1*
(1국립안동대학교 생명과학과, 2국립안동대학교 식품영양학과, 3경북바이오산업 연구원)
오미자는 전통적인 한약재로서 schizandrin과 gomisin A와 같은 다양한 생리활성물질을 함유하고 있는 것으로 알려져 있다. 본 연구에서는 오미자 박의 에탄올 추출물(PSC)과 schizandrin (SZ) 및 gomisin A (GA)에 의한 항염 증 활성 및 그들의 작용기전 연구를 수행하였다. 먼저, PSC는 LPS에 의해 염증이 유도된 RAW264.7 세포에서 세 포생존율에는 영향을 미치지 않고 농도의존적으로 nitric oxide (NO) 생성을 감소시켰다. PSC는 염증유발유전자 인 iNOS와 COX-2의 발현을 감소시켰으나, TNF-α의 발현에는 영향을 주지 않았다. 또한 오미자 박의 에탄올 추출 물은 p38, ERK1/2 및 JNK의 총 단백질의 발현에는 영향을 주지 않으면서, 그들의 인산화를 감소시켰다. 이러한 결과는 PSC가 MAPK 신호를 저해함으로써 염증 반응을 조절할 수 있음을 시사한다. 또한 SZ와 GA도 LPS에 의 해 염증이 유도된 RAW264.7 세포에서 세포 생존율에 영향을 미치지 않으면서 NO 생성을 감소시켰다. SZ은 iNOS 유전자의 발현만을 감소시킨 반면, GA는 iNOS와 COX-2 두 유전자의 발현을 모두 감소시켰다. 종합적으로 이러한 연구결과는 오미자 박 추출물, schizandrin 및 gomisin A에 의해 중재되는 항염증 활성 및 작용기전을 이해하는데 도움을 줄 것이다.
9. Maeng, O., Kim, Y. C., Kim, Y. S., Paik, S. G. and Lee, H.
Y. 2002. The function and regulation of NO; Identification of a gene that is up-regulated in RAW264.7 macrophage cells by nitric oxide. Bulletin of Biotechnology Chungnam National University 8, 1-9.
10. Morita, I. 2002. Distinct functions of COX-1 and COX-2.
Prostaglandins Other Lipid Mediat. 68-69, 165-175.
11. Park, C., Hwang, H. J., Choi, B. T., Choi, T. H., Kim, B.
W., Choi, Y. W. and Choi, Y. H. 2010. G1 arrest of the cell cycle by gomisin N, a dibenzocyclooctadiene lignan, iso- lated from Schizandra chinensis Baill in human leukemia U937 cells. J. Life Sci. 20, 977-982.
12. Park, S., Shin, H. J., Shah, M., Cho, H. Y., Anwar, M. A., Achek, A., Kwon, H. K., Lee, B., Yoo, T. H. and Choi, S.
2017. TLR4/MD2 specific peptides stalled in vivo LPS-in- duced immune exacerbation. Biomaterials 126, 49-60.
13. Park, S. Y., Park, D. J., Kim, Y. H., Kim, Y., Kim, S. G., Shon, K. J., Choi, Y. W. and Lee, S. J. 2011. Upregulation of heme oxygenase-1 via PI3K/Akt and Nrf-2 signaling pathways mediates the anti-inflammatory activity of Schis- andrin in Porphyromonas gingivalis LPS-stimulated macro- phages. Immunol. Lett. 139, 93-101.
14. Shin, Y. R., Kim, Y. K. and Kim, K. M. The Effects of Schizandrae Fructus chloroform fraction on gene expression in liver tissue of dyslipidemic mice. J. Kor. Med. Obes. Res.
15, 111-122.
15. Song, Y. R., Lim, B. U., Song, G. S. and Baik, S. H. Quality
characteristics and antioxidant activity of Makgeolli supple- mented with Omija berries (Schizandra chinensis Baillon). Kor.
J. Food Sci. Technol. 47, 328-335.
16. Szopa, A., Ekiert, R. and Ekiert, H. 2017. Current knowledge of Schisandra chinensis (Turcz.) Baill. (Chinese magnolia vine) as a medicinal plant species: a review on the bioactive com- ponents, pharmacological properties, analytical and bio- technological studies. Phytochem. Rev. 16, 195-218.
17. Tursun, X., Zhao, Y., Alat, Z., Xin, X., Tursun, A., Abdulla, R. and AkberAisa, H. 2016. Anti-inflammatory effect of Rosa rugosa flower extract in lipopolysaccharide-stimulated RAW 264.7 macrophages. Biomole. Ther. (Seoul) 24, 184-191.
18. Vezza, T., Rodriguez-Nogales, A., Algieri, F., Utrilla, M. P., Rodriguez-Cabezas, M. E. and Galvez, J. 2016. Flavonoids in inflammatory bowel disease: a review. Nutrients 8, 211.
19. Wang, X., Hu, D., Zhang, L., Lian, G., Zhao, S., Wang, C., Yin, J., Wu, C. and Yang, J. 2014. Gomisin A inhibits lip- opolysaccharide-induced inflammatory responses in N9 mi- croglia via blocking the NF-kappaB/MAPKs pathway. Food Chem. Toxicol. 63, 119-127.
20. World Health Organization. 2007. WHO monographs on se- lected medicinal plants, vol 3. Fructus Schisandrae. Geneva, 296-313.
21. Yeo, M., Suhr, Y. J. and Ham, K. B. 2006. Crossroad between inflammation and carcinogenesis in colon. Kor. J. Med. 70, 130-137.