관련 문서
Micro- and nano-sized pores were formed on the surface of the alloy using PEO and anodization methods, and the pore shape change according to the Zr
In order to analyze thermal stability and capacity loss according to the temperature of the battery cell by applying different C-rate discharge and
Second, exploratory factor analysis and Cronbach's α were conducted to analyze the validity and reliability of the questionnaire regarding participation
Glutamate excitotoxicity induced by excessive activation of NMDA receptor causes various damage to cells, which leads to cell death.. In previous studies, increased ROS
Consider the motion of a particle of mass m which is constrained to move on the surface of a cone of half-angle α and which is subject to a gravitational force g. Let the
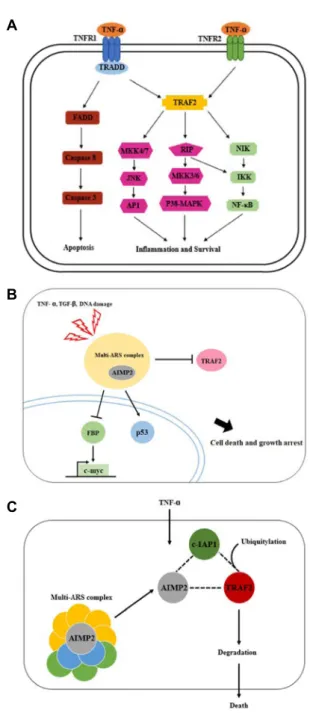
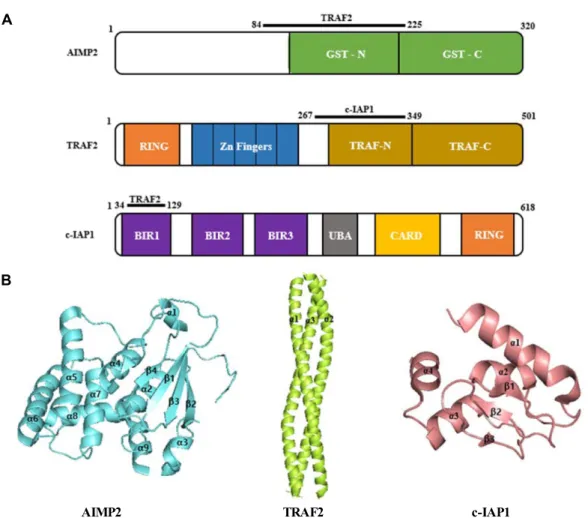
Taken together, these results suggested that latex containing the ficin inhibited the cell growth and induced apoptosis by caspase and Bcl-2 family signaling pathway in
Laser-induced doping of amorphous copper diselenide (α-CuSe 2 ) thin film with gallium (Ga) and indium (In) was performed to control/improve their electrical
Degradation of PE and PP into fuel oil by using solid acid and non-acid catalyst (Sakata et.. of liquid products from thermal and catalytic degradation of (a) HDPE at 430 °C