199 서 론
알킬페놀류인 Nonylphenol은 대표적인 내분비계장애 물질(endocrine disruptors)로 알려져 있으며, 현재까지도 지속적인 관심 대상이 되고 있는 화학물질이다. 특히 에 스트로젠 호르몬 성질(estrogenic potency)을 지니고 있어 생식호르몬계의 장애를 유발하는 것으로 알려져 있다.1)
또한 장기간 노출 시 유방암, 폐암 등의 암을 유발시킬 수 있다는 연구들이 보고된 바 있다.2,3) Nonylphenol은 세 제(detergents), 오일첨가제, 유화제(emulsifiers), 살충제, 제 초제 등을 위한 비이온성 계면활성제(non-ionic detergent) 로 사용되는 nonylphenol ethoxylates의 중간 분해산물이 다. 따라서 nonylphenol의 에스트로젠 유사성질에 따른 독성, 친지질성에 따른 생농축성 등에 의해 유럽의 국가 에서는 가정 및 산업에서 alkylphenol ethoxylate의 세제로
마이크로어레이를 이용한 내분비계장애물질 Nonylphenol에 노출된 선충에서의 유전자 발현 패턴에 관한 연구
1국립환경과학원 환경노출평가과, 2경희대학교 의과대학 약리학교실 및 기초의과학과
이철우1*․황정원2*․정화진2․서영록2․김현미1․정영희1․최경희1
A Study on the Gene Expression Profiles of Nonylphenol-exposed Nematode (Caenorhabditis elegans) Analyzed with Microarray
Chulwoo Lee1*, Jeong Won Hwang2*, Hwa Jin Jung2, Young Rok Seo2, Hyun-Mi Kim1, Young-Hee Chung1 and Kyunghee Choi1
1Environmental Exposure Assessment Division, National Institute of Environmental Research, Incheon 404-708,
2Department of Pharmacology, Institute for Basic Medical Science (IBMS), School of Medicine, Kyung Hee University, Seoul 130-701, Korea
Endocrine disruptor has been known to be one of potent hazardous chemicals able to disrupt the normal hormone signaling resulting in the development of many diseases including breast cancer. In this study, alterations of gene expression in response to endocrine disruptor nonylphenol were investigated in the nematode (Caenorhabditis elegans) as one of useful alternative animal models using a microarray containing of 22K nematode-specific probes. Upon exposure to nonylphenol for 24 hrs, we found 39 different genes that were significantly up- or down-regulated. Phosphatase, esterase and glutathion-s- transferase genes were up-regulated. In contrast, heat shock protein and connective tissue proteins were down-regulated. Our results indicate that nematode (C. elegans) is susceptible to nonylphenol toxicity and nematode microarray would be a reliable tool to speed up investigation of gene expression profiles after exposure to nonylphenol and other endocrine disruptors. (Cancer Prev Res 11, 199-204, 2006) ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ Key Words: Nematode, Caenorhabditis elegans, Microarray, Nonylphenol
책임저자:서영록, ꂕ 130-701, 서울시 동대문구 회기동 1번지 경희대학교 의과대학 약리학교실 및 기초의과학과 Tel: 02-961-0674, Fax: 02-963-0674
E-mail: dream21@khu.ac.kr
접수일:2006년 9월 6일, 게재승인일:2006년 9월 27일
*The authors are equally contributed to this work.
Correspondence to:Young Rok Seo
Departmeny of Pharmacology, Institute for Basic Medical Science (IBMS), School of Medicine, Kyung Hee University, 1, Hoegi-dong, Dong- daemun-gu, Seoul 130-701, Korea
Tel: +82-2-961-0674, Fax: +82-2-963-0674 E-mail: dream21@khu.ac.kr
서의 사용을 규제하고 있다.4)
Nonylphenol은 오수 처리과정에서 슬러지(sludge)에 다 량 함유될 수 있으며, 이러한 슬러지 가운데 일부는 토양 개선을 위해 재활용될 수 있어 nonylphenol에 의해 토양 이 오염될 수 있는 문제점을 안고 있다.5,6) 일부 조사에 따르면 캐나다에서 dry-weight sludge에서 nonylphenol을 조사한 결과, 8.4∼850 mg/kg 범위로 다량 검출된 사례가 있었다.7∼9) 내분비계장애물질을 포함한 환경오염물질에 의한 토양오염을 평가하기 위한 토양 서식 생물 종 개발 은 현재 미흡한 상황이며 따라서 토양생물 가운데 대표 적 생물인 선충, 지렁이 등에 대한 독성연구 및 표준화는 매우 필요하다고 볼 수 있다. 이러한 취지에서 본 시험에 서는 대표적 내분비계장애물질인 nonylphenol에 노출된 예쁜 꼬마선충(Caenorhabditis elegans) 내에서의 유전자 발현 패턴을 DNA 칩 마이크로어레이 시험을 통해 분석하였 다. C. elegans는 다세포 생물 가운데 최초로 게놈 정보가 규명된 생물로서 19,000여 개의 유전자를 가지고 있으며 인간과 공유하고 있는 유전자도 상당수가 되어 인체 내 독성메카니즘을 연구하는 데 있어 매우 유용한 동물이 다. 따라서 금번 시험에서 얻어진 독성자료는 nonylphe- nol에 의한 내분비계장애뿐만 아니라 생체 내 반응기작 해석에 기초를 제공할 것이며, 그 외에도 선충을 사용함 으로써 수질에 비해 상대적으로 취약한 내분비계장애물 질의 토양 및 저질독성 및 발암성에 대한 모니터링 기법 으로서의 가능성을 제시하고자 한다.
재료 및 방법 1. 시험종
시험생물은 선충(Nematode) 가운데 학명이 Caenorhab- ditis elegans (C. elegans)이며, Strain이 N2인 것을 사용하였다.
2. 시험물질의 조제
Nonylphenol을 ethanol에 의해 용해시켰으며, 1 M stock solution을 만든 후 순차적으로 희석을 하였다. 급성독성 을 위한 시험물질의 농도는 대조군(최종농도가 0.5%인 에탄올), 78.1 uM, 156.3 uM, 312.5 uM을 설정하였다.
cDNA 칩 마이크로어레이 시험을 위한 노출로는 산출된 농도(LC10)와 동일한 농도로 처리하였다.
3. 시험물질의 처리
C. elegans에 대한 Nonylphenol의 독성을 측정하기 위해 서 우선적으로 성체가 품고 있는 알(egg)들을 모아 먹이 가 없는 상태에서 하루 동안 배양하여 부화된 L1 stage
단계의 선충을 포집하였다. 포집된 C. elegans를 24 well plate에 well당 20마리씩 투입하고, 농도별로 노닐페놀을 처리한 후, 24시간 동안 배양하여 LC10을 산출하였다. 대 조군 및 처리군에서의 ethanol 최종농도를 동일하게 하였 으며, ethanol에 대한 용매대조군을 별도로 두었다.
4. RNA 추출
노출이 종료된 후 액체질소 하에서 막자사발을 이용 하여 C. elegans를 분쇄하였고, 분쇄물을 RNA 추출 키트 (QIAGEN Co.)를 이용하여 Total RNA를 정제하였다. 정 제방법은 킷트 내의 매뉴얼에 의거해 수행하였으며, 정 제된 RNA 샘플은 -80oC에 보관하였다.
5. DNA 칩 마이크로어레이 시험 및 패턴 분석
동일한 농도로 맞춘 대조군과 처리군의 Total RNA에 4μl의 oligo-(dT) primer를 넣고 3차 증류수를 첨가해 31μl 가 되도록 한 후 잘 섞은 다음, 70oC에서 10분간 가열한 후에 얼음에서 1분간 방치하여 변성시켰다. 이어서 5X First Strand Buffer 10μl, 0.1 M DTT 5μl, 50X aminoallyl- dNTP mix 1μl, RNasin (40 U/ul) 1μl, reverse transcriptase (200 U/ul) 2μl를 넣고 최종볼륨이 50μl가 되도록 한 후, 42oC에서 1시간 반응시키고 reverse transcriptase (200 U/ul) 1μl을 첨가하여 동일한 온도에서 1시간 추가 반응시켰 다. Reverse transcription 반응이 종료된 후 1 M EDTA를 16.5μl 첨가하여 65oC에서 15분 동안 반응시켜 RNA를 변성시키고, 1 M HCl 16.5μl를 넣어 반응액을 중화시켰 다. 본 시료는 마이크로어레이 시험을 위해 최종적으로 Microcon YM-30 (Millipore)을 사용해서 세척하였다. 1 M Na2CO3 (pH 9.0) 1μl을 넣고 잘 섞은 후 DMSO에 녹인 Cy3-OSu를 대조군 샘플에 첨가하고, Cy5-OSu를 처리군 샘플에 첨가하여 잘 섞은 뒤 1시간 동안 실온, 암실에서 반응시켜 coupling하였다.
Coupling 과정이 완료된 시료는 QIAquick PCR purifica- tion kit (QIAGEN)을 사용하여 재차 세척하고, 42oC에서 예열(pre-warming)한 pre-hybridization buffer에 slide를 넣고 60분 동안 활성화시켰다. 이어서 증류수에 slide를 세척 한 후, slide를 700 rpm에서 5분 동안 원심분리하여 건조 시키고 hybridization buffer를 첨가하여 95oC에서 5분 동안 가열하여 labeled sample을 변성시켰다. labeled sample을 hybridization slide에 떨어뜨린 후 cover-slip을 덮고 완전하 게 밀봉한 뒤 42oC에서 16시간 이상 반응시키고, 이어서 slide를 42oC의 washing solution I에서 5분, 실온인 washing buffer II에서 10분간, 실온인 washing buffer III에서 1분간 (모두 암실 조건에서) 반응시켰다. 상기 과정은 4회 반복
하고, 최종적으로 Slide를 700 rpm에서 5분 동안 원심분 리하여 건조시킨 후 세척 과정을 거친 다음 slide를 스캐 닝(scan)하여 마이크로에레이를 실시한 후 발현 패턴을 분석하였다.
결 과
1. Nonylphenol에 의한 선충 독성
Nonylphenol을 78.1μM∼5.0 mM 범위의 농도로 노출하 였으며, 이를 통해 선충 C. elegans의 치사율 중 LC10 (10%
치사농도)을 산출하였다(Fig. 1). 그러나 312.5μM 이상의 농도에서는 nonylphenol이 배지에서 용해되지 않음에 따 라 정확한 치사율, 특히 LC50 값은 측정하지 않았다. 시험 물질 처리 24시간 후, 78.1μM과 156.3μM의 농도에서 각 각 14%와 16.6%의 치사율을 나타내었으며, LC10은 39.7 μM (8.73 mg/L)로 산출되었다. 한편 처리군 생존개체에 서의 생장정도를 비교하였으며(Fig. 2), 그 결과 처리군
개체에서의 생장 억제 또한 대조군에 비해 강하게 영향 받은 것으로 드러났다.
2. Nonylphenol 처리에 따른 선충 내 유전자의 발현 변화
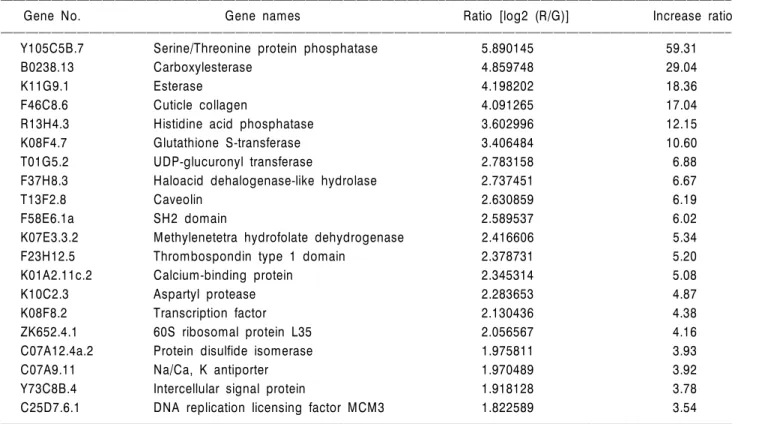
C. elegans에서 내분비계장애물질인 nonylphenol에 의해 유도되거나 억제되는 유전자들을 확인하고자 LC10 농도 의 nonylphenol을 C. elegans에 처리하였고, 그 결과 미규명 유전자(unknown genes)를 포함한 총 22,000개의 유전자 중 68개의 유전자에서 발현 변화가 관찰되었다. 그중 3 배 이상 발현이 증가되거나 억제된 유전자들을 Table 1 과 2에 각각 나타내었다. 이들 유전자 클론(clones)들의 아미노산을 이용하여 Genebank에 등록된 유전자를 검색 하였고, % identity와 E-value를 함께 나타내었다. Serine/
Threonine protein phosphatase, Carboxylesterase, Esterase, Histidine acid phosphatase, Glutathione-S-transferase, UDP- glucuronyl transferase, Haloacid dehalogenase-like hydrolase, Caveolin 등의 유전자가 LC10 농도의 nonylphenol에 의해 발현이 증가되었다. 반면, 이와 동일 농도의 nonylphenol 에서는 Collagen, Pancreatic trypsin inhibitor, Keratin-like protein, Heat shock proten HSP16-1, Cuticular collagen, Pho- sphlipase precursor 등의 유전자의 발현은 억제된 것으로 나타났다. 한편, vitellogenin, estrogen receptor 등과 같은 기 존 어류 등에서 보고된 에스트로젠성 물질에 대해 활성 이 증가되는 유전자의 발현 증가는 본 농도에서 거의 나 타나지 않았다.
고 찰
본 연구에서 발현이 유의하게 증가되거나 억제된 유 전자들 중에는 선충에 있어 현재까지 그 기능이 명확히 밝혀져 있지 않은 유전자들이 다수 포함되어 있었으며,
Fig. 2. Morphology of C. elegans exposed to nonylphenol.
Control 78.1 uM 156.3 uM
Fig. 1. Viability of C. elegans exposed to nonylphenol.
Table 2. Down-regulated genes after exposure (39.7 uM) for 24 hours
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
Gene No. Gene names Ratio [log2 (R/G)] Decrease ratio
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
T06E4.6 Collagen 0.044394 22.53
E01G6.1.1 Pancreatic trypsin inhibitor (kunitz) protein 0.08409 11.89
F07H5.8 Keratin like protein 0.09261 10.80
T27E4.8 Heat shock protein HSP16-1 0.098026 10.20
C46A5.3 Cuticular collagen 0.13765 7.26
F09C8.1 Phospholipase precursor 0.162087 6.17
Y69E1A.5 Phosphatidylethanolamine-bin ding protein 0.174087 5.74
F45G2.1 Metallopeptidase 0.198634 5.03
F28D1.5 Arabidopsis pathogenesis-related protein 0.20945 4.77
F22B7.4 Environmental stress 0.211717 4.72
C54D10.10 Serine protease inhibitor 0.216165 4.63
F38E11.1 Alpha-B-crystallin 0.21793 4.59
T04H1.3 Transthyretin-like family 0.23156 4.32
T25F10.6b.1 Calponin-like protein 0.245973 4.07
R07E5.4 Glutaredoxin 0.249379 4.01
Y37D8A.23b Pyridoxal-dependent decarboxylase conserved domain 0.25051 3.99
T05C3.8 7TM chemoreceptor, srg family 0.264188 3.79
K09C8.3 Metalloprotease 0.271208 3.69
C06B3.5 17beta-Estradiol dehydrogenase 0.296458 3.37
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ Table 1. Up-regulated genes after exposure (39.7 uM) for 24 hours
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
Gene No. Gene names Ratio [log2 (R/G)] Increase ratio
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
Y105C5B.7 Serine/Threonine protein phosphatase 5.890145 59.31
B0238.13 Carboxylesterase 4.859748 29.04
K11G9.1 Esterase 4.198202 18.36
F46C8.6 Cuticle collagen 4.091265 17.04
R13H4.3 Histidine acid phosphatase 3.602996 12.15
K08F4.7 Glutathione S-transferase 3.406484 10.60
T01G5.2 UDP-glucuronyl transferase 2.783158 6.88
F37H8.3 Haloacid dehalogenase-like hydrolase 2.737451 6.67
T13F2.8 Caveolin 2.630859 6.19
F58E6.1a SH2 domain 2.589537 6.02
K07E3.3.2 Methylenetetra hydrofolate dehydrogenase 2.416606 5.34
F23H12.5 Thrombospondin type 1 domain 2.378731 5.20
K01A2.11c.2 Calcium-binding protein 2.345314 5.08
K10C2.3 Aspartyl protease 2.283653 4.87
K08F8.2 Transcription factor 2.130436 4.38
ZK652.4.1 60S ribosomal protein L35 2.056567 4.16
C07A12.4a.2 Protein disulfide isomerase 1.975811 3.93
C07A9.11 Na/Ca, K antiporter 1.970489 3.92
Y73C8B.4 Intercellular signal protein 1.918128 3.78
C25D7.6.1 DNA replication licensing factor MCM3 1.822589 3.54
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
더욱이 nonylphenol과의 관련성이 현재까지 보고되지 않 은 유전자들이 대부분을 차지하였다.
인산화단백질의 탈인산화(dephosphorylation)는 protein phosphatase에 의해 조절되는데 protein phosphatase는 크게 두 가지 기능적 그룹으로 존재하며, 이는 Ser/Thr protein phosphatase와 Tyrosine protein phosphatase이다.10) 이 가운 데 Ser/Thr protein phosphatase는 3가지 그룹으로 다시 나 눌 수 있는데 각각을 PP1, PP2A, PP2B라 한다. 특히 PP1 은 진핵생물에서 유전적 보존성(conservation)이 매우 높 은 것으로 알려져 있으며, 생체 내에서 다양한 기능을 수행하는데 cell cycle과 같은 세포주기의 조절, 단백질 합 성, 탄화수소의 대사, 전사(transcription) 및 시그널링 등 에 관여하는 것으로 알려져 있다.11) 본 연구에서 39.7 uM 농도의 nonylphenol에 노출된 결과, Ser/Thr protein phos- phatase의 급격한 증가 양상을 보였으며, 이는 nonylphenol 이 cell cycle 등에 영향을 주어 비정상적인 탈인산화 과정 이 증가될 수 있음을 보여주고 있다. 과거 nonylphenol에 의해 mouse Sertoli 세포에서 connexin의 인산화가 감소된 보고가 있는데 이는 MAPK의 저해로 인한 것이었으며,12) MAPK의 저해와 Ser/Thr protein phosphatase의 활성과의 상관성은 선충에서 명확히 밝혀져 있지 않다. 또한 본 연구에서 탈인산화효소 가운데 histidine acid phosphatase 의 경우도 nonylphenol에 의해 10배 이상 증가함을 나타 내었다. 이 밖에도 처리군에서는 인산화효소 외에도 esterase의 증가도 관찰되었는데 특히 carboxylesterase의 발 현증가가 두드러졌으며, 해독작용과 관련된 glutathione- S-transferase도 10배 이상의 증가율을 나타내었다. 이러한 nonylphenol의 비에스트로젠(non-estrogenic) 성질에 따른 생체 내 반응의 활성화에 대해서는 아직까지 규명되지 않은 사항들이 많으며 따라서 이러한 반응에 대한 체계 적인 연구가 요구되고 있다.
Nonylphenol에 의해 억제된 유전자는 collagen, keratin like protein, cuticular protein 등 생체내 골격 또는 결체조 직과 관련된 유전자의 발현이 저해되었다. 그 외에도 Heat shock protein HSP16-1의 경우도 10배 이상의 감소를 나타내었다. HSP16-1은 선충이 어느 정도의 스트레스를 받거나, 화학물질이 노출되었을 때 증가됨으로써 생체 를 보호하기 위한 작용을 하는 것으로 알려져 있으나, 본 농도에서는 오히려 억제됨으로써 보호하는 기능이 상실되어 가는 것으로 추정되었다. 또한 상기와 같은 유 전자들이 종합적으로 감소되는 것은 선충의 성장이 처 리군에서 급격히 저해되는 것과 밀접한 관계가 있는 것 으로 사료되었다. 한편 vitellogenin, hormone receptor 등과 같은 에스트로젠 유도 유전자들의 활성 증가는 눈에 띄
게 나타나지는 않았으며, 따라서 에스트로젠성 화학물 질인 nonylphenol에 의한 이들 유전자의 활성 관계에 대 해서는 보다 저농도의 노출시험을 통한 지속적인 조사 가 필요하였다.
선충(C. elegans)은 비록 해부학적으로나 유전적인 측면 으로 볼때 간단한 생물이기는 하나, 동물로서 지녀야 하 는 기본적인 체계는 모두 갖추고 있으며, 소기관을 둘러 싸고 있는 표피가 투명하여 성체로 발달하기까지의 과 정을 관찰하기가 용이하다는 장점이 있다. 따라서 시험 물질을 처리하였을때 성장이 멈추는 시기와 치사상태를 구분할 수 있었으며, 그에 따른 유전자들의 변화 역시 마이크로에레이(microarray)라는 시험을 통하여 가능하였 다. 이는 현재 우려되고 있는 환경문제인 내분비계장애 물질을 포함한 환경유해물질을 직접 선충에 노출하여 생체 내 유전자 변화의 양상을 밝히는데 기여할 수 있을 것이다. 결론적으로 본 시험은 선충을 이용한 microarray profiling을 통해 환경오염물질의 독성을 평가하는 데 있 어 보다 구체적이고 정확한 방법이 될 것으로 보인다.
따라서 선충을 모델로 하여 발암성, 내분비계장애 등의 성질을 지닌 화학물질에 의한 유전자 발현 유형을 독성 유전체 방법을 이용해 추적함으로써 향후 이러한 물질 들의 생태위해성평가는 물론 생물학적 환경기준 설정에 유용하게 이용될 수 있을 것으로 기대된다.
참 고 문 헌
1) Ren L, Marquardt MA, Lech JJ. Estrogenic effects of nonylphenol on pS2, ER and MUC1 gene expression in human breast cancer cells MCF-7. Chemco-Biological Interactions 104, 55-64, 1997.
2) Andreescu S, Sadik OA, McGee DW. Effect of natural and synthetic estrogens on A549 lung cancer cells: correlation of chemical structures with cytotoxic effects. Chem Res Toxicol 18, 466-474, 2005.
3) Terasaka S, Inoue A, Tanji M, Kiyama R. Expression profiling of estrogen-responsive genes in breast cancer cells treated with alkylphenols, chlorinated phenols, parabens, or bis- and benzoylphenols for evaluation of estrogenic activity. Toxicol Lett 163, 130-141, 2006.
4) Renner R. European bans on surfactants trigger transatlantic debate. Environ Sci Tehnol 31, 316-320, 1997.
5) Environmental Canada. Canadian environmental quality gui- delines for nonylphenol and its ethoxylates (water, sediment and soil), Science-Based solutions Report No. 1-3, 10-38, August 2002.
6) Hawrelak M, Bennett E, Metcalf C. The environmental fate of the primary degradation products of alkylphenol ethoxylate surfactants in recycled paper sludge. Chemosphere 39, 745-752,
1999.
7) Lee HB, Peart TE. Determination of 4-nonylphenol in effluent and sludge from sewage treatment plants. Anal Chem 67, 1976-1980, 1995.
8) Lee HB, Peart TE, Bennie DT, Maguire RJ. Determination of nonylphenol-polyethoxylates and their carboxylic acid metabolites in sewage treatment plant sludge by supercritical carbon dioxide extraction. J Chromatogr 785, 385-394, 1997.
9) Bennie DT, Sullivan CA, Lee HB, Maguire RJ. Alkylphenol polyethoxylate metabolites in Canadian sewage treatment plant waste streams. Water Qual Res J Can 33, 231-252, 1998.
10) Konstantina N, Huong H, Scott W, Surojit P, Nunzio B,
Kjetil T, Paul J, Tomas M. Haematopoietic protein tyrosine phosphatase (HePTP) phosphorylation by cAMP-dependent protein kinase in T-cells: dynamics and subcellular location.
Biochem J 378, 335-342, 2004.
11) Yamamoto M, Suzuki Y, Kihira H, Miwa H, Kita K, Nagao M, Tamura S, Shiku H, Nishikawa M. Expressions of four major protein Ser/Thr phosphatases in human primary leukemic cells. Leukemia 13, 595-600, 1999.
12) Aravindakshan J, Cyr DG. Nonylphenol alters connexin 43 levels and connexin 43 phosphorylation via an inhibition of the p38-mitogen-activated protein kinase pathway. Biol Reprod 72, 1232-1240, 2005.