92 http://dx.doi.org/10.7850/jkso.2015.20.2.92
Free Access
수치모델을 활용한 자란만에서 유해 와편모조류 Cochlodinium polykrikoides와 규조류 Skeletonema sp.의 경쟁에 미치는 영양염의 영향
오석진1 · 김현정1 · 권형규2 · 양한섭1 · 김석윤1,*
1부경대학교 해양학과
2부경대학교 해양과학공동연구소
Effect of Nutrients on Competition among the Harmful Dinoflagellates Cochlodinium polykrikoides and the Diatom Skeletonema sp. in Jaran Bay
Using a Mathematical Model
S
EOKJ
INO
H1, H
YUNJ
UNGK
IM1, H
YEONGK
YUK
WON2, H
AN-S
OEBY
ANG1 ANDS
EOKY
OONK
IM1,*
1Department of Oceanography, Pukyong National University, 45 Yongso-ro, Nam-Gu, Busan, 608-737 Korea
2Korea Inter-University Institute of Ocean Science, Pukyong National University, 45 Yongso-ro, Nam-Gu, Busan, 608-737 Korea
수치모델을 활용한 Cochlodinium polykrikoides와 Skeletonema sp.의 영양염에 대한 종간경쟁을 바탕으로 2013년 하계 자란만에서 C. polykrikoides의 적조 미발생 원인을 고찰하였다. 2013년 하계 자란만의 영양염 조건을 모델에 적용시킨 결과, 실제 현장과 유사하게 Skeletonema sp.의 세포밀도는 C. polykrikoides보다 높았다. 또한 다양한 변수에 대한 민감도 분석에서, C. polykrikoides는 어떠한 변수의 변화에도 일정한 세포밀도를 보였으며, Skeletonema sp.의 세포밀도는 영양염 농도의 증·감에 민감하게 반응하였다. 이러한 결과를 통해 C. polykrikoides는 급격한 환경변화 에도 영향을 받지 않고 생장을 유지할 수 있을 것이다. 하지만 영양염 공급이 변화하는 연안환경에서는 Skeletonema sp.의 생장이 촉진될 수 있을 것이다. 따라서 외부로부터 영양염의 공급이 크게 변화하는 자란만에서 Skeletonema sp.와 같은 규조류의 번무에 따라서 C. polykrikoides의 출현이 억제된 것으로 보인다.
We investigated cause of non-outbreak of Cochlodinium polykrikoides blooms in the western coast of Jaran Bay during summer, 2013, based on the species competition among C. polykrikoides and Skeletonema sp. using a mathematical model. As a result of the model simulation where the nutrient conditions in Jaran Bay was applied during summer, the cell density of Skeletonema sp. was higher than that of C. polykrikoides. In the sensitivity analyses by doubling and halving the parameters, any parameter had little effect on the cell density of C. polykrikoides. The cell density of Skeletonema sp. was significantly affected by changes in the parameter values. These results indicated that the growth of C. polykrikoides could be unaffected by rapidly changing environments. However, the growth of Skeletonema sp. may have been promoted by the changing nutrient supply of coast environments. Therefore, C. polykrikoi- des might have been suppressed by diatom blooms, such as Skeletonema sp., in changing nutrient supply condi- tion of Jaran Bay.
Keywords: Cochlodinium polykrikoides, Skeletonema sp., Dissolved nutrient, Mathematical model, Species competition
서 론
연구해역인 자란만은 남해 동부해역에서 위치하고 있으며, 만 북부에는 5개의 하천이 만내로 영양염을 공급하여 일차생산자의 생장에 있어 유리한 환경이 조성되어 있다. 이로 인하여 굴 수하
식, 우렁쉥이 등의 양식어장, 가두리 및 정치망어장 등을 이용한 양식산업이 활발하며, 수출용 패류생산지정해역으로도 지정되어 있다(MIFAFF, 2008). 하지만 자란만도 다른 남해안의 폐쇄성 연 안역과 같이 부영양화, 빈산소 및 적조 등에 따라 어장생산성이 악 화되고 있으며, 특히 2014년의 경우 Cochlodinium polykrikoides의 적 조가 만 입구까지 고밀도로 발생하였다(http://portal.nfrdi.re.kr/
redtide). 2013년 동 종에 따라 남해안과 동해안에서 247 억원의 Received February 5, 2015; Revised March 16, 2015; Accepted May 11, 2015
*Corresponding author: [email protected]
2000). 이러한, 환경에서 일차생산자는 19세기에 제시된 Liebig 최 소율의 법칙(Law of minimum)에 따라 생장이 제한이 되며(Ryther and Dunstan, 1971), Tilman(1982)은 영양염의 절대적인 농도보다 도 영양염의 구성비에 따라서 식물플랑크톤 생장에 영향을 미친 다고 보고하여, 제한 영양염의 화학양론비(stoichiometry ratio)의 변화는 결국 식물플랑크톤의 생장, 군집조성 및 종 천이 등에 영향을 미치는 것으로 알려져 있다(Justic et al., 1995; Arrigo, 2004).
한편, 영양염에 따른 식물플랑크톤의 생장식과 이를 이용한 종간 경쟁 수치모델은 많은 연구에서 보고되어 있다. Tilman(1981)은 식물플랑크톤이 외부 영양염에 따라 생장한다는 Monod 식을 활 용한 자원량 경쟁 모델을 구축하여 종 천이를 설명하였다. 하지만, 세포 내 영양염 함유량(cell quota)을 고려한 첫 번째 모델은 그 보 다 앞선 Droop(1973)에 의해 보고되었으며, Lehman et al.(1975) 에 따라 다중 영양염 모델(multiple nutrient model)로 발전되었으 며, 영양염 흡수(Michaelis-Menten 식)와 생장(Droop 식)을 분리 시켜 모델을 설계하였다. Roelke et al.(1999)은 앞선 경쟁 모델을 더욱 발전시켜 비정상상태에서 영양염과 식물플랑크톤 생장에 의 한 재생산과정을 모델화하여, 식물플랑크톤의 군집구조가 어떻게 변화하는지 확인하였다. 지금까지 모델은 대부분이 용존태 무기 영양염만 고려한 모델이다. 하지만 일부 해역에서 용존태 유기 영 양염이 무기 영양염과 유사하거나 높은 비율을 보이기 때문에 무기 영양염 제한시 유기 영양염이 식물플랑크톤 생장에 중요한 영양염 공급원이 될 수 있다(Cembella et al., 1984; Kwon et al., 2013).
본 연구에서는 용존태 영양염을 첨가한 배지에서 유해 와편모 조류 C. polykrikoides와 우리나라 연안에서 우점하여 출현하는 대 표적인 규조류인 Skeletonema sp.의 혼합배양을 실시하고, 그 결과를 이용하여 Truncation 모델(Dugdale et al., 1981)을 기반으로 하는 종간 경쟁모델을 구축하였다. 또한 구축된 모델에 2013년 하계 자 란만의 영양염 자료를 입력하여, C. polykrikoides의 적조 미발생을 식물플랑크톤의 종간 경쟁으로 고찰하였다.
재료 및 방법
C. polykrikoides와 Skeletonema sp.의 분리 및 유지배양
C. polykrikoides는 한국해양과학기술원 해양시료도서관(No.LMBE-VI)에서 순수 분리한 종을 분양받아 이용하였으며, Skeletonema sp.는 2013년 4월 자란만의 서쪽 연안(34°55'30.59''N, 128°12'17.77''E) 의 표층해수에서 pasteur pipette(Ø 50~100 μm)을 이용하여 분리 하였다. 분리한 세포는 여과해수를 이용하여 3~4회 반복 세척 하
및 건조 멸균(120oC, 3 hr) 후 사용하였으며, 모든 실험은 무균대 에서 수행하였다.
C. polykrikoides와 Skeletonema sp.의 단일 및 혼합 배양
C. polykrikoides와 Skeletonema sp.의 질소 및 인에 대한 종간 경쟁을 살펴보기 위해 단일 및 혼합배양을 실시하였다. 실험에 사 용된 배지는 인공해수(AK; artificial Keller)를 기반으로 반 L1 배 지이며(Keller et al., 1987), 질소 원과 인 원으로 각각 질산염과 인산염을 사용하였다. 질산염은 최종농도가 19 μM, 인산염은 1 μM 이 되도록 조제하였으며, C. polykrikoides의 단일 배양을 위한 배 지는 Skeletonema sp.의 단일 배양 및 두 종의 혼합 배양을 위한 배지와는 달리 규산염을 첨가하지 않았다. 배양은 2 L의 Erlenmeyer flask를 이용하였으며, 인산염과 질산염이 첨가된 1.5 L의 배지에 대수증식기 후기의 세포를 약 100 cells mL-1이 되도록 접종하였 다. 배양온도와 염분 및 광량은 유지배양과 동일한 조건으로 설정 하였다. 세포밀도는 매일 일정한 시간에 도립현미경(TE 2000-S, Nikon)을 이용하여 계수하였으며, 영양염 농도 측정을 위한 시료는 이틀에 한 번씩 획득하였다. 획득한 배양시료는 유리 섬유 여과지 (GF/F, 47 mm, 0.7 μm pore size, Whatman)를 이용하여 식물플랑 크톤 세포가 파괴되지 않는 30 mm Hg 이하의 낮은 압력 하에서 진공여과한 후, 해양환경공정시험기준(MLTM, 2010)에 의거하여 측정하였다. 그리고 모든 실험은 duplicate로 실시하였다.수치모델의 구성
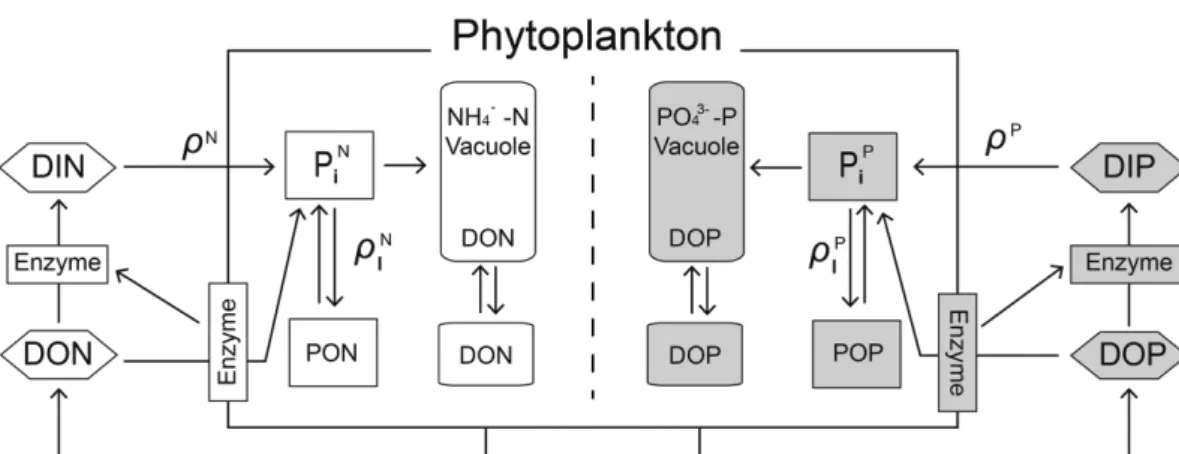
식물플랑크톤의 영양염 흡수는 환경수 중의 영양염 농도에 반 응하지 않고 세포 내 영양염 함유량에 의존하며, 시간에 따라 변 화한다고 가정하였다(Dugdale et al., 1981). 식물플랑크톤의 상대 생장속도(specific growth rate)는 세포내의 영양염 함유량에 의해 조절된다. Fig. 1은 모델의 근간이 되는 개념도이며, 식물플랑크톤의 내부에는 internal pool(PiN, PiP)과 고분자량의 질소 및 인 화합물 (PON, POP)이 존재하는 것으로 구성하였다. 식물플랑크톤에 의 해 흡수된 영양염은 internal pool로 이동한 후 동화작용에 의해 체내구성물질(PON과 POP)로 합성된다. 그리고 환경수 중의 영양 염이 일정 수준 이하로 감소하여 식물플랑크톤의 생장에 제한요인으 로 작용하며, 고갈이 되면 internal pool의 영양염을 전부 사용한 후 생장을 멈추게 된다. 한편, internal pool이 포화되면 세포는 더 이 상 영양염을 흡수하지 못하며, 흡수속도는 세포의 구성 물질과 세 포내 축적물질 등의 동화속도에 의해 제한된다. 본 연구에서는 세 포내 영양염 함유량(cell quota; internal pool+세포의 구성 물질과 세
포내 축적물질)이 최대에 이르면, 세포의 구성물질과 세포내 축적 물질이 소모될 때까지 흡수속도를 0으로 설정하였다. 모델에서 C.
polykrikoides와 Skeletonema sp.의 영양염 흡수(ρ)는 Michaelis- Menten 식에 근거한 효소활성 반응에 따라 흡수되는 것으로 가정 하였다(Dugdale, 1967).
(1)
ρN,P: 질소 또는 인 흡수속도(nitrogen or phosphorus uptake rate;
pmol cell-1 hr-1)
: 최대 질소 또는 인 흡수속도(maximum nitrogen or phosphorus uptake rate; pmol cell-1 hr-1)
KsN,P: 질소 또는 인에 대한 반포화상수(half-saturation constant of nitrogen or phosphorus; μM)
SN,P:배양액 중 질소 또는 인의 농도(external nitrogen or phosphorus concentration; μM)
C. polykrikoides와 Skeletonema sp.의 생장(µ)은 세포내 영양염 함유량에 따라 이뤄지는 것으로 가정하였다(Droop, 1973).
(2)
µN,P: 질소 또는 인에 대한 상대생장속도(specific growth rate on nitrogen or phosphorus; day-1)
µ'maxN,P: QN 또는 QP가 무한대일 때 최대상대생장속도(maximum growth rate when QN or QP is infinity; day-1)
: 세포내 최소 질소 또는 인 함유량(minimum nitrogen or phosphorus cell quota; pmol cell-1)
QN,P: 세포내 질소 또는 인 함유량(cell quota of nitrogen or phosphorus;
pmol cell-1)
여기서 QN과 QP는 각각 질소의 internal pool(PiN)과 PON의 합 그리고 인의 internal pool(PiP)과 POP의 합으로 구성되었다. S, Q, Pi의 시간에 따른 변화는 다음과 같다.
(3) (4)
(5) 세포밀도(N)의 시간에 따른 변화는
(6) 로 계산될 수 있다. 그러나 제한된 공간에서는 영양염이 풍부하더 라도 세포의 생장에 따라 배출된 화학물질(waste material)과 공간의 제약이 발생하므로, Michaelis-Menten 식과 Droop 식을 이용한 생 장곡선은 배치배양 상태의 세포밀도를 재현하기 어렵다(Oh, 2002).
이에 Yamamoto(1999)는 내적자연증가율(intrinsic rate of natural increase)을 표현할 수 있는 지수함수를 제안하였으며, 다음과 같이 나 타낼 수 있다.
(7) r: 내적자연증가율(intrinsic rate of natural increase; day-1) h: Verhulst-Pearl coefficient
정상기의 환경수용력은 K=r/h로 계산되며, Droop 식은 다음과 같이 변형된다.
(8) 또한, C. polykrikoides와 Skeletonema sp.의 생장속도는 복수 영 양염 가운데 제한되는 영양염에 따라 생장이 억제되는 Blackman의 최소율 법칙(Blackman’s rate-limiting concept)을 이용하였다(Cullen et al., 1992).
(9) 용존 유기 영양염은 식물플랑크톤에 흡수된 후 효소에 의해 가 수분해 되어 용존 무기 영양염의 형태로 이용되지만, 해수 중 존 재하는 효소에 의해 용존 무기 영양염의 형태로 전환된 후 식물 플랑크톤의 생장에 이용되기도 한다. 따라서 본 모델에서는 다음 과 같이 용존 유기 영양염의 가수분해가 가능하도록 설정하였다.
(10) (11) H: 가수분해율(hydrolysis rate; %)
ρN P, ρmaxN P, SN P, KsN P, +SN P, ---
⋅
=
ρmaxN P,
μN P, μ′maxN P, 1 Q– minN P,
QN P, ---
⎝ ⎠
⎛ ⎞
⋅
=
QminN P,
dS/dt=–ρN dQ/dt ρ μQ= –
dPiN P, /dt ρ ρ= – I
dN/dt μN=
dN/dt N r hN= ( – )
dN/dt N μ= {( max(1 Q– min/Q)) hN– }
μ MIN μ= { maxN (1 Q– minN /QN) μ, maxP (1 Q– minP /QP)}
DIN=DON H⋅ DIN=DOP H⋅
Fig. 1. A conceptual model showing uptake and assimilation process in phytoplankton cell. ρ: uptake, ρI: assimilation, N; external nitrogen, P; external phosphorus, PiN: intracellular nitrogen pool, PiP: intracellular phosphate pool, PON: structural and other storage nitrogen com- ponent, POP; structural and other storage phosphorus component.
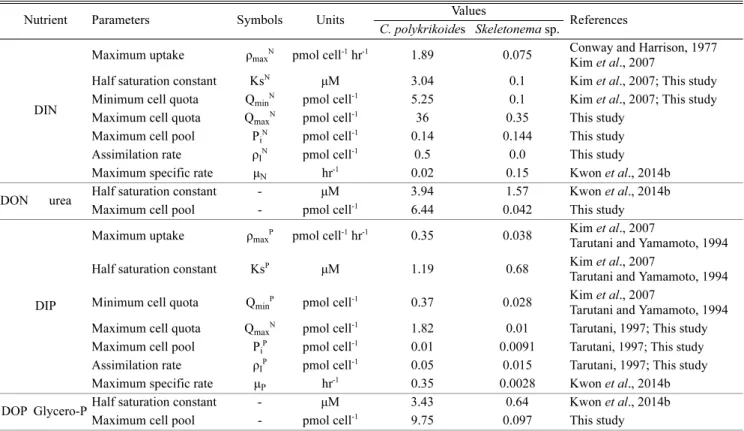
본 논문의 수치모델은 수치모델 프로그램인 STELLA(version 10.0.6; High Performance System Inc., USA)를 이용하였으며, 모 델에 적용된 변수들은 Table 1에 표시하였다.
2013년 하계 자란만의 영양염 조건에서 구현된 종간 경쟁 모델
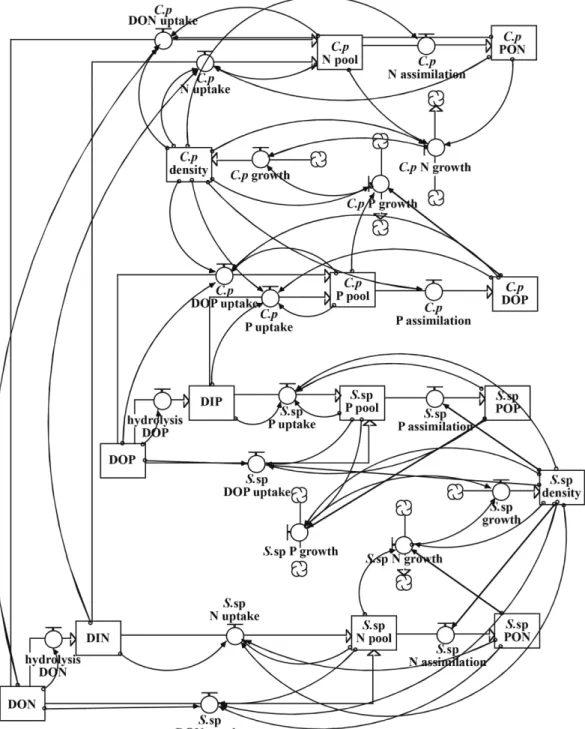
2013년 하계 자란만의 영양염 조건에서 구현된 모델을 이용하여 C. polykrikoides와 Skeletonema sp.의 종간 경쟁을 검토하였다. 모 델에서 용존태 무기 질소(dissolved inorganic nitrogen; DIN), urea, 용존태 무기 인(dissolved inorganic phosphorus; DIP) 그리고 용존태 유기 인(dissolved organic phosphorus; DOP) 농도는 Kwon et al.(2014a)에 의해 보고된 자란만의 현장농도인 각각 2.62 μM, 1.08 μM, 0.56 μM 및 0.33 μM로, 2013년 7∼9월에 자란만 북서쪽해역에서 측정된 평균 농도이다. 그리고 C. polykrikoides와 Skeletonema sp.의 초기 세포밀도는 1.0×102 cells mL-1로 설정하였다. 또한 감도분석을 수행하기 위해 각각의 변수들을 2배로 증·감시킨 후, C. polykrikoides 와 Skeletonema sp.의 세포밀도를 살펴보았다. 감도 분석에 이용된 변 수로는 urea의 가수분해율, DOP의 가수분해율, DIN, urea, DIP, DOP, C. polykrikoides의 초기 세포밀도 그리고 Skeletonema sp.의 초 기 세포밀도이며, C. polykrikoides와 Skeletonema sp.에 가장 큰 영향을 미치는 변수를 파악하기 위해 실시하였다. 그리고 모델의 기본 구성은 Fig. 2, 사용된 STELLA 프로그램의 모식도는 Fig. 3에 나타내었다.
결 과
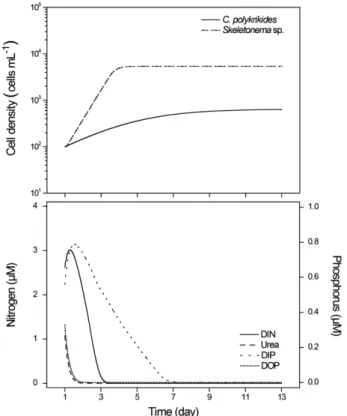
수치모델을 이용한 C. polykrikoides와 Skeletonema sp.의 단일배양 재현
Fig. 4에 C. polykrikoides와 Skeletonema sp.의 단일배양에서 재 현한 수치모델의 결과를 나타내었다. C. polykrikoides의 단일배양 에서 배지 중 질산염은 배양 개시 후 9일째 1.02 μM로 급격하게 감소하였으며, 이후에는 약 1 μM 이하의 농도를 보였다. 인산염의 Maximum cell pool - pmol cell-1 6.44 0.042 This study
DIP
Maximum uptake ρmaxP pmol cell-1 hr-1 0.35 0.038 Kim et al., 2007
Tarutani and Yamamoto, 1994 Half saturation constant KsP μM 1.19 0.68 Kim et al., 2007
Tarutani and Yamamoto, 1994 Minimum cell quota QminP pmol cell-1 0.37 0.028 Kim et al., 2007
Tarutani and Yamamoto, 1994 Maximum cell quota QmaxN pmol cell-1 1.82 0.01 Tarutani, 1997; This study Maximum cell pool PiP pmol cell-1 0.01 0.0091 Tarutani, 1997; This study Assimilation rate ρIP pmol cell-1 0.05 0.015 Tarutani, 1997; This study Maximum specific rate μP hr-1 0.35 0.0028 Kwon et al., 2014b DOP Glycero-PHalf saturation constant - μM 3.43 0.64 Kwon et al., 2014b
Maximum cell pool - pmol cell-1 9.75 0.097 This study
Fig. 2. A schematic diagram of the simulation model for species competition. Both DIN and DON were used as the source of nitro- gen. Both DIP and DOP were used as the source of phosphorus.
농도는 배양 개시 후 3일째 0.20 μM까지 급격하게 감소하였으며, 13일째 검출한계 이하의 농도를 보였다. 그리고 C. polykrikoides의 세 포밀도는 약 10일째 정상기에 도달하였으며, 최대 세포밀도는 6.50×102 cells mL-1로 나타났다. Skeletonema sp.가 접종된 배지의 질산염은 배양 개시 후 3~9일 사이에 급격한 농도 감소를 보였으며, 9일 이후부터는 약 1 μM이하로 검출되었다. 인산염은 C. polykrikoides 의 경우와 마찬가지로 배양 개시 후 3일째 0.67 μM까지 급격하게 감 소하였으며, 7일째부터는 0.50 μM 이하의 농도로 나타났다.
Skeletonema sp.의 세포밀도는 6일째에 정상기에 도달하였으며, 최대
세포밀도는 5.24×104 cells mL-1로 나타났다. 또한 C. polykrikoides와 Skeletonema sp.의 단일배양으로부터 Verhulst-Pearl coefficient(h)를 구하였으며, h는 실제 배양에 따른 세포밀도의 증가에 근사해가는 점근법에 따라 획득하였다. 그 결과 h는 C. polykrikoides에서 3.1×10-8그리고 Skeletonema sp.에서 8.10×10-10로 나타났다. 안정 기(K=r/h)에서 C. polykrikoides와 Skeletonema sp.에 대한 환경수 용력은 각각 6.40×102 cells mL-1과 6.16×104 cells mL-1로 나타났 으며, 이들 결과는 단일배양에서 관찰된 각 종의 최대 세포 밀도와 유사하였다.
Fig. 3. The structure of the simulation model of species competition drawn by the STELLA software. Both DIN and DON were used as the source of nitrogen. Both DIP and DOP were used as the source of phosphorus.
수치모델을 이용한 C. polykrikoides와 Skeletonema sp.의 혼합배양 재현
Fig. 5에 C. polykrikoides와 Skeletonema sp.의 혼합배양 및 혼 합배양을 재현한 수치모델의 결과를 나타내었다. 혼합배양에서 질 산염은 배양 개시 후부터 5일째까지 급격한 감소를 보였으며, 7일 이후부터는 약 1 μM의 농도였다. 인산염은 3일째 0.52 μM까지 급격하게 감소하였으며, 배양 개시 후 7일째에는 0.30 μM 이하의 농도를 보였다. 그리고 C. polykrikoides와 Skeletonema sp.의 세 포밀도는 각각 6일과 4일째 정상기에 도달하였으며, 최대 세포밀 도는 각각 3.90×102 cells mL-1, 9.83×103 cells mL-1로 나타났다.
혼합배양을 재현한 모델에서 질산염과 인산염은 식물플랑크톤의 생장과 동시에 급격히 감소하여 4일째 완전히 고갈되었다. C.
polykrikoides의 세포밀도는 지속적으로 증가하여 13일째 6.40×102 cells mL-1에 도달한 반면 Skeletonema sp.는 5일째 정상기에 도달 하여 5.73×103 cells mL-1의 최대 세포밀도를 보였다.
2013년 하계 자란만에서 C. polykrikoides와 Skeletonema sp.의 종간 경쟁
2013년 7~9월에 영양염 조건을 모델로 구현한 결과, DIN과 DIP의 농도는 배양 개시 후부터 2일째까지 증가하였으며, 증가량은 각각 0.40 μM과 0.23 μM이었다(Fig. 6). 이 후 DIN과 DIP는 점차 감소하여 각각 3일 그리고 7일째 완전히 고갈되었다. DON과 DOP의 농도는 식물플랑크톤의 생장에 따라 급격히 감소하여 1일째 완전히 고갈되었다. C. polykrikoides의 세포밀도는 10일째에 정상기에 도 달하였으며, 최대 세포밀도는 6.30×102 cells mL-1로 나타났다.
Fig. 4. Simulation results of cell growth using the Michaelis-Menten equation and the Droop equation. The curves and symbols are the model output and experimental results, respectively.
Fig. 5. Simulation of the species competition experiments. Batch culture with 19.2 μM DIN and 1.2 μM DIP. Cell densities of C.
polykrikoides and Skeletonema sp., Nutrient concentrations of the ambient water, The curves and symbols are the model output and experimental results, respectively.
Fig. 6. Simulation of species competition assuming the environ- mental condition of Jaran Bay in summer.
Skeletonmea sp.의 경우, 약 3일째 정상기에 도달하여 8.11×103 cells mL-1의 최대 세포밀도를 보였다.
변수들의 증·감에 따른 모델의 감도 분석 결과, C. polykrikoides의 세포밀도는 C. polykrikoides 초기 세포밀도를 증·감시킨 경우에만 반응을 보였다(Fig. 7). C. polykrikoides의 초기 세포밀도를 2배로
증·감 시킨 경우, C. polykrikoides 세포밀도는 각각 1.0% 증가, 2.1% 감소를 보였으며, Skeletonema sp. 세포밀도는 각각 60.3%
감소, 109% 증가를 보였다. Skletonema sp.의 초기 세포밀도를 증·감시킨 경우에, Skeletonema sp. 세포밀도는 각각 50.2% 증가, 37.5% 감소하였다. Skeletonema sp.의 최대 세포밀도는 DIN의 Fig. 7. Sensitivity analyses for species competition assuming environmental condition of Jaran Bay in summer.
(3) 흡수한 영양염을 세포 내로 축적하는 저장전략(storage strategy)으로 구분하여 제시하였다. Skeletonema sp.는 C. polykrikoides와 비교 하여 빠른 생장속도를 지니며, 상대적으로 DIN 및 DIP에 대한 친 화성이 높은 종으로 친화성 및 생장전략을 가지는 대표종으로 알 려져 있다(Noh et al., 2010; Kwon et al., 2014b). 실제 단일 및 혼 합배양에서 Skeletonema sp.는 C. polykrikoides보다 빠르게 정상 기에 도달하였으며 세포밀도도 높았다. 반면 C. polykrikoides는 혼 합배양과 이를 재현한 모델에서 7일 이후 배양액 중 영양염이 고갈되었음에도 불구하고 세포밀도가 증가하였다. 특히, C.
polykrikoides는 Skeletonema sp.보다 세포 내 질소 및 인의 함 유량이 큰 종으로 보고되어있기에(Table 1), 흡수한 질소 및 인을 세 포내에 축적하여 생장하는 저장전략을 가지는 것으로 보인다.
한편, 2013년 하계 자란만 영양염 조건을 모델에 적용시킨 결과, Skeletonema sp.의 세포밀도는 C. polykrikoides 보다 높게 나타났 으며, 실제 해역에서의 세포밀도보다 높았다. 과거 규조류의 주요 포식자는 요각류(copepod)로 여겨져 왔으나, Jeong et al.(2004)에 의해 요각류인 Acartia spp.가 S. costatum 개체군의 최대 5%를 포식하는 것으로 보고되어, S. costatum에 대한 요각류의 포식압은 크지 않은 것으로 밝혀졌다. 반면 종속영양성 와편모조류 Protoperidinium bipes는 S. costatum 개체군의 최대 약 70%를 포식할 수 있는 것으로 보고되어(Jeong et al., 2004), Protoperidinium spp.가 요각류보다 S. costatum의 세포밀도에 큰 영향을 주는 것으로 나타났다. 실제 자란만의 현장관측 결과, 종속영양성 와편모조류 Protoperidinium spp.는 2013년 5~10월 동안 꾸준히 출현하였으며, 특히 P. bipes 는 하계에 집중적으로 출현하였다(Kwon et al., 2014a). 따라서 모 델에서 Skeletonema sp.의 세포밀도는 모델의 단순화로 포식압을 제외하였기에 실제 현장보다 높게 평가된 것으로 보인다. 반면에, C.
polykrikoides의 포식생물은 종속영양성 와편모조류 Polykrikoides kofoidii와 Noctiluca scintillans 및 무피성 섬모류(nakted ciliate) Strombidinopsis spp. 등으로 보고되어 있으나, 하계 자란만에서 Polykrikoides spp.와 Noctiluca scintillans는 출현하지 않았으며 (Kwon et al., 2014a), C. polykrikoides를 포식하는 섬모류의 경우 크기가 150 μm 이상 되는 소수 Strombidinopsis spp.만이 효과적 으로 포식할 수 있는 것으로 보고되어 있어(Jeong et al., 1999), 포식압에 의한 C. polykrikoides 세포밀도 영향은 없었을 것으로 생각된다.
식물플랑크톤은 용존태 유기 질소(dissolved organic nitrogen;
DON)와 DOP를 이용하기 위해 가수분해효소가 필요하며, 화합물 의 종류에 따라 다양한 효소가 존재한다. DON 중 urea는 urease에
(1993)의 결과를 참고하여 용존 유기 영양염의 가수분해율을 15%로 설정하였다. 하지만 가수분해율을 2배로 증·감시킨 민감도 분석결과 (Fig. 7), C. polykrikoides의 세포밀도는 아무런 변화를 보이지 않 았으며, Skeletonema sp.의 세포밀도는 2%이하의 증·감을 보여, 본 모델에서 가수분해율은 C. polykrikoides와 Skeletonema sp.의 밀 도변화에 거의 영향을 미치지 않는 것으로 나타났다.
감도분석에서 C. polykrikoides의 세포밀도는 어떤 환경변수에도 크 게 영향을 받지 않는 것으로 나타나, 영양염 제한상태에서도 기회 종보다 영향을 받지 않고 생장할 수 있을 것이다. 하지만, 연안내 만역의 경우 담수유입 및 영양염 용출 등과 같은 지속적인 영양 염 공급이 발생하며, 기회종인 Skeletonema sp.의 세포밀도는 외 부 영양염 공급에 따라 크게 변화하기 때문에 C. polykrikoides보 다 빠르게 생장하여 번무할 가능성이 있으며, 실제 하계 자란만은 규조류의 극우점 현상이 나타나(Kwon et al., 2014a), 종간 경쟁 에 따라 자란만 내만역으로 C. polykrikoides 적조가 확산 되지 못 할 것으로 보인다.
본 연구결과를 바탕으로 2013년 하계 자란만 내의 내만역에서 C. polykrikoides 적조 미발생 이유를 다음과 같이 생각할 수 있다.
자란만 인근 해역에서 관측된 풍향의 수평유동성분(u, v)을 시계열의 stick vector diagram으로 나타낸 결과, 2013년 7월에는 남서풍이 1.1~4.9 cm s-1, 8월에는 남동풍이 0.8~2.8 cm s-1의 속도를 보이며 우세하게 나타났다(Fig. 8). 2013년 9월에는 1.6~4.3 cm s-1의 동 풍계열과 1.2~2.7 cm s-1의 남풍계열이 번갈아가며 나타났다. 이로 인해 C. polykrikoides 적조 발생 초기에는 적조패치가 점차 동쪽으로 확산되어 섬의 서쪽에 집적되는 현상을 보였다. C. polykrikoides 적조 발생 중기에는 남동풍의 영향으로 적조가 연안역으로 유입 되었으며, 섬의 동쪽과 내만지역에서도 적조가 관측되었다. 하지 만 통영과 인접한 자란만에서는 C. polykrikoides의 유영세포는 발 견되지 않았으며, 오히려 규조류의 극우점현상이 관측되었다(Kwon et al., 2014a). C. polykrikoides의 유영세포는 남동풍의 영향으로 통영인근 해역에서 자란만 내로 유입되었을 가능성이 있으며, 만내로 유입된 C. polykrikoides는 제한 영양염에 대해 규조류와 경쟁관계에 위치하였을 것이다(Fig. 8). Skeletonema sp.는 C. polykrikoides보다 용존 유기 영양염에 대한 이용능력이 높고, 용존태 영양염에 대한 친화성이 크기 때문에(Kwon et al., 2014b), 두 종의 종간 경쟁에서 C. polykrikoides가 Skeletonema sp.보다 불리한 위치에 있었을 것 으로 생각된다. 결과적으로 해양 생태계의 다양한 환경 요인 중 이·
화학적 요인만을 고려하였을 경우, 2013년 하계 자란만은 C.
polykrikoides보다 Skeletonema sp.의 생장에 유리한 환경으로 생
각되며, 이는 2013년 하계 자란만에서 규조류가 극우점하여 C.
polykrikoides의 유입이 억제되었을 것으로 판단된다.
사 사
이 논문은 부경대학교 자율창의학술연구비(2014년)에 의하여 연 구되었습니다.
참고문헌(References)
Arrigo, K.R., 2004. Marine microorganisms and global nutrient cycles.
Nature, 437: 349−355.
Borchardt, M.A., 1996. Nutrients. In: Algal Ecology: Freshwater Benthic Ecosystems, edited by Stevenson, R.J., M.L. Bothwell and R.L. Lowe, pp. 184−228.
Bronk, D.A., 2002. Dynamics of organic nitrogen. In: Biogeochem- istry of marine dissolved organic matter, edited by Hansell, D.A.
and A.C. Craig, Academic Press, San Diego, pp. 153−247.
Cembella, A.D., N.J. Antia and P.J. Harrison, 1984. The utilization of inorganic and organic phosphorus compounds as nutrients by eukaryotic microalgae: a multidisciplinary perspectitive: Part I.
CRC Crit. Rev. Microbiol, 10: 317−391.
Conway, H.L. and P.J. Harrison, 1977. Marine diatoms grown in che- mostats under silicate or ammonium limitation. IV. Transient response of Chaetoceros debilis, Skeletonema costatum, and Thalassiosira gravida to a single addition of the limiting nutrient. Mar. Biol., 43: 33−43.
Cullen, J.J., X. Yang and H.L. MacIntyre, 1992. Nutrient limitation of marine photosynthesis. In: Primary productivity and biogeo- chemical cycles in the sea, edited by Falkowski, P.G. and A.
Woodhead, Plenum Press, New-York, pp. 69−88.
Dugdale, M.R., 1967. Nutrient limitation in the sea: dynamics, iden- tification and significance. Limnol. Oceanogr., 12: 685−695.
Droop, M.R., 1973. Some thoughts on nutrient limitation in algae1.
J. Phycol., 9: 264−272.
Dugdale, R.C., B.H. Jones Jr, J.J. MacIsaac and J.J. Goering, 1981.
Adaptation of nutrient assimilation. Can. Bull. Fish. Aquat. Sci., 201: 234−250.
Doblin, M.A., S.I. Blackburn and G.M. Hallegraeff, 1999. Growth and biomass stimulation of the toxic dinoflagellate Gymnodinium catenatum (Graham) by dissolved organic substances. J. Exp. Mar.
Biol. Ecol., 236: 33−47.
Jeong, H.J. J.H. Shim, J.S. Kim, J.Y. Park, C.W. Lee and Y. Lee, 1999. Feeding by the mixotrophic thecate dinoflagellate Fragi- lidium cf. mexicanum on red-tide and toxic dinoflagellates. Mar.
Ecol. Prog. Ser., 176: 263−277.
Jeong, H.J., Y.D. Yoo, J.S. Kim, T.Y. Kim, J.H. Kim, N.S. Kang and W.H. Yih, 2004. Mixotrophy in the phototrophic harmful alga Cochlodinium polykrikoides (Dinophycean): prey species, the effects of prey concentration, and grazing impact. J. Eukuryot. Microb., 51: 563−569.
Justic, D., N.N. Rabalais, R.E. Turner and Q. Dortch, 1995. Changes in nutrient structure of river-dominated coastal waters: stoichio- metric nutrient balance and its consequences. Estua. Coast. Shelf Sci., 40: 339−356.
Keller, M.D., R.C. Selvin, W. Claus and R.R..L. Guillard, 1987. Media for the culture of oceanic ultraphytoplankton. J. Phycol., 23: 633−
638.
Kim, D.I., T. Matsubara, S.J. Oh, Y. Shimasaki, Y. Oshima and T.
Honjo, 2007. Effects of nitrogen and phosphorus sources on the utilization and growth kinetics of the harmful dinoflagellate Cochlod- inium polykrikoides isolated from Yatsushiro Sea, Japan. Nippon Suisan Gakkaishi, 73: 711−717.
Fig. 8. A schematic diagram for interspecific competition in Jaran Bay.
「The Sea」, 19: 232−242.
Lehman, J.T., D.B. Botkin and G.E. Likens, 1975. The assumptions and rationales of a computer model of phytoplankton population dynamics. Limnol. Oceanogr., 20: 343−364.
Mann, K.H., 2000. Ecology of coastal waters: with implications for management. Wiley-Blackwell, pp. 432.
MIFAFF (Ministry of Food, Aquriculture, Forestry and Fisheries), 2008. Qnnual report of KSSP(Korea Shellfish Sanitation Program) for 2007. MIFAFF, pp. 3−5.
MLTM (Ministry of Land, Transportation and Marine Affairs, Korea), 2010. Standard methods for marine environmental analysis. Seoul, pp. 495.
Noh, I.H., Y.H. Yoon, J.S. Park, L.S. Kang, Y.K. An and S.H. Kim, 2010. Seasonal fluctuations of marine environment and phyto- plankton community in the southern part of Yeosu, southern sea of Korea. J. Kor. Soc. Mar. Environ. Eng., 13: 151−164.
Oh, S.J., 2002. DOP utilization by the two toxic dinoflagellates, Alexan- drium tamarense and Gymnodinium catenatum, and its advantage in species competition. Ph. D. Thesis, Hiroshima University, Hiro- shima, Japan, pp. 115.
Park, M.G. J.H. Shim and B.C. Cho, 1993. Adaptation of estuarine and
pp. 57−106.
Tarutani, K. and T. Yamamoto, 1994. Phosphate uptake and growth kinetics of Skeletonema costatum (Bacillariophyceae) isolated from Hiroshima bay (in Japan). J. Faculty of Applied Biological Sci- ence-Hiroshima University (Japan).
Tarutani, K. 1997. Ecophysiological studies on the population dynamics of toxic dinoflagellate Alexandrium tamarense. Ph.D. Thesis, University of Hiroshima, Higashi-Hiroshuma.
Tilman, D., 1981. Tests of resource competition theory using four spe- cies of Lake Michigan algae. Ecol., 62: 802−815.
Tilman, D., 1982. Resource Competition and Community Structure.
Princeton University Press, Princeton, pp. 296.
Yamamoto, T. and K. Tarutani, 1999. Growth and phosphate uptake kinetics of the toxic dinoflagellate Alexandrium tamarense from Hiroshima Bay in the Seto Inland Sea, Japan. Phycol. Res., 47:
27−32.
2015년 2월 5일 원고접수 2015년 3월 16일 수정본 접수 2015년 5월 11일 수정본 채택 담당편집위원: 조양기