관련 문서
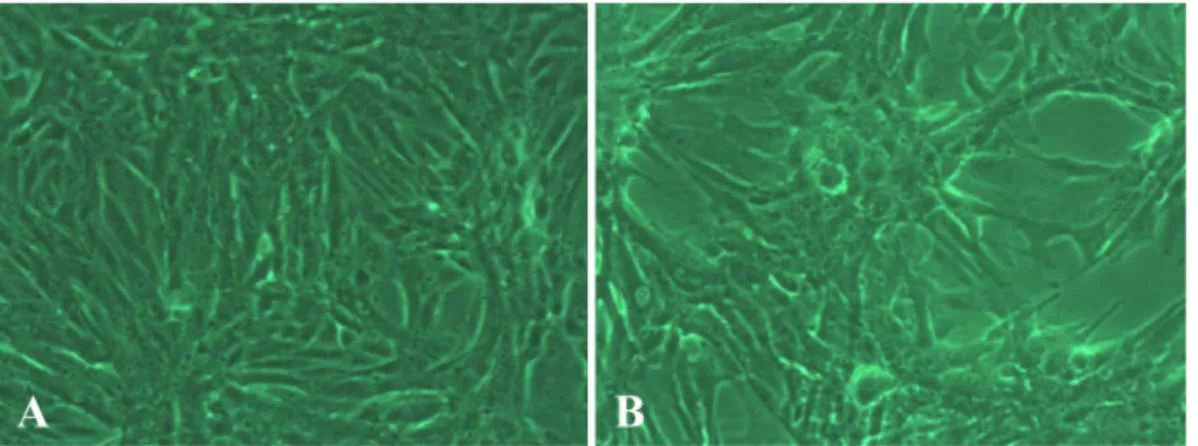
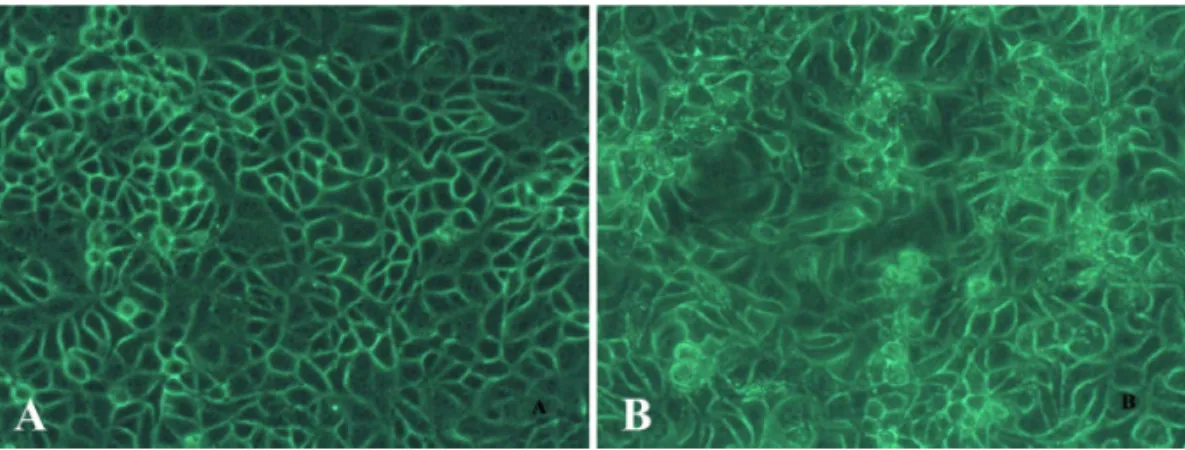
Results: The human breast cancer cell subline MCF-7/MX5 cells selected in the presence of 5 µg/ml mitoxantrone (MX) were more resistant to MX (15.7... Western blot and
In this study, two experiments were conducted to understand the effects of additional charge on the detailed growth mechanism of Alq 3 and to determine the effect of
Terms in exhibition panels on physics contents were extracted and they had their own grades according to the national curriculum and textbook glossary..
Comparison of genotypes using REP-PCR and Integron-IS26 PCR results among 78 IRAB strains isolated from 2 university hospitals.. The result of REP-PCR genotypes and
This study researched the possibility of using natural products for hair growth agents and hair loss prevention materials based on the test for the antioxidant and
Infection, Genetics and Evolution. gyrB Multiplex PCR To Differentiate between A cinetobacter calcoaceticus and Acientobacter Genomic Species 3.. Validation of
This study presented effective make-up techniques which help the subjects to change images as follows : good image were highlighted and bad images were
Diesel fuel spray makes aerosol form and the size of droplets is one of the important parameter that influences the combustion process.. The combustion of