서 론
어류의 산란장 및 성육장에서의 먹이생물의 양과 포 식자로부터의 도피 등은 자치어의 생존율을 좌우하는 주요요인으로 알려져 있다(Blaxter, 1984). 이러한 점에 서 한국 남서~서남부 해역은 먹이생물이 풍부한 다도 해로, 각종 어류의 산란장 및 성육장으로 이용되며, 특히
제주도 인근해역에서 춘~하계에 산란, 부화된 자치어들 이 완도연안까지 수송되어 성육하는 것으로 보고된 바 있다(Kim et al., 2002). 따라서, 이 지역 출현 자치어의 신속, 정확한 종동정을 위하여 성장에 따른 형태발달에 관한 상세 정보가 필요하며, 이는 조상의 형태, 유사분류 군과의 유연관계를 추정하고, 나아가 유어기의 생태적인 적응형태를 이해하는데 도움을 준다(Kim et al., 2000)
자치어에 대한 연구는1980년대 후반부터 초기생활사
─
─ 289 ──
완도연안에서 채집된 자치어 4종의 형태발달
김 진 구∙김 용 억*
국립수산과학원 남해수산연구소 목포분소, *부경대학교 해양생물학과
Morphological Development of the Four Fish Larvae and Juveniles Collected in Coastal Waters off Wando, Korea
Jin Koo Kim and Yong Uk Kim*
Mopko Laboratory, National Fisheries Research and Development Institute, Mokpo, 530-140, Korea
*Department of Marine Biology, Pukyong National University, Busan, 608-737, Korea
This paper presents morphological development and change in distribution of melanophores by developmental stage of four species caught by long net in coastal waters off Wando, Korea, from May to October, 1999.
Pennahia argentatus reached juvenile stage at 10.1 mm total length (8.8 mm standard length), and showed characteristics of the family Sciaenidae, including an increase in dorso- lateral melanophores, elongate caudal fins, etc. Gymnapogon urospilotus reached juvenile stage at 12.8 mm standard length, and it seemed unlikely to begin to differentiate the pelvic fins. The species is similar to G. japonicus, but the two differed in the number of pectoral fin rays (12 in G. japonicus c.f. 14~15 in G. urospilotus). It is an unrecorded species of the Korean fish fauna, so we provide a Korean name, “Kko-ri-jeom-min-dong-gal-dom”. Platycephalus sp. reached juvenile stage at 8.2 mm total length (7.1 mm standard length). It showed a tendency towards increased melanophores along the lateral side of the head and dorso- laterally on the body, and to develop the neural spine with growth. At 11.6 mm standard length Hapalogeny mucronatus is distinguished from Hapalogeny nitens and Branchiostegus japonicus having pelvic fins very long and the 10 preopercle spines very sharp.
Key words : Pennahia argentatus, Gymnapogon urospilotus, Hapalogeny mucronatus, Platycephalus sp. morphological development, unrecorded species
와 분포상 연구로 시작하여, 최근에는 광양만 부유성 난, 자치어의 종조성 및 초기 섭식생태(Park, 1999), 자 연산 및 인공산 조피볼락, Sebastes schlegeli종묘의 생 물학적 특성 비교(Park, 1999), 대한해협의 수괴 구조에 따른 자치어의 분포 특성(Kim, 1999) 등 다양한 분야에 서 연구가 진행되고 있지만, 자치어의 빈약한 형태 정보 는 이들의 가입기작에 대한 연구를 어렵게 만들고 있다. 따라서, 본 연구의 목적은 한국 남서부 해역에 위치한 완도연안에서 채집한 자치어 4종을 대상으로 발육단계 에 따른 형태발달 및 체색변화를 상세히 기술하고자 한 다.
재료 및 방법
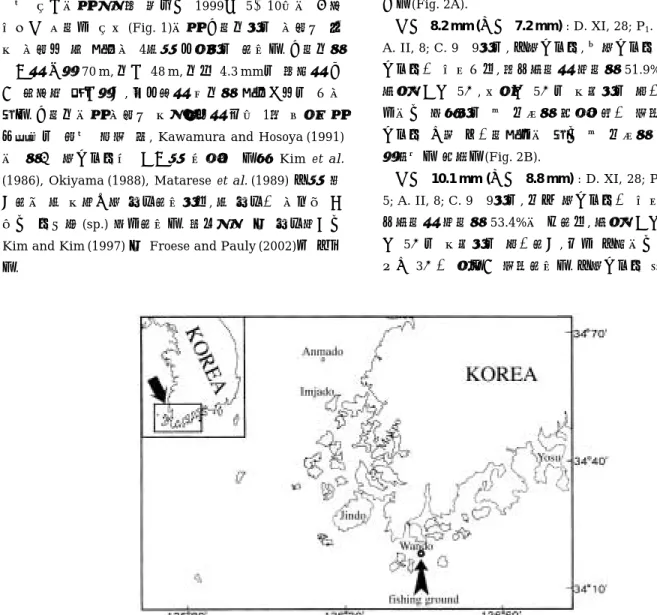
본 연구에서 사용한 재료는 1999년 5~10월에 걸쳐 완도군 석장리 연안(Fig. 1)에서 낭장망으로 어획된 부 수 어획물 중 자치어 4종을 대상으로 하였다. 낭장망의 규격은 길이 70 m, 망구48 m, 망목4.3 mm로 한쪽은 넓 게 펼쳐진 입구이고, 반대편은 세망의 자루그물로 되어 있다. 낭장망에서 어획된 수산생물은 매월 1회 선상에서 무작위로 표본 추출 후, Kawamura and Hosoya (1991) 에 의거 지느러미와 골격을 염색한 다음 Kim et al.
(1986), Okiyama (1988), Matarese et al. (1989) 등을 참 고하여 종 수준까지 분류하였으며, 종 분류가 어려운 경 우는 미동정(sp.) 처리하였다. 학명사용 및 분류체계는 Kim and Kim (1997) 및 Froese and Pauly (2002)를 따랐 다.
결 과
1. 보구치, Pennahia argentatus (Houttuyn, 1782)
전장 7.3 mm (체장 6.4 mm): D. IX, 25; P1. 6; P2. 4; A.
II, 8; C. 8±7으로, 뒷지느러미만 완성되어 있었다. 머리 는 비대하고 체고는 머리 뒷가장자리에서 가장 높았다. 항문전장은 체장의 50.0%에 달하며, 꼬리지느러미는 장 타원형태를 나타내었다. 눈은 크고, 눈의 윗가장자리를 따라 3개의 안상극이 출현하며, 전새개골극 4개, 주새개 골극1개 나타났다. 입은 경사지며, 윗턱의 뒷가장자리는 동공의 중앙 아래를 지나지만 눈의 뒷가장자리에는 못 미쳤다. 머리에는 흑색소포가 없지만 복강과 뒷지느러미 기저말단 부위에 미약하게 별 모양의 흑색소포가 나타 났다(Fig. 2A).
전장 8.2 mm (체장 7.2 mm): D. XI, 28; P1. 14; P2. I, 5;
A. II, 8; C. 9±9으로, 등지느러미, 배지느러미 및 꼬리지 느러미가 완성되며, 항문전장은 체장의51.9%에 달하고, 전새개골극 5개, 안상극 5개로 수적으로 증가하였다. 머 리에는 처음으로 별 모양의 흑색소포가 출현하며, 뒷지 느러미 기저 뒷가장자리에 있는 별 모양의 흑색소포가 이전보다 커졌다(Fig. 2B).
전장 10.1 mm (체장 8.8 mm): D. XI, 28; P1. 18; P2. I, 5; A. II, 8; C. 9±9으로, 모든 지느러미가 완성되었다. 항 문전장은 체장의53.4%에 달하며, 전새개골극 5개, 안상 극 5개로 수적으로 증가하고, 머리 등쪽에는 볏 모양의 돌기3개가 새롭게 출현하였다. 등지느러미 앞쪽과 가슴
Fig. 1. Map showing the sampling area in the coastal waters off Wando, Korea.
지느러미 위쪽에 별 모양의 흑색소포가 새롭게 출현하 였다(Fig. 2C).
전장 15.6 mm (체장 12.6 mm) : 항문전장은 체장의
57.9%로 시간의 경과와 함께 항문이 몸의 뒷쪽으로 이
동하며, 꼬리지느러미는 길고 끝이 뾰죽해졌다. 전새개골 극 6개, 주새개골극 2개, 안상극 8개로 수적으로 증가하 고, 하새개골극 3개, 간새개골극 4개, 제1안하극 6개가 새로이 출현하였다. 체측 중앙과 등쪽을 따라 점 모양 및 별 모양의 흑색소포가 1열로 간헐적으로 출현하고, 머리 등쪽 흑색소포는 수적으로 증가하는 한편 꼬리지 느러미 앞쪽으로 점 모양의 흑색소포가 나타났다(Fig.
2D).
전장 17.4 mm (체장 13.8 mm) : 항문전장은 체장의 55%로 항문은 몸의 중앙보다 약간 뒷쪽에 위치하였다.
뺨 부위에 별 모양의 흑색소포가 새롭게 출현하며, 체측 중앙 및 등쪽에 있는 점 모양의 흑색소포가 별 모양으 로 바뀌고, 꼬리지느러미 기저에 띠 모양의 흑색소포가 새롭게 출현하였다(Fig. 2E).
부기 : 본종은 과거 Argyrosomus속에 포함되었지만, Sasaki and Kailola (1988)에 이르러 Pennahia속으로 정 리되었으며 이 후 Froese and Pauly (2002)가 이를 받아 들였다.
2. 꼬리점민동갈돔 (국명신칭), Gymnapogon urospilotus Lachner, 1953 (New Korean name : Kko-ri-jeom-min-dong-gal-dom)
체장 3.9 mm: P2. I, 5으로 배지느러미는 완성되어 있 고, 척색 말단은 비스듬히 경사져 있었다. 항문전장은 체 장의 50.4%, 근절 높이는 체장의 10.1%, 배지느러미 길 이는 체장의22.9%로 배지느러미는 길게 신장되어 뒷끝 이 항문에 달하였다. 배지느러미에는 나뭇가지 줄기 모 양의 흑색소포가 조밀하게 분포하며, 몸 중앙의 체측 등 쪽과 중앙은 점 모양, 배쪽은 나뭇가지 줄기 모양의 흑 색소포가 분포하였다(Fig. 3A).
체장 6.3 mm: D. IV-I, 10; P1. 9; P2. I, 5; A. I, 6; C.
5±6으로, 제2등지느러미와 배지느러미만 완성되어 있 었다. 입은 크며, 경사지고 뒷끝이 동공의 중앙 아래까지 달하였다. 항문전장은 체장의 60.3%, 체고는 체장의
30.4%, 배지느러미는 체장의 35.1%로 배지느러미는 매
우 길게 신장되어 뒷끝이 뒷지느러미 기부를 훨씬 지났 다. 안후부와 머리 등쪽 부위에 미약하게 흑색소포가 출 현하며, 제2등지느러미와 뒷지느러미 사이 및 배지느러 미에는 나뭇가지 모양의 흑색소포가 조밀하게 분포하였 다(Fig. 3B).
체장 12.8 mm: D. VI-I, 10; P1. 15; P2. I, 5; A. II, 9; C.
9±8으로, 이 시기에 모든 지느러미가 완성되었다. 윗턱 의 뒷끝은 동공의 뒷가장자리 아래까지 달하며, 양턱에 는 1열로 날카로운 송곳니가 나 있었다. 전새개골의 뒷 가장자리에는 2개의 극이 있고, 아래쪽 극은 끝이 2갈래 로 분지되어 있었다. 항문전장은 체장의 59.0%, 체고는 체장의 24.9%, 배지느러미는 체장의 29.7%이었다. 눈의 앞쪽으로 흑색소포가 새롭게 출현하며, 머리 등쪽과 안 후부의 흑색소포는 수적으로 증가하였다. 또한, 제2등지 느러미와 뒷지느러미 사이의 흑색소포는 일부가 나뭇가 지 모양에서 점 모양으로 바뀌며, 수적으로는 증가하였 다(Fig. 3C).
체장 16.1 mm: 항문전장은 체장의 60.0%, 체고는 체
장의 25.7%, 배지느러미는 체장의 25.5%으로 배지느러
Fig. 2. Development of larvae and juveniles of Pennahia argentatus. A: 6.4 mm in standard length (SL); B:
7.2 mm SL; C: 8.8 mm SL; D: 12.6 mm; E: 13.8 mm.
A
B
C
D
E
미는 체장의 증가와 함께 감소경향을 나타내었다. 주새 개골에 커다란 나뭇가지 모양의 흑색소포가 새롭게 출 현하며, 제2등지느러미와 뒷지느러미 사이의 흑색소포는 작은 점 모양으로 바뀌면서 점차 희미해졌다(Fig. 3D).
부기: Kim and Lee (1994)가 제주도에서 채집한Gym- napogon japonicus를 미기록종으로 보고하면서 Gym- napogon속의 국명을 “민동갈돔속”으로 제창한 이후 지 금까지 국내에는 민동갈돔 1종만이 보고되어 있었다. 본 종은 민동갈돔과 형태 및 계수형질에서 유사하지만, 가 슴지느러미수(민동갈돔은 12개인 반면, 본 종은 14~15 개를 가짐)에서 잘 식별된다. 본 종의 국명으로는 성어 기에 꼬리지느러미 기저에B모양의 독특한 점을 가지는 점에서“꼬리점민동갈돔”으로 신칭한다.
3. 양태속 어류, Platycephalus sp.
전장 6.4 mm (체장 5.6 mm): D. II-V-I, 12; P1. 15; P2. I, 5; A. 9으로, 배지느러미만 완성되어 있고 나머지 지느 러미는 분화 중이었다. 입은 크며, 경사지고 윗턱의 뒷끝 이 동공의 앞가장자리 아래까지 뻗어 있었다. 항문전장 은 체장의 67.5%, 체고는 26.7%, 두장은 44.8%이며, 가 슴지느러미와 배지느러미가 매우 길어 뒷가장자리가 뒷 지느러미 기부를 지나고 있었다. 머리에는 발달된 안상
극과 두정극이 나타나고, 두정극의 앞가장자리는 톱니 모양이며, 전새개골의 뒷가장자리에는 4개의 날카로운 극이 존재하였다. 머리에는 전새개골과 눈의 앞쪽 부위 에 나뭇가지 모양 혹은 점 모양의 흑색소포가 미약하게 출현하며, 등지느러미 연조 기부 아래쪽과 꼬리지느러미 기저에 국한되어 별 모양의 흑색소포가 나타났다. 지느 러미에는 흑색소포가 거의 없지만, 가슴지느러미 상반부 에만 별 모양의 흑색소포가 조밀하게 분포하는 것이 특 징적이었다(Fig. 4A).
전장 8.2 mm (체장 7.1 mm): D. II-VII-I, 13; P1. 18;
P2. I, 5; A. 13으로, 이 시기에 모든 지느러미가 완성되었 다. 아래턱이 머리의 앞끝에 위치하며, 윗턱의 뒷끝은 동 공의 중앙 아래까지 뻗어 있었다. 머리에는 다양한 머리 부분극이 출현하기 시작하는데, 익이극, 주새새극 등이 새롭게 출현하며, 항문전장은 체장의 64.5%, 체고는 24.4%, 두장은 41.7%이었다. 머리에는 새개부에 나뭇가 지 모양 및 별 모양의 흑색소포가 수적으로 증가하여 머리를 가로지르는 띠처럼 보이고, 꼬리지느러미 기저의 흑색소포는 별 모양에서 점 모양으로 바뀌었다(Fig.
4B).
전장 9.8 mm (체장 8.8 mm): 아래턱이 머리의 앞끝에 위치하며, 윗턱의 뒷끝은 동공의 뒷가장자리 아래에 달 한다. 가슴지느러미를 지지하는 견대부의 쇄극과 후측두 극 및 안부의 안하극이 새롭게 출현하고, 등지느러미 및 배지느러미 극이 커지는데 특히, 등지느러미 2번째 가시 는 두꺼워지는 반면 3번째 가시가 길게 신장되었다. 항 문전장은 체장의 61.4%, 체고는 21.1%, 두장은 44.5%이 었다. 두정부에 점 모양의 흑색소포가, 등지느러미 극조 부의 기저에 1열로 별 모양의 흑색소포가 새롭게 출현 하며, 등지느러미 연조부 아래쪽의 흑색소포는 커지고 수적으로 증가하여 몸의 중앙부위가 어둡게 보였다(Fig.
4C).
전장 11.9 mm (체장 10.2 mm): 양태과의 특징적인 머 리부분극이 시간의 경과와 함께 더욱 날카로와지고, 전 새개골의 극은5개로 증가하며 안하극은 외형이 뚜렷해 졌다. 항문전장은 체장의 65.8%, 체고는 18.6%, 두장은
41.5%으로, 체고가 지속적으로 감소하는 경향을 나타내
었는데, 이로써 양태속이 후기자어기부터 몸이 종편되는 것으로 생각된다. 몸통 중앙부위의 흑색소포는 앞쪽으로 점차 확산되면서 전체적으로 어둡게 보이며 등지느러미 연조부의 중앙 아래의 몸통 뒤쪽으로는 전혀 흑색소포 가 없고 단지 꼬리지느러미 기저에만4~5개의 점 모양 의 흑색소포가 나타났다(Fig. 4D).
부기 : Lee and Joo (1998)는 한국산 양태과 어류의 분 류학적 재검토에서 양태속, Platycephalus 어류에 양태, Fig. 3. Development of larvae and juveniles of Gymnapo-
gon urospilotus. A: 3.9 mm SL; B: 6.3 mm SL; C:
12.8 mm SL; D: 16.1 mm SL.
A
B
C
D
P. indicus및 미기재종 P. sp의 2종을 보고하였으며, 이 후 Lee et al. (1999)이 근단백질 및 isozyme 분석결과 P.
sp.가 양태와는 별종임을 재확인하였다. 본 조사에서 양 태속 자치어는 상기 2종의 분류형질인 측선비늘수의 계 수가 어렵고, 안경, 양안간격 등 계측이 자치어기에는 적 용하기 어려운 점을 감안하여 미동정종으로 처리하였다.
4. 군평선이, Hapalogeny mucronatus (Eydoux and Souleyet, 1850)
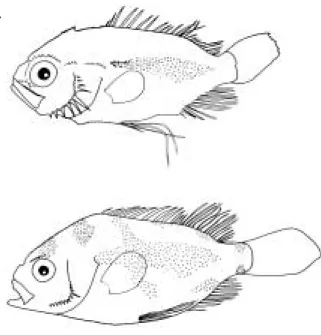
전장 11.6 mm (체장 9.7 mm): D. XI, 15; P1. 19; P2. I, 5; A. III, 9으로, 모든 지느러미가 완성되어 있었다. 입은 경사지고 윗턱의 뒷끝이 동공의 중앙 아래까지 뻗어 있 었다. 체표면에는 털 모양의 매우 미세한 가시가 나 있 고, 안상극 10개, 안하극 3개, 익상극 3개로 머리부분에 는 크고 작은 많은 극이 출현하였다. 또한, 머리 상단부 의 등쪽면에는 톱니 모양의 커다란 돌기가 나 있고, 전 새개골의 뒷가장자리에는 10개의 날카로운 극이 나 있 었다. 배지느러미는 매우 길어 뒷끝이 뒷지느러미의 기 부를 훨씬 지나며, 머리에는 흑색소포가 없고, 체측 중앙 에만 점 모양의 흑색소포가 분포하여 좁은 세로띠 모양 을 나타내었다(Fig. 5A).
전장 23.5 mm (체장 19.1 mm): 머리부분의 날카로운 극은 성장하면서 작아지고, 안상극은 7개로 줄어든 반
면, 익상극 5개, 전새개골극 17개로 증가하였다. 흑색소 포는 눈의 등쪽, 머리의 등쪽 및 체측의 등쪽과 배쪽면 에 새롭게 출현하며 성어의 체색과 유사해졌다(Fig.
5B).
부기 : 체표면을 덮고 있는 미세한 가시는 옥돔, Bran- chiostegus japonicus자치어와 유사하지만, 옥돔은 본 종 에 비하면 배지느러미가 짧다. 또한, 머리의 상단부에 있 는 볏모양의 돌기는 동갈돗돔, Hapalogenys nitens과 유 사하지만, 동갈돗돔은6개의 전새개골극 및 짧은 배지느 러미를 가지는 점에서 본종과 잘 구별된다.
고 찰
본 연구는 우리 나라 남서해역에 위치한 완도연안에 서 1999년 5~10월 사이에 출현한 주요 자치어 4종의 발육단계별 형태발달 및 체색변화 과정을 검토하였다.
자치어는 성어와 다른 형태 및 체색을 지니기 때문에, 골격발달과 계수형질(Ahlstrom and Moser, 1981), 계측 과 색소포(Russell, 1976) 등이 중요한 자치어기의 분류 형질로 간주되며, 본 연구에서는 계수형질, 계측형질, 머 리부분극의 형태발달, 흑색소포의 형태와 수적 변화 등 을 중점적으로 조사하였다.
본 조사에서 보구치 자치어는 전장 7.3~17.4 mm으 로, 성장하면서 꼬리지느러미가 둥근 형태에서 뾰죽하고 긴 형태로 변형되는데 이는 조류가 강한 지역에서 적응 Fig. 4. Development of larvae and juveniles of Platyceph-
alus sp. A: 5.6 mm SL; B: 7.1 mm SL; C: 8.8 mm SL; D: 10.2 mm SL.
Fig. 5. Development of juveniles of Hapalogenys mucro- natus. A: 9.7 mm SL; B: 19.1 mm SL.
A
B
C
D
A
B
하기 위한 것으로 생각된다. 보구치의 주산란기는 6~7 월, 생물학적 최소체장은 17.38 cm (2.46세)로(Kang et al., 1999), 본 조사에서 채집된 보구치는 당해연도 7월 산란개체로 추정된다. 보구치는 최근 자원량의 현저한 감소로 어획개시연령을 1.12세에서 2세로 조정하는 방 책이 거론되고 있지만(Zhang et al., 1999), 성육장에서의 가입 이전의 잠재자원에 대한 보호 대책이 수립되어야 할 것이다.
꼬리점민동갈돔의 자치어기에서 볼 수 있는 배지느러 미의 비정상적인 분화, 즉 배지느러미가 가장 빨리 분화 되는 점과 비정상적으로 길어지는 점은 민동갈돔속 어 류와는 흡사하지만, 동갈돔속(Apogon) 어류와는 잘 구별 된다(Okiyama, 1988). 성어기의 꼬리점민동갈돔은 꼬리 지느러미 기저에 B모양의 점을 가져, 동갈돔과의 다른 속과는 잘 구별되지만, 본 조사에서는 체장 16.1 mm 개 체까지 B모양의 점이 관찰되지 않아 자치어기의 식별형 질로는 타당하지 않은 것으로 생각된다. 자치어기에 지느 러미가 특화되는 현상은 동갈돔과(Apogonidae), 바리과 (Serranidae), 퉁돔과(Lutjanidae), 양쥐돔과(Acanthuri- dae), 갈치꼬치과(Gempylidae), 갈치과(Trichiuridae), 둥 글넙치과(Bothidae), 넙치과(Paralichthyidae), 참서대과 (Cynoglossidae) 등의 종에서 볼 수 있다(American So- ciety of Ichthyologists and Herpetologists, 1984).
양태속 자치어는 전장 5.6~10.2 mm로, 머리부분 골 질돌기의 발달과 가슴지느러미 및 배지느러미가 길게 발달되는 점이 특징적이다. 양태속의 분류학적 연구에서 Lee and Joo (1998), Lee et al. (1999)은 처음으로 한국산 양태속 어류에1신종 포함2종이 서식하는 것으로 보고 하였지만, 2종의 분류형질인 측선린수, 안경, 양안간격 등 계수, 계측형질을 자치어기와 비교하기 곤란한 점 때 문에 미동정종으로 처리하였다. 따라서, 완도 연안 출현 양태속 자치어의 정확한 분류를 위해서는 향후 자치어를 대상으로 한 단백질 전기영동 등의 조사가 필요하겠다.
군평선이는 11.6~23.5 mm의 치어 및 유어가 출현하 였는데, 전새개골극이 분화되어 있고, 배지느러미가 길어 진 형태적 특징을 띤다. 군평선이 치어는 몸 전체에 털 모양의 미세한 가시를 가지는 점이 특징적인데, 이러한 형태적 특징은 옥돔과(Branchiostegidae)의 자치어와 유사하지만, 옥돔과 자치어는 배지느러미가 길지 않아 잘 구별된다(Okiyama, 1988).
자치어에 대한 연구는 초기에 산란군 형성, 자어의 이 동 및 수평∙수직 분포에 대한 조사에 초점이 맞추어 졌지만, 정량 샘플링이 가능해짐에 따라 산란군 크기의 추정, 자원-가입 관계의 추정 등 개체군의 평가 및 다양 한 해양 가설을 검증하기 위한 실질적인 연구로 방향이
전환되고 있다(Blaxter, 1984). 우리 나라에서는 현재까 지 자치어 형태발달, 해역별 종조성 및 분포 등을 중심 으로 연구가 진행되어 왔고, 일부 자원연구에 적용된 바 있지만 대상종이 한정되어 있으며, 치어 및 가입 이전의 미성어는 연구 대상에서 제외되어, 수산자원으로의 가입 기작에 대한 연구가 미비한 상황이다. 이처럼 자연 그대 로를 대표할 수 있는 채집방법에 대한 연구가 급선무인 데, Cha (2002)는 주간의 상층(표층) 채집시 출현종수의 감소 및 출현량의 과소 평가를 우려하였고, 저자가 자치 어의 주야간 수직이동을 조사한 바에 의하면, 어란은 표 층에서는 주야간 차이를 나타내었지만 중층에서는 주야 간 차이가 없었고, 자치어는 표~중층에서 모두 주야간 차이를 나타내었다. 따라서, 자치어의 현존량을 정확하게 파악하기 위하여 수층별 채집이 이루어져야 할 것으로 생각된다.
적 요
본 논문에서는 한국 남해 서부해역에 위치한 완도에 서 1999년 5~10월에 낭장망으로 어획된 자치어 4종의 발육에 따른 형태발달 및 흑색소포 변화를 나타내었다.
보구치는 전장 10.1 mm (체장 8.8 mm)에서 치어기에 도달하였고, 발육하면서 체측 흑색소포가 증가하며 꼬리 지느러미가 길게 신장되는 등 전형적인 민어과의 특징 을 나타내었다.
꼬리점민동갈돔은 체장 12.8 mm에서 치어기에 도달 하며, 배지느러미부터 분화되기 시작한다는 점이 특이해 보인다. 본종은 민동갈돔과 유사하지만 가슴지느러미 줄 기수(민동갈돔은 12개, 본 종은 14~15개)에서 잘 구별 되며, 우리나라에서는 처음으로 보고되는 미기록종으로
“꼬리점민동갈돔”으로 국명을 부여하였다.
양태속은 전장 8.2 mm (체장 7.1 mm)에서 치어기에 도달하며, 발육하면서 머리부분극은 더욱 발달하고, 머리 의 측면과 몸의 등쪽, 측면을 따라 흑색소포가 증가하는 경향을 나타내었다.
군평선이는 전장 11.6 mm개체에서 매우 긴 배지느러 미와 10개의 날카로운 전새개골극을 가지는 점에서 옥 돔 및 동갈돗돔과 잘 구별되었다.
사 사
본 연구는 국립수산과학원 경상시험연구비로 수행된 결과의 일부이며, 연구비 지원에 감사드립니다. 또한, 시
료 분석 및 스케치에 협조하여 주신 목포대학교 나종태 군, 목포해양대학교 정수동군에게 감사드립니다.
인 용 문 헌
Ahlstrom, E.H. and H.G. Moser. 1981. Systematics and development of early life history stages of marine fish- es: achievements during the past century, present sta- tus and suggestioins for the future. Rapp. P. V. Reun.
Cons. Int. Explor. Mer, 178 : 541~546.
American Society of Ichthyologists and Herpetologists.
1984. Ontogeny and systematics of fishes. Special Publication Number 1. Allen Press INC., Lawrence, 760 pp.
Blaxter, J.H.S. 1984. Ontogeny, systematics and fisheries.
pp. 1~6 in H.G. Moser, editor. Ontogeny and Systema- tics of Fishes. The American Society of Ichthyologists and Herpetologists, Lawrence.
Cha, S.S. 2002. Review on the studies of ecology of fish in their early life stages off Korea. Korean J. Ichthyol., 14 (1) : 76~82. (in Korean)
Froese, R. and D. Pauly. Editors. 2002. FishBase. World Wide Web electronic publication. www.fishbase.org, 18 September, 2002.
Kang, Y.J., D.H. Kwon and C.I. Zhang. 1999. A population ecological study of white croaker, Argyrosomus argen- tatus Houttuyn in Korean waters III. Maturation and spawning. J. Korean Soc. Fish. Res., 2 : 61~67. (in Korean)
Kawamura, K. and K. Hosoya. 1991. A modified double staining technique for making a transparent fish-ske- letal specimen. Bull. Natl. Res. Inst. Aquaculture, 20 : 11~18. (in Japanese)
Kim, I.S. and W.O. Lee. 1994. New records of seven species of the order Perciformes from Cheju Island, Korea.
Korean J. Ichthyol., 6(1) : 7~20.
Kim, J.K., O.I. Choi, D.S. Chang and J.I. Kim. 2002.
Fluctuation of bag-net catches off Wando, Korea and the effect of sea water temperature. J. Korean Fish.
Soc., 35(5) : 497~503. (in Korean)
Kim, J.K., Y.U. Kim and S.G. Byun. 2000. Egg and larval development of Chelon lauvergnii from Korea. Korean
J. Ichthyol., 12(2) : 137~145. (in Korean)
Kim, J.M., J.M. Yoo, J.K. Myoung and J.Y. Lim. 1986.
Guides to the early stages of marine fishes in the Korean waters. KORDI Report, BSPE00060-98-3, 369 pp. (in Korean)
Kim, S. 1999. Distribution of the fish larvae in relation to the water masses in the Korea strait. Ph. D. Thesis, University of Inha, 128 pp. (in Korean)
Kim, Y.U. and I.S. Kim. 1997. Pisces. Pages 243~281. In H.S. Kim, editor. List of animals in Korea (excluding insects). The Korean Society of Systematic Zoology, Seoul. (in Korean)
Lee, C.L. and D.S. Joo. 1998. Taxonomic review of flathead fishes (Platycephalidae, Scorpaeniformes) from Korea.
Korean J. Ichthyol., 10(2) : 216~230. (in Korean) Lee, C.L., J.H. Kim and C.S. Kim. 1999. Taxonomic review
of the genus Platycephalus (Platycephalidae) from Korea. Korean J. Ichthyol., 11(2) : 143~148. (in Korean) Matarese, A.C., A.W. Kendall, D.M. Blood and B.M. Vinter.
1989. Laboratory Guide to Early Life History Stages of Northeast Pacific Fishes. NOAA Technical Report NMFS 80, 625 pp.
Okiyama, M. 1988. An Atlas of the Early Stage Fishes in Japan. Tokai Univ. Press., 1154 pp. (in Japanese) Park, K.J. 1999. Species composition of the ichthyoplank-
ton and feeding ecology of early stage in Kwangyang Bay, Korea. Ph. D. Thesis, University of Chonnam, 131 pp. (in Korean)
Park, M.W. 1999. Comparison of the biological characteri- stics between wild and cultured juvenile black rockfish, Sebastes schlegeli. Ph. D. Thesis, University of Inje, 89 pp. (in Korean)
Russell, F.S. 1976. The eggs and planktonic stages of British marine fishes. Academic Press, London, pp.
20~35.
Sasaki, K. and P.J. Kailola. 1988. Three new Indo-Aus- tralian species of the sciaenid genus Atrobucca, with a reevaluation of generic limit. Japan. J. Ichthyol., 35(3) : 261~277.
Zhang, C.I., Y.J. Kang and M.W. Lee. 1999. A population ecological study of white croaker, Argyrosomus argen- tatus Houttuyn in Korean waters IV. Population ecolo- gical characteristics and biomass. J. Korean Soc. Fish.
Res., 2 : 68~76. (in Korean)
Received : November 3, 2002 Accepted : December 20, 2002