벼 등숙일수에 따른 이삭 착생 부위별 종자의 수발아 변이
백정선 ・ 정남진†
전북대학교 농업생명과학대학 작물생명과학과
Pre-Harvest Sprouting Variation of Rice Seeds Located on Each Panicle Position According to Grain Filling Days
Jung-Sun Baek and Nam-Jin Chung†
Department of Crop Science and Biotechnology, Chonbuk National University, Jeonju 561-756, Republic of Korea
22
†
Corresponding author: (Phone) +82-63-270-2512 (E-mail) [email protected]
<Received 30 April, 2013; Revised 30 September, 2013; Accepted 6 February, 2014>
한작지(Korean J. Crop Sci.), 59(1): 22~26(2014) DOI : http://dx.doi.org/10.7740/kjcs.2014.59.1.022
ABSTRACT This research was implemented to investigate pre-harvest sprouting characteristics in two Korean rice cultivars, Shindongjin (SDJ) and Hopum (HP). The panicles of both varieties were sampled after 15 days after heading (DAH) to 60 DAH at intervals of 5 days. As soon as sampled, the panicles were imbibed in water for 14 days at 25℃ to determine the vigor and germination ability of seeds according to location on panicle. To investigate the cause of non-germination of seeds in panicles, tetrazolium test and dormancy breaking were performed. The pre- harvest sprouting of HP started at 20 DAH while that of SDJ began at 30 DAH. The germination of seeds located in high rank branches within a panicle and the terminal seeds within a branch were earlier and faster and the germination patterns were same in both cultivars. The times at more than 50% of germination in a panicle were 35 DAH (57.0%) in HP and 45 DAH (56.8%) in SDJ. Pre- harvest sprouting was more than 80% at 50 DAH (82.6%
of HP, 92.3% of SDJ) and more than 99% at 60 DAH (99.5%, 99.4%, HP and SDJ). These results indicated that the rate of PHS in a panicle increased with accumulation of the days after heading. The cause of non-germinated seeds at 15-25 DAH in panicle was immature embryo.
After 30 DAH, however, when the non-germinated seeds were hulled, they started to germinate due to dormancy breaking, in which the germination percentage was higher in SDJ than HP. In conclusion, the pre-harvest sprouting varied according to days after heading, the seed position on panicle, and the dormancy intensity of varieties.
Keywords : rice, seed, pre-harvest sprouting, dormancy
우리나라와 같은 온대지역에서는 벼 등숙기에 태풍이나 강우로 인해여 재해를 입는 경우가 자주 발생한다(Lee et al., 2006). 2012년 태풍 “볼라벤”에 이어 북상한 “덴빈”으 로 인해 우리나라에서는 15,139 ha에 걸쳐 벼가 도복되거나 침수되는 피해가 발생하였다. 2010년에는 태풍 “곤파스”로 인해 쌀 생산량이 전년대비 12.6%, 평년대비 9.8%로 크게 감소되었다(Korean Rural Economic Institute, 2012). 이처 럼 벼 등숙기에 발생한 태풍이나 강우는 벼 도복이나 침수 와 같은 피해를 불러올 뿐만 아니라, 이삭에 달려있는 상태 로 종자의 발아를 유발하는 수발아(Pre-harvest sprouting) 를 발생시켜 쌀의 품질과 생산량에 영향을 미치기도 한다 (Ko et al., 2005; Kim et al., 2008).
그 동안 종자의 수발아로 인한 피해는 다양한 연구에 의 해 밝혀져 왔다. 수발아는 종자의 전분과 단백질이 가수분 해 된 상태이기 때문에 종자 활력을 감소시킨다. 뿐만 아니 라 각종 세균들의 감염이 용이하여 종자부패의 원인이 되기 도 한다(Biddulph et al., 2008; Gubler et al., 2005; Huang et al., 2012; Rasul et al., 2012). 또한, 왕겨 밖으로 유근이 나 유아가 돌출되지 않는 정도의 수발아가 진행되어도 저장 중에 곰팡이와 같은 미생물이 번식이 조장되어 종자의 수명 및 저장기간을 단축시키기도 한다(Li et al., 2004).
한편, 벼의 수발아는 강우나 안개, 강풍이나 집중호우 등 의 다양한 습윤 환경의 영향을 받으며(Park and Kim, 2009), 재배시기, 수확시기, 일장, 온도 등과 같은 외부적 요 인들도 종자의 수발아에 영향을 미친다. 종자의 수발아는 이 같은 외부적 요인뿐만 아니라 종자의 휴면, 종피의 투수 성, α-amylase 활성, 내적 호르몬의 수준 등 종자의 내적 요
Table 1. Meteorological data from heading to harvest of rice in 2012.
month days Daily mean temperature (°C) Precipitation
average maximum minimum amount (mm) number of days
Aug. middle 28.0 32.1 24.6 103.5 5
last 25.6 30.1 22.3 163.5 8
Sep.
early 23.2 28.3 18.9 109.0 6
middle 20.7 24.5 17.8 100.0 4
last 19.6 26.0 14.5 0.1 1
Oct. early 17.8 24.7 12.2 0.1 1
middle 14.2 22.0 7.8 1.5 1
벼에서 출수 후 일수에 따른 수발아 양상을 조사하고, 수발 아 피해가 발생할 수 있는 시기를 검토함으로써 수발아 방 제를 위한 기초자료를 얻고자 하였다.
재료 및 방법
본 실험에 사용한 벼 품종은 호품벼와 신동진벼로 농촌진 흥청 국립식량과학원으로부터 분양 받아 전북대학교 실험 농장에서 30일간 육묘하여 6월 1일에 기계이앙하여 재배하 였으며, 벼 재배관리는 농촌진흥청 표준재배법에 의하였다.
벼의 출수기는 두 품종 모두 8월 15일이었고 10월19에 수확하였다. 출수기부터 수확기까지의 평균온도는 21.0℃, 강수량은 총 477.7 mm 이었다. 강수는 출수기부터 출수 후 34일(9월 17일)까지 집중되었고, 그 이후부터 수확기까지의 강수량은 1.7 mm였다(Table 1).
벼 출수 일수에 따른 수발아율 검정
벼 출수 후 일수에 따른 수발아율과 수발아 종자의 이삭 내 착생 위치를 조사 하기 위하여 출수 후 15일 후인 8월 31일부터 10월 15일까지 5일 간격으로 10 이삭씩 채취하여 수발아율을 측정하였다. 수발아는 항온항습기(25℃) 내에
되는 왕겨를 제거하여 발아율을 검정하였다. Tetrazolium 염색은 수분이 흡수된 미발아종자를 면도날을 이용하여 배 (embryo)가 포함되도록 세로로 절단하여 1% tetrazolium 용액(pH 6.5-7.5)에 30℃에서 2시간 동안 처리하였고, 해부 현미경을 통하여 배의 발달정도를 관찰 하였다. 왕겨를 제 거한 종자는 14일 동안 25℃의 항온항습기 내에서 발아율 을 측정하였다.
결과 및 고찰 벼 출수 일수에 따른 발아력 검정
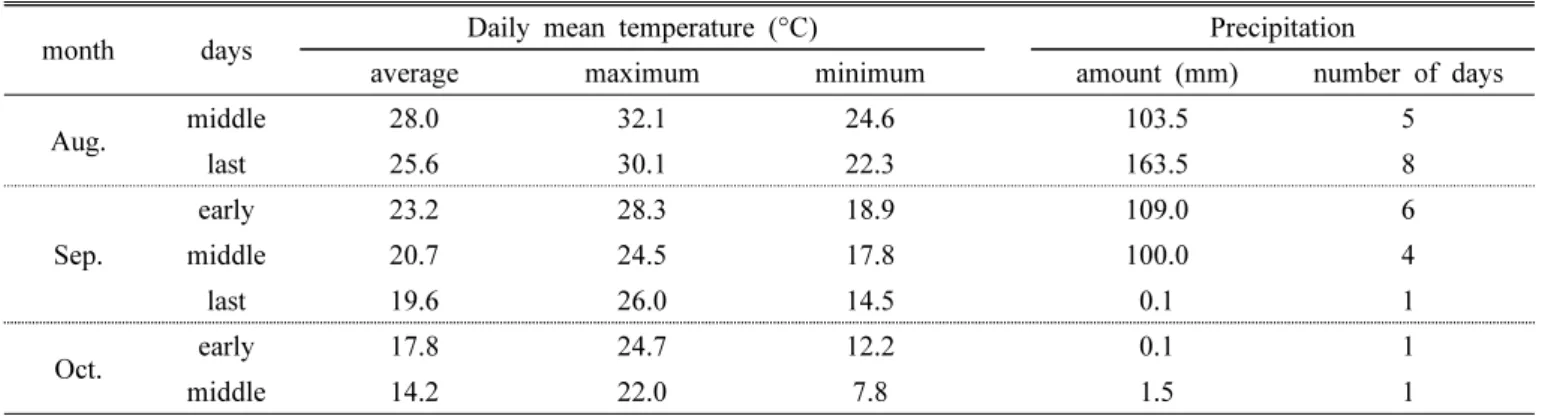
출수 후 종자의 등숙이 진행되는 과정에서 호품벼는 출수 후 20일부터, 신동진벼는 출수 후 30일부터 발아가 시작되 었다. 즉, 호품이 신동진에 비해 종자의 발아력이 생성되는 시기가 빨랐다. 이후 50% 이상의 종자가 발아력을 갖는 시 기는 호품벼가 약 35일, 신동진벼가 약 45일로 이 역시 호 품이 신동진에 비해 빠르게 나타났다(호품 57.0%, 신동진 56.8%). 출수 후 50일의 발아율은 호품벼와 신동진벼 모두 80% 이상이었으며(호품 82.6%, 신동진 92.3%), 출수 후 60 일에는 99% 이상의 발아율을 보였다(호품 99.5%, 신동진 99.4%)(Fig. 1).
Fig. 1. Seed vigor and germination percentage of panicles according to days after heading in Hopum and Shindongjin(vertical bars in graph are standard deviation).
Fig. 2. Order of pre-harvest sprouting of seeds in a panicle according to days after heading in Hopum and Shindongjin.
결론적으로 등숙일수에 따른 발아시기는 호품벼와 신동 진벼 간의 차이를 보였으나 출수 후 55일 이후에는 두 품종 에서 모두 높은 수발아율을 보였다. 이는 기존의 연구 결과 (Benech-Arnold and Sánchez, 2003; Gubler et al., 2005;
Mora et al., 2013)와도 일치하는 것으로, 벼의 등숙 후기에 강우나 과습한 상황이 지속될 경우 수발아 피해가 크게 발 생 될 수 있다.
한편, 한 이삭 내에서 종자의 위치에 따른 수발아의 순서
는 상위부 > 중위부 > 하위부 순으로 나타났으며 지경별로 는 2차 지경보다는 1차 지경의 것이 높게 나타났다(Fig. 2).
종자의 수발아 순서는 품종에 관계없이 상위 지경 일수록 빠르고 지경 내에서는 최선단의 종자가 빨랐다. 이러한 결 과는 벼의 출수기에 이삭 내 영화의 개화순서(Hoshikawa, 1989)와 유사한 경향을 보여 종자의 발아순서도 영화의 분 화발달의 순서와 일치하는 것으로 판단된다.
Fig. 4. Germination percentage of non pre-harvest sprouting seeds after hulling in Hopum and Shindongjin (vertical bar: standard deviation).
Fig. 3. Tetrazolium staining to observe morphological development of ungerminated seeds in Hopum and Shindongjin.
수발아 되지 않은 종자의 휴면성 검정
벼 출수 일수에 따른 수발아 검정에서 발아되지 않은 종 자를 tetrazolium으로 염색하여 배의 발달형태를 조사한 결 과, 출수 후 15일된 벼 종자도 배나 배유 조직의 형태적 미 숙 상태는 없는 것으로 관찰되었다(Fig. 3).
수발아는 종자 휴면과 밀접한 관련이 있는 것으로 보고되 었다(Farnsworth, 2000; Li et al., 2004; Yang et al., 2007).
출수 후 일수 별로 벼 이삭을 침종하여 수발아를 검정하는 동안 발아되지 않은 종자가 휴면에 의한 것인지를 검정하기 위하여 휴면의 원인으로 보고된 왕겨 (Koornneef et al., 2002; Lee et al., 2006)를 제거하고 발아율을 조사한 결과, 출수 후 15-20일 된 종자는 두 품종에서 모두 발아되지 않 았다(Fig. 4).
출수 후 30일 된 종자는 호품벼와 신동진벼에서 각각 8.8%와 50.5%가 발아되었으며, 출수 후50일에는 호품벼가 71.0%, 신동진벼가 81.5%가 발아되었다. 그러나 출수 후55 일 된 호품벼의 발아율은 24.6%, 신동진벼의 발아율은 43.2%였으며, 출수 후 60일에는 두 품종에서 모두 0%의 발 아율을 보였다. 이처럼 출수 50일 이후에 종자의 현미발아 율은 급격히 감소하는 추세를 보였는데, 이는 등숙후기에 미발아한 종자가 대부분 불완전립(출수 후 55일 불완전립 율: 호품 74.5%, 신동진 56.8%, 출수 후 60일 불완전립율:
두 품종 모두 100%)이었기 때문이다.
출수 후 30~45일에 신동진벼는 호품벼보다 수발아율이 낮았으나, 수발아 되지 않은 종자의 왕겨를 제거하여 검정 한 발아율을은 신동진벼가 호품벼보다 훨씬 높게 나타났다.
이는 신동진벼가 호품벼보다 등숙중기에 종자 휴면성이 높 기 때문으로 판단된다.
Gao et al.(2008)과 Gubler et al.(2005)은 종자휴면이 수
발아 저항성과 직접적인 관련이 있으며 이것은 종자발아를 억제하는 적응형질이라고 하였다. 마찬가지로 본 연구에서 도 벼 수발아성은 종자의 휴면 정도와 밀접한 관련이 있음 을 확인하였다. 이것으로 볼 때 벼의 수발아를 피하기 위해 서는 품종의 휴면성 도입이 바람직하고 볼 수 있만 종자의 강한 휴면은 종자 파종 시 불균일한 발아를 가져올 수 있으 므로 적당한 수준의 휴면성 도입이 요구된다.
적 요
본 연구는 벼의 출수 후 일수에 따라 수발아 양상을 조사 하고 수발아 피해가 발생할 수 있는 시기를 예측하여 수발 아 방제를 위한 기초자료를 얻고자 수행하였으며, 그 결과 를 요약하면 다음과 같다.
1. 벼 등숙기간 중 호품벼는 출수 후 20일, 신동진벼는
출수 후 30일부터 수발아 조건이 충족되면 발아가 시 작되었으며, 50% 이상의 종자가 발아력을 갖는 시기 는 호품벼가 약 35일, 신동진벼가 약 45일이었고, 출 수 후 50일에는 두 품종에서 모두 80% 이상의 종자가 발아하였다.
2. 한 이삭 내에서 종자의 위치에 따른 수발아의 순서는 상위부 > 중위부 > 하위부 순으로 나타났으며 지경별 로는 2차 지경보다는 1차 지경의 것이 높게 나타났다.
3. 수발아 검정 시 미발아된 종자는 배의 미숙, 종자휴면, 종자의 등숙불량 (불완전립)이 원인인 것으로 판단되 었다.
4. 출수 후 30~45일의 등숙중기에 신동진벼는 호품벼보 다 종자 휴면성이 높았다.
인용문헌