Isolation of Rice Putative STY Kinase Gene Induced by both Abiotic and Biotic Stress
Mi-Jeong Jeong,* Beom-Gi Kim, Jin-Ohk Lee, Sun-Young Kim, Soo-Chul Park and Sung-Joo Go
Molecular Physiology Division, National Institute of Agricultural Biotechnology, Rural Development Administration, Suwon 441-707, Korea
Received November 6 2003; Accepted March 11 2004
We isolated a novel protein kinase gene from Oryza sativa, and observed that expression of the gene is induced in respond not only to cold stress but also to drought and wounding stresses. Amino acid sequence deduced from the gene showed homology with that of a possible dual-specificity Ser/Thr/Tyr (STY) kinase. Time course expression profiling showed that mRNA expression of this gene reached maximal level at the early stage of stress by cold, drought, or wounding treatments. Our data suggest that the putative STY kinase plays a key role in upstream of abiotic and biotic signal transduction pathways and may also be involved in convergent signal pathways of abiotic and biotic stresses. In addition, we isolated several stress-related genes encoding proteins that interact with the putative STY kinase using the yeast two-hybrid screening system.
Key words: Abiotic and biotic stresses, signal transduction, dual-specificity protein kinase
Growth and productivity of plants are affected by exposure to biotic or abiotic factors such as drought, low temperature, and high salt. Wounding also presents a constant threat to plant survival because it not only physically destroys plant tissues, but also provides a pathway for pathogen invasion.
Plants responding to these abiotic and biotic stress signals have been extensively studied at physiological and biochemical levels, but the perception of the signals and intracelluar transmission mechanisms are largely unknown.1~4)
Protein phosphorylation plays important roles in many cellular signaling processes in eukaryotic system. In mammalian and yeast cells, mitogen-activated protein kinase (MAPK) modules, constituted by three functionally interlinked protein kinases, mediate various cellular responses to a variety of extracellular stimuli.5~8) There are increasing number of evidences indicating that protein kinase pathways are also involved in signal transduction of plants.9~13) After exposure of plant cells to fungal elicitors9) and ethylene12), and during establishment of freezing tolerance, changes in protein phosphorylation patterns were reported by several research groups.11) Furthermore, a number of plant genes encoding protein kinase have been reported.9,12)
Phosphorylation of protein on tyrosine (Tyr) and serine (Ser)/threonine (Thr) residues is achieved by protein Tyrrosine kinases and protein Serine/Threonine kinases, respectively. It is well known that phosphorylation of Ser/Thr and Tyr residues on target proteins represents an important
biochemical mechanism for regulating enzyme activity and many other cellular processes.13,14) Dual-specificity protein kinases are a group of protein kinases able to phosphorylate both Tyr and Ser/Thr residues. Dual-specificity kinases ATN1, APK1, and ADK1 from Arabidopsis15~17), PRK1 from Petunia18), PK12 from tomato19), and PmGK6 from soybean10) have been isolated. However, biological roles of these kinases remains unknown. Stress-induced STY kinase from peanut has been studied for their possible involvement in stress signaling.7) According to the Northern blot result, the STY kinase gene expression was markedly increased by cold and salt treatments, suggesting that the peanut STY kinase participates in signal transduction pathways induced by both cold and salt stresses.
Here, we report the identification of a new class of dual- specificity Serine/ Threonine/Tyrosine (STY) kinase of rice (Oryza sativa) involved in biotic and abiotic stresses.
Materials and Methods
Plant treatments. Tetrad stage rice plant (ecotype:
Donggin) was used in this study. Cold stress was conducted by exposing a portion of the plant to a temperature of 4oC under continuous light. Wounding treatment was performed by cutting the leaves of rice plant with scissors and placing them on a bench for 4 h. For drought treatment a portion of the rice plant was dehydrated on Whatman 3 MM paper under room conditions. For salt stress, plants were grown in soil containing 250 mM NaCl solution. Plant samples subjected to stress treatments for various periods were frozen in liquid nitrogen for further analyses.
*Corresponding author
Phone: +82-31-299-1724; Fax: +82-31-299-1722 E-mail: [email protected]
Construction of cold-induced subtractive library and isolation of the putative STY kinase gene. Total RNA was isolated with Trizol Reagent (Gibco-BRL) from non-stressed control and cold-stressed rice plant. Poly(A)+ RNA from the control plant was purified using biotinylated Oligo(dT)25. Cold-stressed rice plant cDNA was synthesized using the SMART PCR cDNA Synthesis kit (Clontech, USA).
Subsequent selective subtraction was carried out using the Streptavidin MagneSphere Magnetic separation products (Promega, USA) according to the manufacturer’s instructions.
Double-strand cDNA from the subtracted mRNA was synthesized with the SMART PCR cDNA Synthesis kit. After the second amplification, the PCR products were directly cloned in the pGEM vector (Promega, USA) and transformed into an Escherichia coli (XL1-Blue strain). Isolated DNAs from white clones were dot-blotted onto a Hybond-N’
membrane (Amersham Pharmacia Biotech, London). To search for low-temperature responsive genes, two copies of dot-blots were hybridized with 32P-labeled control and cold- stressed total cDNA.
RT-PCR and Northern blot analysis. Total RNA was extracted from leaves of the non-stressed control and various stress-treated plants. Reverse transcription was carried out in 80µl aliquots of reaction mixture containing 5x buffer (Promega, USA), 0.25 mM each of four deoxynucleotides (dNTPs), 2µg of synthetic poly-dT primer (5’-TTTTTTTTT TTTTTT-3’), 200 units of Moloney Murine Leukemia Virus (M-MLV) reverse transcriptase (Promega, USA), and 8µl DNase-treated RNA preheated at 72oC for 2 min. The reaction was performed at 37oC for 1 h. The first strand cDNA products (5µl) was then employed as a template DNA for the amplification of putative STY kinase cDNA. The RT-PCR products were separated on 0.8% agarose gel. Total RNA samples (20µl) were electophoresed on agarose gel, and transferred to membrane. RNA blot was hybridized by the methods described by Sambrook et al.20), using the putative STY kinase cDNA as a probe. Hybridization was carried out at 65oC for 16 h.
Yeast two-hybrid system. Strain AH109 of Saccharomyces cerevisiae was used in this study. For two-hybrid assays, competent yeast cells were cotransformed with two plasmids
(bait and prey). The bait, pGBKT7, carried a GAL4 DNA- binding domain and TRP prototrophic marker. The prey, pGADT7-Rec, carried a GAL4 activation domain and a Leu prototrophic marker. The putative STY kinase gene was fused to the GAL4 DNA-binding domain. The ds cDNA was amplified by Long-distance PCR (LD-PCR) using BD SMARTTM Technology (Clonethch, USA). LD-PCR products were purified through column chromatography with a BD CHROMA SPIN-400 column. A cDNA/AD fusion library was constructed by cotransforming the yeast with SmaI-linearized pGADT7-Rec and ds cDNA generated with the BD SMART library construction protocol. The ability to drive the HIS3 reporter gene was assessed by growing yeast transformants containing bait and prey plasmids on selective SD medium lacking Adenine, Histidine, Leucine, and Tryptophan. The activity of the MEL1 reporter gene was measured by in vivo agar plate assay using X-α-Galatosidase according to the manufacturer's protocol (Clontech, USA).
Results and Discussion
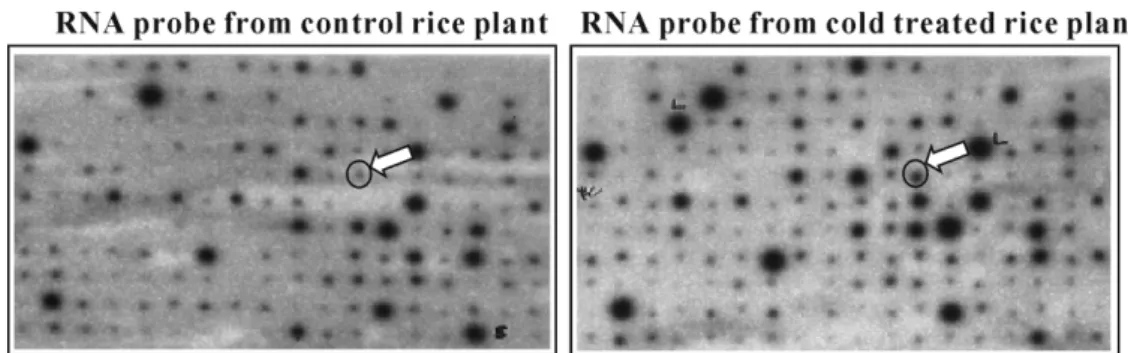
Isolation and sequence analysis of cold-induced putative STY kinase gene from rice. Subtractive screening with reverse Northern was performed to isolate cDNA clones induced by cold treatment. After second screening of the cDNA library by reverse Northern, over 20 clones were selected for sequencing. Finally, a cDNA clone (1.2 kb) showing high homology to putative protein kinase up- regulated in cold-treated rice plant was isolated (Fig. 1).
BLAST search revealed that a putative protein kinase encoded by the cDNA has 382 amino acids, with features characteristic of dual-specificity protein kinase. The putative protein was predicted to have an estimated molecular mass of 42 kD and a pI of 8.32, and contained nine conserved catalytic domains that are well conserved among dual specificity STY kinases.
The putative STY kinase shared 61% homology with a protein kinase from Populus nigra. Five other protein kinases, Arabidopsis PBS1 (52%), Vitis aestivalis protein kinase (54%), rice protein kinase (56%), Arabidopsis CDG1(56%), and Brassica PERK1(50%), shared identities with the putative STY kinase over the catalytic domain.
Fig. 1. Second screening of cold-induced rice genes isolated by subtractive screening with Reverse Northern. cDNA probes of the control and 5 min cold-treated early stage rice plants were used to isolate cold-induced rice genes.
Expression pattern of the putative STY kinase gene under various stress condition. RT-PCR was carried out to investigate the activation of the putative STY kinase gene by environmental stresses such as cold, salt, drought, wound, and darkness. Rice plants were subjected to 4oC cold treatment for 1 h, and 0.25 M NaCl solution for 3 h for salt stress. Rice plants were harvested 1 h after dehydration treatment for drought stress, 1 h after wounding treatment for biotic stress, or 4 days after darkness treatment. Results revealed that level
of mRNA of the putative STY kinase gene increased when rice plants were placed under cold, drought, or wounding stress conditions, whereas no significant changes were observed with salt or darkness treatments (Fig. 3).
Time course expression pattern of STY kinase gene under cold stress. Increase in putative STY kinase gene transcipt levels was observed with cold treatment (Figs. 4).
Therefore, cold-treated rice plants were harvested at different time points and their total RNA were isolated. RT-PCR and Northern blot analysis showed that putative STY kinase gene expression increased dramatically within 15 min of cold treatment. After 1 h, however, transcript level of the putative STY kinase gene slightly decreased, and then decreased to even lower level than that of control by 4 h.
Time course expression pattern of STY kinase gene under drought and wound stress. Increase in mRNA level was observed in rice plants subjected to dehydration, with maximum between 1 and 2 h, then decreased to that of the control by 4 h. In the case of wounding treatment, the putative STY kinase gene was rapidly and strongly expressed within 10 min and thereafter slightly decreased to level close to that of the control. After 4 h, no significant mRNA accumulation Fig. 2. Analysis of mRNA expression of putative STY
kinase gene in response to cold, high salt, drought, wound, and dark by RT-PCR reaction. Total RNA (20 µg) isolated from rice plants were subjected to various stresses. The puta- tive STY kinase cDNA was generated by specific anti-sense primer and amplified by PCR reaction with specific sense and antisense primer set. Cold: 4oC for 1 h; Salt: 0.25 M NaCl for 3 h; Drought: dehydrated for 1 h; Wounding: 1 h after initia- tion of wound treatment; Dark: 4 days after darkness treatment.
Fig. 3. Time course analysis of putative STY kinase gene expression in response to cold treatment. Total RNA (20 µg) isolated from rice plants were treated at 4oC for indicated times.
The putative osSTY cDNA was generated by specifically designed antisense primer, and amplified by PCR reaction with specific sence and antisense primer set.
Fig. 4. Time course analysis using RT-PCR reaction of puta- tive STY kinase gene expression subjected to drought (A) and wounding (B). Total RNA (20 µg) was isolated from rice plants treated with drought or wounding at various time points.
The putative protein kinase cDNA was generated by specific antisense primer, and amplified by PCR reaction with specific sense and antisense primer set.
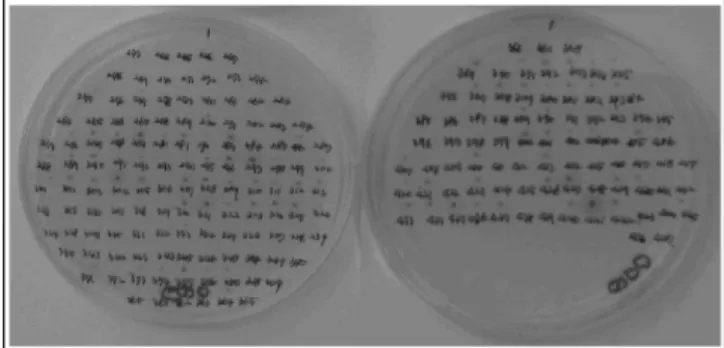
Fig. 5. Screening of Oryza sativa cDNA Library for two- hybrid interactions. Rice cDNA, pGADT7-Rec, and pGBKT7/
bait were cotransformed into yeast strain AH109. The presence of both plasmids was monitored by growth on synthetic com- plete media lacking Ade, His, Leu, and Trp (QDO). Activity of the MEL1 gene was measured by in vivo agar plate assay using X-α-Gal.
possible interaction between wounding and abiotic stress responses. In our study similar result was obtained in that this gene rapidly responded to both biotic and abiotic stresses, an indication that it is likely to play a key role in upstream of abiotic and biotic signal pathways, possibly cross-talking.
Identification of the genes encoding proteins that interact with the putative STY kinase. To screen for rice cDNAs encoding proteins that bind to the putative STY kinase, a yeast two-hybrid assay was performed. The putative STY kinase fused to the GAL4 DNA-binding domain reacted with expressed protein pool of rice cDNA libraly fused to the GAL4 activation domain gene. The interaction between proteins of putative STY kinase gene and cDNAs resulted in the expression of HIS3, which enabled the yeast cells to grow on the SD selective medium lacking Ade, His, Leu, and Trp.
As a negative control, the yeast cells cotransformed with the GAL4-fused putative STY kinase gene and Lam-fused cDNA library construction did not grow on the selective medium.
Using the yeast two-hybrid screening system, we isolated several genes encoding including PAL gene, -glucanase gene, and heat shock protein gene that specifically interact with the putative STY kinase by their coding proteins. Results of in vivo agar plate assay using X-Gal reconfirmed this result.
Further studies such as RNAi gene blocking experiment will be performed to reveal the function of this gene and downstream events of the signal pathway regulated by this gene.
Acknowledgments
This work was supported by the Biogreen21 Program grant provided by Rural Development Administration (RDA).
References
1. Cheong, Y-H., Chang, H-S., Gupta, R., Wang, X., Zhu, T., and Luan, S. (2002) Transcriptional profiling reveals novel interactions between wounding, pathogen, abiotic stress and hormonal responses in Arabidopsis. Plant Physiol. 129, 661-677.
2. Knight, H and Knight, M R. (2001) Abiotic stress signal- ing pathways; specificity and cross-talk. Trends in Plant Science Vol. 6, 262-267
3. Reymond, P., Weber, H., Damond, M., and Farmer, EE.
(2002) Differential gene expression in response to mechani- cal wounding and insect feeding in Arabidopsis. Plant Cell 12, 707-719.
4. Yamaguchi-Shinozaki, K. and Shinozaki, K. (1994) A novel
6. Ozaki I., Tani E., Ikemoto H., Kitagawa H., Fujikawa H.
(1999) Activation of stress-activated protein kinase/c-Jun NH2-terminal kinase and p38 kinase in calphostin C- induced apoptosis requires caspase-3-like proteases but is dispensable for cell death. J. Biol. Chem. 274, 5310-5317.
7. Robinson, MJ. and Cobb, MH. (1997) Mitogen-activated protein kinase pathways. Curr. Opin. Cell Biol. 9, 180-186.
8. Toda, T., Dhut, S-FG., Gotoh, Y., Nishida, E., Sugiura, R., and Kuno, T. (1996) The fission yeast pmk1+ gene encodes a novel mitogen-activated protein kinase homolog which regulates cell integrity and functions coordinately with the protein kinase C pathway. Mol. Cell Biol. 16, 6752-6764.
9. Droillard, M-J., Thibilliers, S., Cazale, A-C., Barbier-Bry- goo. H., and Lauriere, C. (2000) Protein kinases induced by osmotic stresses and elicitor molecules in tobacco cell uspensions : two crossroad MAP kinases and one osmoreg- ulation-specific protein kinase. FEBS Letters 474, 217-222.
10. Feng, XH., Zhao, Y., Bottino, PJ., and Kung, SD. (1993) Cloning and chracterization of a novel member of protein kinase familyfrom soybean. Biochem. Biophys. Acta. 1172, 200-204.
11. Monroy, AF., Sathan, F., and Dhindsa. RS. (1993) Cold- induced changes in freezing tolerance, protein phosphoyla- tion, and gene expression. Plant Physiol. 102, 1227-1235.
12. Ouaked, F., Rozhon, W., Lecourieux, D., and Hirt, H.
(2003) A MAPK pathway mediated ethylene signaling in plant. EMBO. 22, 1282-1288.
13. Rudrabhatla, P., and Rajasekharan, R. (2002) Developmen- tally regulated dual-specificity kinase from peanut that is induced by abiotic stresses. Plant Physiol. 130, 380-390.
14. Hunter, H. 1987. A thousand and one protein kinases. Cell 50, 823-829.
15. Ali, N., Halfter, U., and Chua, NH. (1994) Cloning and bio- chemical chracterization of a plant protein kinase that phos- phorylates serine, threonine and tyrosine. J. Biol. Chem.
269, 31626-31629.
16. Hirayama, T. and Oka, A. (1992) Novel protein kinase of Arabidopsis thaliana (APK1) that phosphorylates tyrosine, serine, and threonine. Plant Mol. Biol. 20, 653-662.
17. Tregear, JW., Jouannic, S., Schwebel-Dugue, N., Kreis, M.
(1996) An unsual protein kinase displaying chracteristics of both the srine/threonine kinase and tyrosine families is encoded by the Arabidopsis thaliana gene ATN1. Plant Sci.
117, 107-119.
18. Mu, JH., Lee, HS., and Kao, TH. (1994) Characterization of a pollen-expressed receptor-like kinase gene of Petunia inflata and the activity of its encoded kinase. Plant Cell 6, 709-721.
19. Sessa, G., Raz, V., Savaldi, S., and Fluhr, R. (1996) PK12, a plant dual-specificity protein kinase of the LAMMER family, is regulated by the hormone ethylene. Plant Cell 8, 2223-2234.
20. Sambrook, J. and Russell, DW. (2001) Molecular cloning: a laboratory manual, 3rd ed. Cold Spring harbor Laboratory Press, Cold Spring Harbor, H. Y.