- 149 -
Inhibition of Murine Allergic Response by Monoclonal Interleukin-4 Receptor Antibody
Chul Hee Lee, M.D.1, Ja Bock Yun, M.D.2, Chae-Seo Rhee, M.D.1, Seok-Won Park, M.D.1 and Jeong-Whun Kim, M.D.1
ABSTRACT
Background and Objectives:Several strategies were used to control the IgE production by interfering the functional activities of IL-4. However, most of them revealed limited effects to reduction of allergic response. This study was aimed to investigate the effects of interleukin-4 receptor (IL-4R) antibody on allergic response in animal model of allergic rhinitis.
Materials and Methods:::Male BALB/C mice were sensitized with intraperitoneal injection of ovalbumin (OVA). IL-4R anti-: body was injected intravenously before intranasal challenge of OVA with ultrasonic nebulizer. Allergic symptoms, a number of eosinophils in nasal mucosa and serum level of OVA-specific IgE were evaluated. Results:In IL-4R antibody treated mice, allergic symptom score were decreased (53.5%) than in control mice. The number of eosinophils in nasal mucosa were also reduced (47.4%). However, serum level of OVA-specific IgE were not obviously reduced (14.3%). Conclusion:These results suggested that IL-4R antibody has a potential effect for the treatment of allergic rhinitis in vivo.
KEY WORDS:Interleukin-4 receptor antibody·Allergic rhinitis·Treatment.
INTRODUCTION
Allergic rhinitis is an IgE-mediated inflammatory di- sease of nasal mucosa. It induces nasal symptoms such as sneezing, rhinorrhea, and itching sensation. It also shows significant eosinophilic infiltration into nasal mu- cosa. Interleukin-4 (IL-4) is essential in the development of allergic disease because of its influence over helper T-cell development and IgE production. Further, IL-4 causes upregulation of vascular cell adhesion molecule- 1 on endothelial cells, which promote eosinophilic adh- esion and transmigration from blood into the tissues.1) Therefore, inhibition of IL-4 would be particularly he- lpful in the treatment of allergic diseases.
Several strategies were used to control the IgE pro- duction by interfering the functional activities of IL- 4;One of them is the utilization of cytokines that inhibits the IgE production. IL-4-induced IgE produc- tion is effectively inhibited by interferon-gamma.2) The other strategy is the treatment with antibodies directed against IL-4 or IL-4 receptor.3-5)
The effect of IL-4 is mediated through cell membrane associated receptors. IL-4 receptor (IL-4R) is heterodi- mer composed with 825 aminoacids. IL-4R is expressed on many cell types including B and T cells, monocytes, fibroblasts, epithelial cells and endotheli-al cells.6)
It is well known that IL-4 plays a critical role in the pathogenesis of allergic disease. Thus, inhibition of the role of IL-4 by blocking is considered as possible can- didate for the treatment of allergic rhinitis. In patient with allergic disease, allergen challenge induces signifi- cant increase in IL-4 receptor in nasal biopsy specimen.7) Therefore, molecules that interfere with binding of IL- 4 to its respective receptor would be of great interest in consideration of treatment of allergic disease. However, there were few reports for use of antibodies against IL- 4R to control allergic rhinitis. We have developed a murine allergic rhinitis model characterized by the pre-
1Department of Otorhinolaryngology-Head and Neck Surgery, Seoul National University College of Medicine, Seoul, Korea
2Department of Otolaryngology, Sanggye Paik Hospital, Inje University, Seoul, Korea
Address correspondence and reprint requests to Chul Hee Lee, M.D., Department of Otorhinolaryngology-Head and Neck Sur- gery, Seoul National University College of Medicine, 28 Yongon- Dong, Chongno-Gu, Seoul 110-744, Korea
Tel:82-2-760-3646, Fax:82-2-745-2387 E-mail:[email protected]
Accepted for publication on December 5, 2000
sence of IgE antibodies to an inhaled antigen and eosi- nophilic infiltration into nasal mucosa. In this animal model, we have investigate the inhibitory role of IL-4R antibody in the development of allergen-specific IgE, allergic symptoms, and eosinophilic infiltration into airway mucosa.
MATERIALS AND METHODS Mice
Experiments were performed with 8 week old male BALB/C mice, healthy in appearance, weighing 50- 80 gm (Korea Beagle Farm, Seoul, Korea).
Reagents
The following monoclonal antibodies (MoAbs) were used in this study;a rat anti-mouse IL-4 receptor (CDw124)(Genzyme, Cambridge, MA), purified rat IgG2a immunoglobulin isotype standard (PharMingen, San Diego, CA), and a HRP-conjugated rat anti-mouse IgE (PharMingen, San Diego, CA). Ovalbumin (chicken egg albumin, grade V, Sigma Chemical Co, Phillipsberg, NY) and aluminium hydroxide (Al (OH)3, Sigma Chemical Co, Phillipsberg, NY) were used in the indu- ction of allergic rhinitis in mice as allergen and adjuvant, respectively.
Induction of allergic rhinitis in mice
Allergic rhinitis was induced in 15 of the 20 mice, and 5 were used as a normal control group for comparative observation.
For the allergic group (n=15), 10 μg ovalbumin and 1mg Al (OH)3 gel was dissolved in normal saline.
0.1 ml of this solution (0.05 ml of 20 μg/ml oval- bumin and 0.05 ml of 2 mg/ml Al (OH)3 gel) was injected intraperitoneally in mice on the 1st, 5th, and 21th days. Antigen was challenged by nebulizing 1%
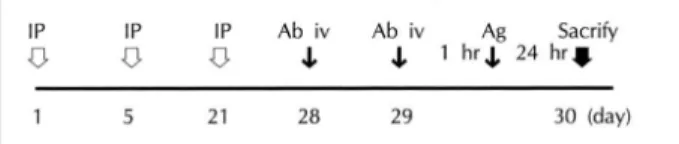
ovalbumin solution in a transparent acryl box for 5 mi- nutes. After then, we measured the symptoms (sneezing and nasal rubbing) for 30 minutes (Fig. 1). Ovalbu- min-sensitized animals were divided into three groups.
Each of group A mice was injected intravenously 100 μl of 1 mg/ml IL-4R MoAb into the tail vein, 24 hour and 1 hour before antigen challenge. In a group B, each mice was injected intravenously 100 μl of 1 mg/ml
isotype IgG2 antibody. In a group C, mice was received same amount of phosphate buffered saline (PBS). Each group was composed of 5 animals. The aerosol was ge- nerated with an ultrasonic nebulizer (PulmoAide, De- Villbiss, Somerset, Pa). The nebulized particle size was in the range of 1-5 μm. For normal control group (n
=5), same experiments was done using the equivalent amount of normal saline alone, instead of ovalbumin.
Eosinophilic infiltration in nasal mucosa
To evaluate the amount of eosinophilic infiltration in a mucosa, nasal mucosa was taken from the nasal septum at 24 hours after antigen challenge.
The nasal mucosa was fixed in 10% formalin fixa- tive for 24 hours, and embedded in paraffin. 4 μm of paraffin sections was placed on the slide and was stained with hematoxylin and eosin (H & E), or with Luna stain.
We examined the number of eosinophils under a light microscope at the magnification of 200×. The mean value of eosinophil counts in the 10 fields was taken to the eosinophil count in the specimen examined.
Determination of OVA-specific IgE
The serum from four groups of mice (immunization and IL-4R injection, immunization and PBS injection, immunization and isotype antibody injection, and nor- mal control) was collected on 24 hours after antigen challenge by inhalation. The OVA-specific IgE levels were determined by ELISA as follows. One hu-ndred microliters of OVA (5 μg in 0.2 mol/L carbonated buffer, pH 9.6) was dispensed in each well of a poly- styrene microtiter plate and incubated overnight at 4℃.
The OVA-coated plates were washed three times in 0.05% PBS-Tween buffer. The sera from the immunized and control groups were placed in the OVA-coated plate overnight at 4℃. The plates were washed five times with 0.05% PBS-Tween buffer and reacted with HRP-conjugated anti-mouse IgE antibody for 1 hour at
Fig. 1. Experimental Schedule. IP:intraperitoneal injection of 10 μg ovalbumin solution mixed with 1 mg A1 (OH)3, Ab iv:
intravenous injection of 0.1 mg anti- IL-4 receptor antibody, Ag:inhalation of 1% ovalbumin solution.
4℃. The plates were washed five times (0.05% PBS- Tween bufer) before the addition of citric acid-phos- phate buffer (pH 5.0) containing 0.15 mg/ml of o-phe- nylenediamine (Sigma Co., St. Louis, MO). The color was developed at room temperature and the reaction was stopped by 2.5 mol/L sulfuric acid. The color was measured at 492 nm using ELISA reader (Bio-Rad, Ri- chmond, CA).
Statistic analysis
The mean of measured values in a control and experi- mental groups was compared with Wilcoxon rank sum test at the significance of 0.05.
RESULTS Frequency of nasal symptoms
The number of sneezing and nasal rubbing were observed in the mice from the allergen-sensitized group into which ovalbumin was intranasally inhalated,. We presented sum of the frequency of sneezing and nasal rubbing as the symptom score. Allergic symptom score was reduced in IL-4R MoAb treated mice (9.3±3.8) compared to PBS-injected mice (20.8±4.8)(p<0.05) (Fig. 2). There was no difference in allergic symptom score between isotype IgG2a-treated mice (22.0±7.8) and PBS-injected mice (20.8±4.8)(p>0.05).
Eosinophilic infiltration into nasal mucosa
Hematoxylin and eosin and Luna stainings showed eosinophilic infiltration, mainly in the basement mem- brane. Eosinophil counts significantly decreased in IL- 4R MoAb-treated mice (9.0±8.2) than in PBS-injected mice (17.1±6.9)(p<0.05)(Fig. 3). There was, however, no difference in the number of eosinophils between isotype IgG2a-treated mice and PBS-injected mice (p>
0.05).
OVA-specific IgE responses
The groups of mice immunized with allergen (group A, B, and C) showed higher IgE responses than did normal control group. However, intravenous injection of IL-4R MoAb showed no evident reduction of OVA- specific IgE in group B mice (Fig. 4). Injection of iso-
Fig. 2. Effect of anti-IL-4R monoclonal antibody on the sym- ptom score induced by ovalbumin in sensitized mice. Anti-IL- 4R monoclonal antibodies were injected intravenously 24 hour and 1 hour before antigen challenge. Antigen challen- ge was performed by nebulization of 1% ovalbumin for 5 minutes. Nasal symptom score was counted for 30 min after antigen challenge. Symptom score is the sum of sneezing and nasal rubbing of mice. PBS:Phosphate buffered saline.
Fig. 3. Effect of anti-IL-4R monoclonal antibody on the infilt- ration of eosinophils induced by ovalbumin in sensitized mice.
Anti-IL-4R monoclonal antibody were injected intravenously 24 hour and 1 hour before antigen challenge. Antigen ch- allenge was performed by 1% ovalbumin nebulization for 5 minute. Nasal mucosal tissue was obtained and stained with Luna stain 24 hour after antigen challenge. PBS:Phosphate buffered saline.
Fig. 4. Effects of anti-IL-4R monoclonal antibody on the im- munoglobulin concentration in sensitized mice. Anti-IL-4R monoclonal antibody was injected intravenously 24 hour and 1 hour before antigen challenge. Antigen challenge was performed by 1% ovalbumin nebulization for 5 minute.
Serum was collected 24 hour after antigen challenge. Measu- rement of immunoglobulin concentration was done as des- cribed in Materials and Methods. PBS:Phosphate buffered saline.
type IgG2 antibody showed no changes in OVA-spe- cific IgE responses in group C mice, too.
DISCUSSION
Monoclonal anti-IL-4 antibody has been applied for inhibition of allergic response in a number of in vitro and in vivo studies.3)4) Although this approach is effec- tive in suppression of IgE production, large amount of MoAb is required for effective result. For this reason, in vivo treatment of anti-IL-4 antibody to allergic rhinitis is deemed impractical.
Another approach is the use of monoclonal anti-IL-4 receptor antibody. Induction of B cell activities by IL-4 can be inhibited by a receptor-specific monoclonal antibody in vitro.8) Recently, it has been reported that soluble IL-4 (sIL-4) receptor can inhibit IgE synthesis in mice in response to inhaled allergen.9)10) However, the other studies reported that sIL-4 receptor has a supportive role in maintaining IL-4 function and may prolong the half-life of IL-4.11)12) These reports suggest that soluble IL-4 receptor may inhibit or enhance aller- gic responses. Therefore, we used anti-IL-4R MoAb in this study.
In our study, allergic symptom score and eosinophilic infiltration were significantly decreased by anti-IL-4R MoAb, although allergen-specific IgE were not different between anti-IL-4R group and PBS injected groups (Fig. 3, 4). This result is somewhat different from previous report in which anti-IL-4R antibody blocked IgE responses by greater than 95%.5)
The reason of incomplete extent of inhibition in our study may be due to the action of other cytokines. IL- 13, a cytokine related to IL-4 that can bind to the alpha chain of the IL-4 receptor, and can induce allergic reaction in IL-4 independent fashion.13) Although IL-4 has been known to be a key role in allergy, IL-13 can mimic many of IL-4 effects with exception of Th2 di- fferentiation. Previous report demonstrated that selective neutralization of IL-13 ameliorate the asthma phenotype including airway hyperresponsiveness and eosinophil recruitment.14) Cytokine- mediated inhibition of the all- ergic response may depend on blocking both IL-4 and IL-13 activities.6)
Another possibility is that methods for induction of allergic model are different between previous reports and this study. Induction schedules or dosage of allergen may plays a role in a difference in serum IgE level in
animal models of allergy.
Even though allergen-specific IgE were not different between anti-IL-4R group and PBS-injected or IgG-in- jected groups, allergic symptom score and eosinophilic infiltration was significantly decreased by anti-IL-4R MoAb (Fig. 3, 4).
In human, some studies reported that specific IgE in serum may be sustained high titer despite of prolonged immunotherapy for allergy. Seasonal rises of IgE anti- bodies to ragweed did not return after treatment was stopped.15) Therefore, these reports suggested that re- duction of IgE in serum do not always be in company with the decrease of degree of allergic symptoms and eosinophil infiltrates.
IgE production is the result of an complex pathway such as T and B cell interaction, B cell activation, B cell proliferation and differentiation into plasma cells.
Possible mechanism of suppressive effect for allergic response by anti-IL-4R MoAb is the antibody binding to IL-4R, which interfere ligand-receptor interaction.
Another mechanism may be the inhibitory effect of en- dothelial cells on the regulation of adhesion molecules.
Endothelial cells respond to IL-4 with an upregulation of VCAM-1, while decreasing expression of ICAM-1 and ELAM-1. This leads to a transmembrane migration of eosinophils from circulating blood to tissues.1) Re- duction of the number of eosinophils in nasal mucosa in IL-4R MoAb treated group in this study may be due to the inhibition of this regulatory role of endothelial cells to adhesion molecules.
Thus, in this animal study using a monoclonal anti- body, despite of its incompleteness of reduction of allergen-specific IgE in murine serum, blocking of re- ceptor of IL-4 may have an important role for control of allergic responses.
CONCLUSION
We have demonstrated the inhibition of allergic res- ponse in murine allergic model by a monoclonal anti- body of IL-4 receptor. Despite of its incompleteness of reduction of allergen-specific IgE in murine serum, the amount of allergic symptoms and eosinophil infiltrate in nasal mucosa reduced markedly in this study. This showed that the administration of IL-4R antibody was one of effective tools in the treatment of allergic rhinitis.
AcknolwedgementThis research was supported by a grant from Seoul National University Hospital research fund, 04-95-028.
REFERENCES
1) Moser R, Fehr J, Bruijnzeel PLB. IL-4 controls the selective endothelial deriven transmigration of eosinophils from allergic in- dividuals. J Immunol 1992;149:1432-8.
2) Thyphronitis G, Tsokos GC, June CH, Levine AD, Finkelman FD.
IgE secretion by EB virus-infected purified human B-lymphocytes is stimulated by interleukin-4 and suppressed by interferon-γ.
Proc Natl Acad Sci USA 1989;86:5580-4.
3) Finkelman FD, Katona IM, Urban JF Jr, Holmes J, Ohara J, Tung AS, et al. Interleukin-4 is required to generated and sustain in vivo IgE response. J Immunol 1988;141:2335-41.
4) Finkelman FD, Katona IM, Urban JF Jr, Snapper CM, Ohara J, Paul WE. Suppression of in vivo polyclonal IgE responses by monoclonal antibody to the lymphokine BSF-1. Proc Natl Acad Sci USA 1986;83:9675-8.
5) Finkelman FD, Urban JF Jr, Beckmann MP, Schooley KA, Ho- lmes JM, Katona IM. Regulation of murine in vivo IgE and IgG responses by a monoclonal anti-IL-4 receptor antibody. Int Immunol 1991;3:599-607.
6) Ryan JJ. Interleukin-4 and its receptor: Essential mediators of the allergic response. J Allergy Clin Immunol 1997;99:1-5.
7) Wright E, Christodoulopoulos P, Small P, Frenkiel S, Hamid Q.
Th-2 type cytokine receptors in allergic rhinitis and in response to
topical steroids. Laryngoscope 1998;108:1528-33.
8) Maliszewski CR, Sato TA, Vanden Bos T, Beckmann MP, Gra- bstein KH. Induction of B cell activities by IL-4 is inhibited by a receptor-specific monoclonal antibody in vitro. Eur J Immunol 1990;20:1735-40.
9) Renz H, Enssle K, Lauffer L, Kurrle R, Gelfand EW. Inhibition of allergen-induced IgE and IgG1 production by soluble IL-4 rece- ptor. Int Arch Allergy Immunol 1995;106:46-54.
10) Renz H, Bradley K, Enssle K, Loader JE, Larsen GL, Gelfand EW.
Prevention of the development of immediate hypersensitivity and airway hyperresponsiveness following in vivo treatment with so- luble IL-4 receptor. Int Arch Allergy Immunol 1996;109:167-76.
11) Maliszewski CR, Sato TA, Davice B, Jacobs CA, Finkelman FD, Fanslow WC. In vivo biological effects of recombinant soluble IL-4 receptor. Proc Soc Exp Biol Med 1994;206:233-7.
12) Sato TA, Widmer NB, Finkelman FD, Madani H, Jacobs CA, Grabstein KH, et al. Recombinant soluble murine IL-4 receptor can inhibit or enhance IgE responses in vivo. J Immunol 1993;
150:2717-23.
13) Punnonen J, Aversa G, Cocks BG, McKenzie ANJ, Menon S, Zurawski G, et al. Interleukin 13 induces interleukin 4 indepe- ndent IgG4 and IgE synthesis and CD23 expression by human B cells. Proc Natl Acad Sci USA 1993;90:3730-4.
14) Grunig G, Warnock M, Wakil AE, Venkayya R, Brombacher F, Rennick DM, et al. Requirement for IL-13 independently of IL-4 in experimental asthma. Science 1998;282:2261-3.
15) Naclerio RM, Proud D, Moylan B, Balcer S, Freidhoff L, Kagey- Sobotka A, et al. A double-blind study of the discontinuation of ragweed immunotherapy. J Allergy Clin Immunol 1997;100:293-300.