Biomedicine & Pharmacotherapy 137 (2021) 111359
Available online 16 February 2021
0753-3322/© 2021 The Authors. Published by Elsevier Masson SAS. This is an open access article under the CC BY license
(http://creativecommons.org/licenses/by/4.0/).
Original article
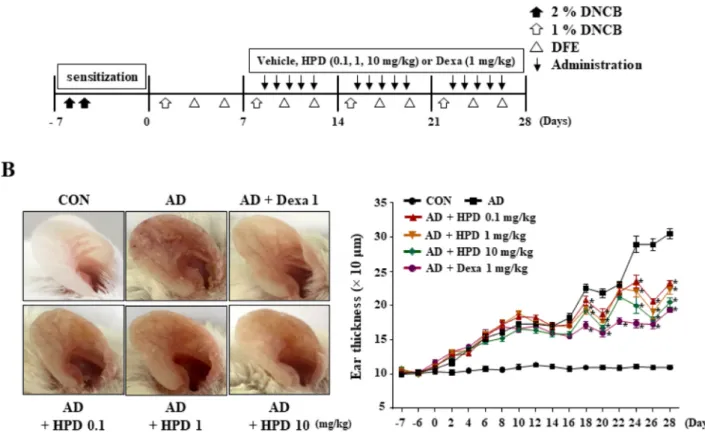
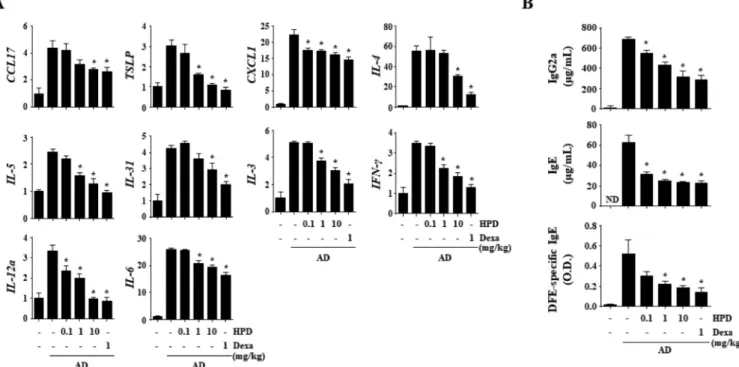
Hispidulin alleviates 2,4-dinitrochlorobenzene and house dust mite extract-induced atopic dermatitis-like skin inflammation
Jinjoo Kang a , Soyoung Lee b , Namkyung Kim a , Hima Dhakal a , Young-Ae Choi a , Taeg Kyu Kwon c , Dongwoo Khang d , * , Sang-Hyun Kim a , *
a
Cell & Matrix Research Institute, Department of Pharmacology, School of Medicine, Kyungpook National University, Daegu, Republic of Korea
b
Immunoregulatory Materials Research Center, Korea Research Institute of Bioscience and Biotechnology, Jeongeup, Republic of Korea
c
Department of Immunology, School of Medicine, Keimyung University, Daegu, Republic of Korea
d