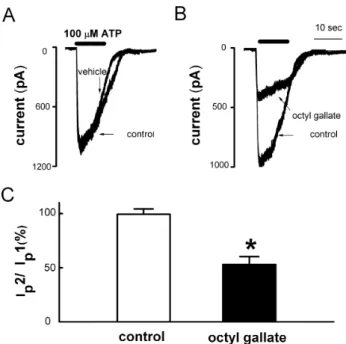
Octyl Gallate Inhibits ATP-induced Intracellular Calcium Increase in PC12 Cells by Inhibiting Multiple Pathways
8
0
0
전체 글
(2)
(3)
(4)
(5)
(6)
(7)
(8)
수치
![Fig. 1. Concentration-dependent inhibitory effects of octyl gallate on the ATP-induced [Ca 2+ ] i increase in PC12 cells](https://thumb-ap.123doks.com/thumbv2/123dokinfo/5471448.660206/3.892.89.815.134.711/concentration-dependent-inhibitory-effects-octyl-gallate-induced-increase.webp)
관련 문서
It also suggest that cell proliferation inhibits by a novel signal transduction for adenosine effect in human FaDu hypopharynx squamous cell carcinoma cells....
interaction with anti-apoptotic Bcl-2 family members, we investigated whether Mcl-1 can inhibit the Noxa-induced cell death.. As expected, Mcl-1 significantly inhibits
In other words, pomegranate extract has an effect of inhibiting the progression of alveolar bone loss due to periodontal inflammation by reducing the expression of COX-1 and
(d) displays the CA curves of the Ni2P2O7 electrocatalyst at an applied potential of 1.47 V (vs. • The steady-state CA curve acquired by the Ni 2 P 2 O 7 microsheets
In this study, it is shows that the stream of CO 2 is more effective in the reduction of unreacted TEGDMA and the increase of surface microhardness than that of N 2
Glutamate excitotoxicity induced by excessive activation of NMDA receptor causes various damage to cells, which leads to cell death.. In previous studies, increased ROS
Taking account of these results, in this work, it looks that chlorogenic acid depresses the CA secretory effect revealed by cholinergic secretagogues as well as Ang II
These results suggest that the bilobalide inhibits the cell proliferation and induces the apoptotic cell death in FaDu human pharyngeal squamous cell carcinoma via both
