대한소화기학회지 1999;34:632 - 643
8)
접수: 1998년 12월 26일, 승인: 1999년 5월 30일 연락처: 김학철, 570-711, 전북 익산시 신용동 344-2번지
원광의료원 소화기내과
Tel: (0653) 850-1069, Fax: (0653)
※ 이 논문은 원광대학교 1998년 교내연구비의 일부 지원 에 의해 수행되었음.
※ 이 논문의 요지는 1998년도 제37차 소화기학회 추계학 술대회에서 발표되었음.
서 론
B형 간염 바이러스(hepatitis B virus, HBV) DNA 는 약 3.2 kb의 이중 나선 DNA로서 4개의 open reading frame인 pre-core/core, pre-S/S, P 및 X 유전
만성 B 형 간염 바이러스 보유자에서 Cor e P r om oter 의 다양성
원광대학교 의과대학 내과학교실
김용성・서검석・송우건・김학철
N u c le o t i d e Va r i a b i l i t y o f Co r e P r o m o t e r R e g i o n i n Ty p e B Ch r o n i c Li v e r D i s e a s e s
Yo n g S u n g Ki m , M.D., Ge om S u k S e o , M.D., Wo o Gu n S o n g , M.D. a n d H a a k Ch e o u l Kim , M.D.
Department of Internal medicine, Wonkwang University College of Medicine, Iksan, Korea
Background/Aims: The core promoter (CP) is included in X-gene of hepatitis B virus (HBV), and
is composed of a upstream regulatory sequence (URS) and a basic core promotor (BCP) which is regulated by URS. Nucleotide (nt) mutation of CP might influence the expression of precore mRNA and pregenomic mRNA and the activity of liver disease. Thus, we investigated the relation between the nucleotide mutations in CP and their effects on HBeAg status, and severity of disease in chronic B viral liver disease. Methods: Samples were obtained from 7 chronic asymptomatic carriers, 9 patients with chronic hepatitis and 20 cirrhotic patients with HBV. Polymerase chain reaction and DNA analysis for CP were performed. Results: Cirrhotic group was older than asymptomatic carrie group and chronic hepatitis group. In BCP, point mutations were observed in 10 positions. Among them, nts 1753, 1762 and 1764 were frequently mutated, and the mutations at the nts 1762 and 1764 were accompanied (double mutation). In URS, point mutations were observed in 19 positions. Among these, nt 1653 (α-box) had significantly high mutation rate and the mutation at nt 1653 was closely associated with mutations at nts 1762/1764 in chronic liver disease and HBeAg (-) groupsConclusions: The α-box of URS is closely related with double-mutation in BCP and HBeAg status
in chronic B viral liver disease. (Kor J Gastroenterol 1999;34:632 - 643)Key Words: HBV, Core promoter, Nucleotide, Mutation,
-box김용성 외 4인. 만성 B형 간염 바이러스 보유자에서 Core Promoter의 다양성 633
자로 이루어져 있으며, 이들은 각각 HBeAg과 HBc- Ag, HBsAg, DNA polymerase 및 X 단백을 합성한 다.1 이러한 바이러스 유전자 산물들의 발현은 mRNA의 합성을 지시하는 4개의 촉진자(promoter) 에 의하여 조절되며, 4개의 촉진자들은 2개의 강화 인자(enhancer)에 의해 영향받고 있다.2
한편 HBV에 의한 감염은 무증상의 만성 보유자 부터 급성 간염, 만성 간염, 급성 전격성 간염 및 간 경변증 등의 다양한 형태로 나타난다. HBV 감염에 의한 간손상 기전은 첫째는 발병의 요인이 되는 HBV의 자체 요인, 즉 감염원인 바이러스의 양, 독 성, 유전자의 변이 형태가 이에 속하고, 둘째는 감염 된 숙주의 요인, 그리고 셋째는 환경적 요인으로 구 분할 수 있다. 이러한 각각의 요인을 명확하게 분석 하면 발병기전을 규명할 수 있을 것으로 생각되며, 최근에는 바이러스성 요인들이 대두되면서, 분자생 물학적 접근이 시도되고 있다.3-7
HBV의 X 유전자는 간종양의 발생과 밀접한 관 련이 있으며,8,9 또한 전격성 간염의 유발과 관련이 있다는 보고들이 있다.4,10 이러한 X 유전자는 465개 의 염기로 이루어져 있으며, 주기능은 교차활성화 (transactivating)를 하는 것으로 알려져 있다.9,11-13 X 유전자에는 core promoter(이하 CP로 약함), enhan- cer II(이하 EnII로 약함), direct repeat 1(이하 DR1으 로 약함), 그리고 direct repeat 2(이하 DR2로 약함) 등을 포함하고 있다.12 광의의 CP는 상부의 조절 서
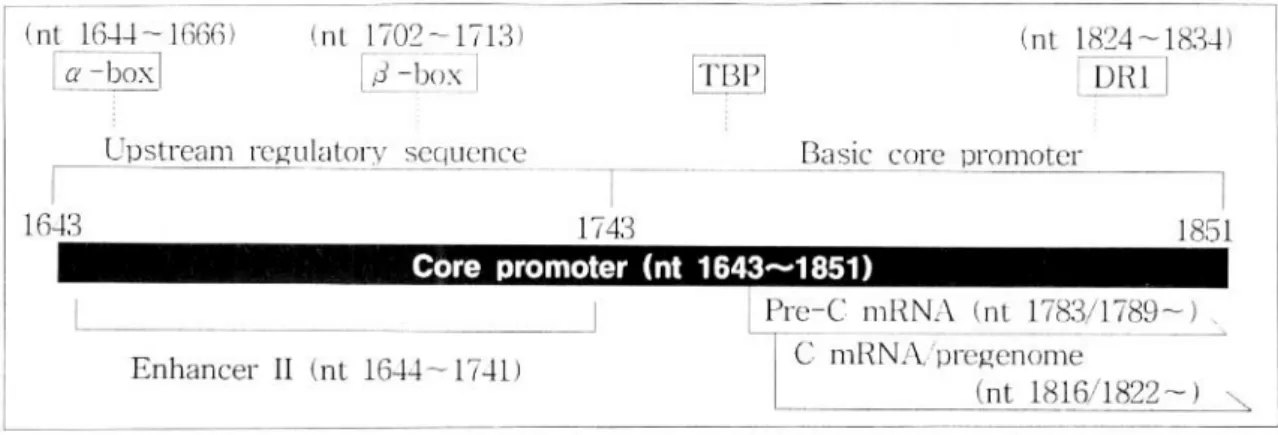
열(upstream regulatory sequence, 이하 URS로 약함) 과 기본 core 촉진자(Basic Core Promoter, 이하 BCP로 약함)로 구성되어 있으며, BCP는 URS에 의 하여 조절되고 있다고 한다(Fig. 1).14-17 CP는 X 유 전자의 후반부부터 C 유전자의 전반부 일부를 포함 하며, 이에는 α-box, β-box, EnII, DR1, hepatocyte nuclear factor(이하 HNF로 약함) III와 IV, TATA-box binding portion(이하 TBP로 약함) 등의 부위가 존재하여 여러 가지 기능을 한다.
HBV 증식은 precore mRNA와 pregenomic mRNA 의 전사에 의하며, 주로 CP의 영향을 받는다고 알려 져 있다.18-23 이에 따라 HBV에 의한 간질환의 심도 (severity)와 CP 변이와의 관련성이 추구되고 있 다.3,4,6,24 또한 CP 부위의 변이는 HBeAg과 core 항 원의 발현에 영향을 미쳐 간질환 활동도를 변화시킬 수 있을 것으로 여겨졌다. 그래서 이제까지의 연구 는 대부분 HBV에 의한 간질환에 대하여 전사와 복 제만의 관계로 추론하고자 하여 BCP 부위의 1762번 과 1764번의 염기 변이만이 연구되었으며,24-30 일부 에서만이 URS을 포함한 것이었다.3,4 이에 저자들은 만성 HBV 보유자에서 간질환의 정도, HBV 보유자 의 연령과 CP 전체 부위의 변이 유형과의 상관관계 를 관찰하고, 또한 변이형의 존재가 HBeAg의 발현 양태에 영향을 미치는가를 확인하고자 하였다.
Fig. 1. Schematic presentation of the structure of core promoter portion. The core promoter is included in X-gene and is composed of a upstream regulatory sequence (URS) and a basic core promotor (BCP). The URS contains α-box/β-box and regulate the BCP. TBP, TATA -box binding portion; DR1, direct repeat 1.
634 The Korean Journal of Gastroenterology : Vol. 34, No. 5, 1999
대상 및 방법
1. 대상 환자
확인된 만성 HBV 보유자로 만성 무증상 보유자 7예, 만성 간염 9예, 간경변증 환자 20예로 총 36명 을 대상으로 하였다. 이들 대상은 기왕력에서 면역 억제제, 면역조절제나 인터페론 등을 사용한 적이 없는 자로 한정하였다.
대상군의 분류는 만성 무증상 보유자군은 HBV에 의한 특이한 병변을 찾을 수 없었던 경우(간조직검 사상 지방간+6개월 이상의 HBV 보유자, 2예) 및 2 년 이상 HBsAg 양성으로 aminotransferase (AST &
ALT)가 정상으로 유지되었던 경우(5예)였고, 만성 간염군은 간조직검사에 의한 경우(6예)와 HBsAg이 5년 이상 양성이면서 관찰 기간 중 aminotransferase 가 2배 이상으로 1회에 6개월 이상 지속적으로 증가 되어 있었던 경우나, 관찰 기간 중 aminotransferase 의 상승이 9개월 이상인 경우(3예)로 정하였고, 간경 변증군은 간조직검사(4예)나 임상증상 등으로 확인 된 경우(16예)를 대상으로 하였다(Table 1).
2. 방 법 1) 혈청학적 검사
혈청학적 HBV의 표지자검사에서 HBsAg, anti- HBs, anti-HBc, IgM anti-HBc, HBeAg 및 anti-HBe 는 상품화된 kit (AUSRIA-II, Abott , Chicago, IL)를 이용하였고, HBV DNA는 탐침자를 이용한 방사면 역법을 이용하였다.
2) 혈청 H B V DN A 추출
대상군의 혈청 300 μl로부터 앞서 이용한 방법31 을 약간 변용하여 DNA를 추출하였다. 요약하면 동 량의 lysis buffer (2% SDS, 10 mM Tris-HCl, 150 mM NaCl, 10 mM EDTA pH 8.0, proteinase K 1 ㎎ /mL)와 혼합하여, 37℃에서 2시간 반응시킨 후, 동 량의 phenol로 잘 혼합하고 원심분리하여 상층액을 채취하였으며, 상층액이 투명할 때까지 반복 시행하 였다. 상층액에 동량의 phenol/chloroform/isoamyl- alcohol (25:24:1)을 혼합하여 원심분리한 후 이의 상 층액을 채취하여, glycogen (5 μg/mL)을 첨가하고, 2배량의 무수알코올을 첨가하여, -20℃에서 20분간 보관 후에 원심분리하여 추출하였다. 추출한 DNA 를 30 μl의 3차 증류수에 녹여, 이중 6 μl를 중합효 소연쇄반응(PCR)의 원형으로 이용하였다.
3) 시발체(pr im e r )의 선택 및 제조
HBV의 4가지의 혈청형(EMBO: adr, X04615;
ayw, Y07587; ayr, M57663; ady, D12980)에서 전체 X유전자 영역을 포함하는 부위의 핵산 배열을 관찰 하고자, X유전자의 외측에서 공통적으로 동일한 핵 산 배열을 갖는 곳을 선택하여, sense를 선택하였으 며, antisense는 저자들32이 국내의 C유전자 부위의 염기서열을 분석하여 변이가 가장 적은 부위를 선택 하였다. 일차 PCR에서 사용된 시발체는 5' -GGC- TCC-TCT-GCC-GAT-CCA-TAC-TGC-3' (sense, 1250
→1273), 5' -ATG-TTC-CGG- AGA-CTC-TAA-GGC- CTC (antisense, 2041→2018)로 증폭산물은 792 염
Table 1. Clinical Characteristics of Subjects
HBeAg Number M:F Mean age (years)±SD Asymptomatic carrier
Chronic hepatitis Cirrhosis of liver
+
-
+
-
+
-
4 3 6 3 14 6
4:0 2:1 5:1 2:1 9:5 2:4
38.1±14.1 (19~61)
38.3±9.9 (27~60) 48.1±12.6 (15~76)*
* denote significant difference between asymptomatic carrier/chronic hepatitis and cirrhosis of liver (p<0.05)
Kim, et al. Nucleotide Variability of Core Promoter Region in Type B Chronic Liver Diseases 635
기쌍(bp)이었고, nested PCR에 이용된 시발체는 5' - GAA-CTC-CTA-GCA-GCT-TGT-TTT-G-3' (sense, 1289→1310), 5' -CAG-AAG-CTC-CAA-ATT-CTT-TAT- A-3' (antisense, 1937→1916)로 최종 증폭산물은 649 bp였다.
4) H B V DN A의 증폭
HBV DNA 증폭은 대상 환자로부터 분리한 DNA 검체 6 μl와 각기의 시발체 20 pmole을 가하여 Taq polymerase 및 dNTPs 등이 함유되어 총 20 μl로 만 들게 준비된 TaKaRa Ex TaqTM (Takara Shuzo Co.
Shiga, Japan) kit를 이용여 Turbo thermal cyclerTM (바이오니아사, 충북 청원)에서 증폭하였다. 일차 PCR의 조건은 pre-denaturation을 94℃에서 5분, denaturation을 94℃에서 1분, annealing은 60℃에서 1분, extension은 72℃에서 1분씩 하여 총 30회 시행 하였으며, 마지막 extension은 72℃에서 5분간 시행 하였다. Nested PCR시는 일차 PCR 산물 2 μl와 이 차 시발체 한 쌍을 넣어서 pre-denaturation은 94℃에 서 5분, denaturation 94℃에서 1분, annealing은 59℃
에서 1분, extension은 72℃에서 1분씩 하여 총 30회 시행하였으며, 마지막 extension은 72℃에서 5분간 시행하였다. 이상의 모든 과정은 aerozol-tip 등을 사 용하면서, 오염 방지를 위한 수칙33을 준수하여 시행 하였다.
5) 염기 서열의 분석
상기에서 증폭된 2차 PCR 산물을 ethidium bro- mide으로 처리한 1.5% 한천겔에서 전기영동하여 650 bp 정도의 단일 증폭밴드임을 확인한 후, 이들 밴드를 절단하여 glass bead (Geneclean II Kit, Boeheringer Manheim, Germany)를 이용하여 순수한 유전자를 분리하였다. 분리된 정제물들의 일부를 채 취하여 ethidium bromide로 처리한 한천겔로 시발체 들이 없는 단일 밴드임을 확인한 후, 이를 2차 PCR 에 이용된 각기의 시발체 30 ng과 ddNTPs에 형광 물질을 결합시킨 것과 dNTPs, Taq polymerase 등을 함유한 Big dye terminator (Perkin-Elmer, CT, USA)를 제조사의 사용지시서대로 GeneAmp PCR system 2400 (Perkin-Elmer, CT, USA)을 이용하여
pre-denaturation은 95℃에서 30초, denaturation 95℃
에서 10초, annealing은 50℃에서 5초, extension은 60℃에서 4분씩하여 총 25회 시행하였다. 증폭된 PCR 산물은 에탄올침전법(0.3 M sodium acetate pH 5.2, 70% 에탄올)으로 정제하여 70% 에탄올로 세척 하였다. 이를 Template suppression reagent (Perkin- Elmer, CT, USA)에 녹여, 95℃에서 5분간 denatura- tion시킨 후 DNA 자동분석기(ABI Prism 310 ; Perkin-Elmer, CT, USA)에 넣어 분석하였다. 분석은 sense와 antisense를 이용한 comparative cross-matching 으로 sense의 방향에서부터 정리하여 이용하였으며, 핵산번호는 EcoR1 절단 부위를 기준으로 하여 정하 였다.
6) 분석된 유전자 염기서열의 다양성
염기서열의 분석은 모든 예에서 시발체로부터 30 bp 이상의 부위인 1330번에서 1880번까지의 550 bp 가 가능하였다. 분석된 모든 염기서열을 배열하여 주된 염기를 정하였고, 이와 차이가 있는 것을 다양 성(variability)으로 정하였다. 저자들이 제시하는 변 이 다발 위치는 각각의 동일한 염기서열에서 주된 염기와 다르게 보이는 곳에서, 이들의 변이가 평균 치±2표준편차 이상을 보이는 곳으로 정하였다. 또 한 전사의 시작 부위에서는 이를 기준 표준 염기 서 열(M12906)과도 비교하였다.
3. 자료 분석 및 통계 처리
모든 CP 부위(1643번→1851번)에서 측정된 자료 는 평균과 표준편차로 표시하였으며, 자료의 분석 및 통계 처리는 Student t-test, 분율 검정, ANOVA 및 Fisher' s exact test를 사용하여 p<0.05 이하를 유 의성으로 정하였고, 0.1<P<0.05를 높은 경향으로 처 리하였다.
결 과
1. 질환군에 따른 H B V 표지자와 임상상의 차이 무증상의 만성 HBV 보유자군, 만성 간염군 및 간 경변증 군에서 HBeAg와 anti-HBe의 발현에는 차이 가 없었으며, 연령은 만성 무증상 보유자군과 만성
636 대한소화기학회지 : 제 34 권 제 5 호 1999
간염군에서는 차이가 없었으나(38.1±14.1 vs 38.3
±9.9세), 간경변증군에서 유의하게 높았다(48.1±
12.6세, p<0.05, Table 1).
Table 2. Point Mutations in the Core Promoter of 36 HBV Carriers
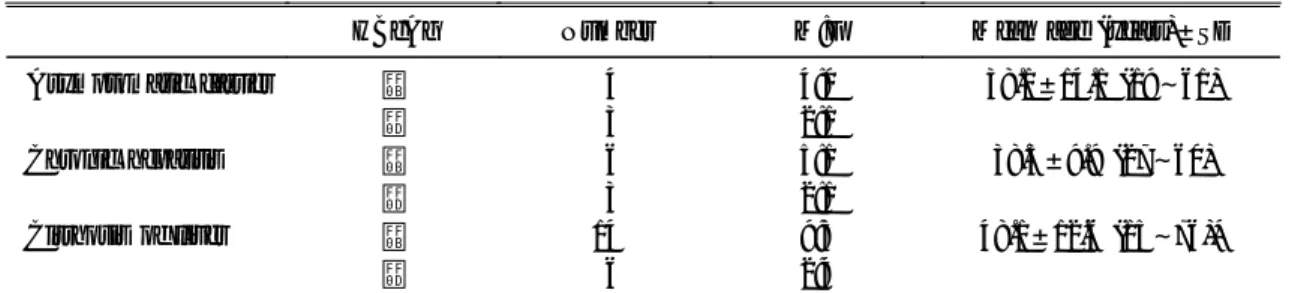
nt (wild type) 1647(G) 1653(C) 1676(A) 1727(A) 1753(T) 1762(A) 1764(G) ASC
ASC ASC ASC ASC ASC ASC CH CH CH CH CH CH CH CH CH LC LC LC LC LC LC LC LC LC LC LC LC LC LC LC LC LC LC LC LC
1*
2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36
- - - C - C - - - - - - - - - - - - - - - - - - - C - - - - - - - - - -
- - - - - - - - - - - - - - T - - - - - - - - - - - - - T T T T T T T T
- - - - - - - - - - - - - - - - - - - C - C - - - - C - - - - - - - - -
- - - - - - - - - - G
- - - - - G
- - - - - - - - - - G
- G
- - - - - -
- - - - - - G
- - - C - - - - - - - - - - - - A
- G A A - - - - - - - -
- - - - T T T - T T T T T T T T - - T T T T T T T T T T T T T T T T T T
- - - - A A A - A A A A A A A A - - A A A A A A A A A A A A A A A A A A ASC, asymptomatic carriers; CH, chronic hepatitis; LC, liver cirrhosis.
* Serial number of subject. The table contains nucleotide that have more than 3 mutations in one position an dashes indicate the same nucleotide as that in the top sequence.
김용성 외 4인. 만성 B형 간염 바이러스 보유자에서 Core Promoter의 다양성 637
2. C or e p r om ot er 부위의 염기서열의 변이 와 이에 따른 아미노산의 변이
CP 부위(1643-1851)에서의 염기 변이는 29곳에 서 발견되었으며, URS (1643-1742)와 BCP (1743-1851) 부위에서 각각 19곳과 10곳이었다(Fig.
1, Table 2). 장소에 따라 36예 중 1개의 변이부터 9 개의 변이까지 다양하게 나타났으며, 총 71개의 변 이가 관찰되었다. 따라서 CP 부위에서의 평균 변이 율은 0.95%였으며, 한 개의 변이 장소에서 평균 2.45±2.13개의 변이를 보였다. 표준의 염기서열 (M12906)과 비교한 경우 본 연구에서 주된 형으로 정한 서열 중 1762 (A→T)번과 1764 (G→A)번이 변 이로 산정되어 115개의 염기 변이를 보였으며, 평균 변이율은 1.54%였고, 변이 장소의 평균 변이는 3.97
±7.14개였다(Table 2). 아미노산의 변이가 없는 불 현성 염기 변이는 14개였으며, 한 곳(codon 127번) 에서는 1예에서 T가 A나 G로 변하므로 아미노산에 변이가 초래되었으나, 1예에서는 T가 C로 전환되어 아미노산의 변이가 없었다. 이러한 아미노산의 변이 는 15곳에서 1개부터 9개까지 다양하여 총 45개가 변화하여 평균 변이율은 1.87%였으며, 동일 변이 장소의 평균 변이율은 3.0±2.67개이었다. 이를 표 준의 염기서열(M12906)로 비교했을때는 평균 변이 율이 3.69%였으며, 동일 변이 장소의 평균 변이율 은 5.93±9.60개였다(Table 3).
3. C or e pr om ot er 부위의 염기서열의 변이의 특성
CP 부위의 평균 염기 변이율은 0.95%로, 변이의 발현율이 1653, 1762, 및 1764번에서 높았고, 1753
번에서는 높은 편이었다. 이들을 X 유전자의 codon 으로 관찰시 변이의 발현율은 94, 130 및 131번에서 높았으며, 127번에서는 높은 편이었다. 기존의 표준 염기서열을 기준으로 비교시, 변이에서 이들 상호간 의 관계는 1762번과 1764번과의 동반 변이가 유의 하게 일어났으며, 1653 (C→T)번의 변이는 1762 (A
→T)번과 1764 (G→A)번의 동반 변이시 높은 경향 (0.05<P<0.1)이었다. 따라서 codon 130과 131번 아 미노산의 동반 치환은 codon 94번에서 아미노산 치 환을 일으키는 경향이었다(0.05<P<0.1,Fig. 2). 이를 기능적인 부위에 따라 관찰하면, DR1 및 TBP에서 는 변이는 없었으며, β-box 및 CP의 말단부 등에서 는 변이율이 낮았으나, 특이적으로 많이 발생하는 부위는 α-box와 BCP 부위의 initiator의 직전방 부 위
Table 3. Amino Acid Changes in Core Promoter Portion
Codon No. X92 X94 X101 X102 X105 X106 X116 X119 X121 X127 X130 X131 X144 X146 PC11 Wild
Mutation V L3*
H Y10
S L1 P1
A S1
T M1
T S1
L V2
D N1
E D1
I N3 T1 S1
K†
M27 I2
V†
I29 S V2
A S1
A T2
X, X-gene codon number (X91~154); PC, Pre-C codon number (PC1~11).
* number of mutated amino acid.
†compared with standard DNA sequence.
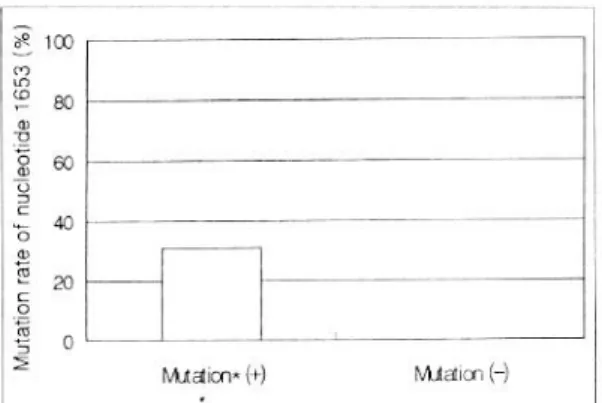
Fig. 2. Mutation rate of nucleotide 1653 according to nucleotide 1762/1764 double-mutation. The mutation of nucleotide 1653, the region of α-box, closely associated with double-mutation of nucleotide 1762/1764. * mutation, 1762/1764 double-mutation.
638 The Korean Journal of Gastroenterology : Vol. 34, No. 5, 1999
였다.
4. 질환군, H B e Ag 의 발현 양태 및 염기 변이 와의 관계
1) 질환군과 H B eA g 의 발현 양태와의 관계 질환군에 따른 HBeAg의 발현 양태는 각 군 간에 통계학적인 차이는 없었다(Fig. 3).
2) 질환군과 염기 변이와의 관계
표준 염기서열(M12906)과 비교한 경우, CP 부위
의 평균 염기 변이율은 무증상의 HBV 보유자에서 1.37%, 만성 간염군에서 1.35%, 간경변군에서 1.68%
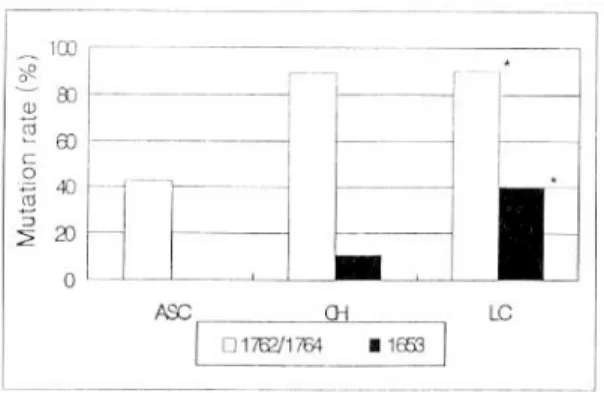
로 심한 간질환일수록 높았지만, 각 군 간에 유의한 차이는 없었다. 질환군에 따른 1762번과 1764번의 동반 변이율은 무증상 HBV 보유자군에 비하여 만 성 간염군에서 많았지만 비교 대상이 적어 경향만 을 보였으나, 무증상 HBV 보유자군에 비하여 간경 변군에서는 유의하게 많았다. 또한 간경변증군에서 1653번 변이율이 높은 경향이었으며 1762번과 1764 번의 동반 변이시 1653번의 변이율이 높았다(Fig.
4).
3) H B eA g 의 발현 양태와 염기 변이와의 관계 HBeAg 음성군에서 1762번과 1764번의 동반 변 이가 많았으나 통계학적인 의의는 없었고, 1653번 의 변이는 HBeAg 음성군이 92% (11/12예)로 HBeAg 양성군의 75% (18/24예)에 비해 의의 있게 많았다(p<0.05, Fig. 5).
4) 간경변증군에서 H B e Ag 의 발현 양태와 염기 변이와의 관계
간경변증군만을 대상으로 했을 때 HBeAg 음성군 에서는 1762번과 1764번의 동반 변이가 많았으나 통계학적인 의의는 없었고, 1653번의 변이는 HBeAg 음성군이 HBeAg 양성군에 비해 의의 있게 많았다(p<0.05, Fig. 6).
Fig. 3. HBeAg status according to subject subgroup.
There was no statistically significant difference between subgroups in HBeAg status. ASC, asymptomatic carriers;
CH, chronic hepatitis; LC, liver cirrhosis.
Fig. 4. Mutation rate of nucleotide 1762/1764, 1653 ac- cording to subject subgroup. There was high mutation rate of nucleotide 1762/1764 in CH and LC. Nucleotide 1653 had significantly high mutation rate in cirrhotic patients with nucleotide 1762/1764 double-mutation. ASC, asymp- tomatic carriers; CH, chronic hepatitis; LC, liver cirrhosis.
* p<0.05 compared to ASC group.
Fig. 5. Mutation rate of nucleotide 1762/1764, 1653 ac- cording to HBeAg status. Nucleotide 1762/1764 double- mutation was no difference according to HBeAg status, but nucleotide 1653 mutation was significantly high in HBeAg (-) subjects.
* p<0.05 compare to HBeAg (+) subjects.
Kim, et al. Nucleotide Variability of Core Promoter Region in Type B Chronic Liver Diseases 639
고 찰
HBV 감염에 의한 간손상의 기전 중 HBV의 자체 요인은 간질환에 따른 변이형의 발현,3-5,7 HBeAg의 발현에 영향을 미치는 pre-C 유전자의 변이, HBV의 한 간종양의 발생,8,9 변이형에 의한 전격성 간염의 유발 등4,10이 그 증거로 제시되고 있다. 이중 CP 부 위의 변이는 HBeAg과 core 단백의 발현의 전사 수 준에서 영향을 미쳐 간질환 활동도를 변화시킬 것 23,29,34이라는 추정하에, CP 부위의 변이와 HBeAg의 음양성, 간질환의 활성도, 간질환의 심화 정도 등 3,28,29,35과의 관계가 보고되고 있다.
이러한 연구의 대부분은 BCP 부위의 1762번과 1764번의 염기서열의 변화만을 관찰하였으며 일부 에서만이 BCP의 전반과 URS을 포함한 것이었다.3,10 하지만 BCP 부위는 상방에 존재하는 URS에 의해 영향을 받으므로, 본 연구에서는 CP (208 nucleo- tide)의 전 부위에서 변이를 관찰하고자 하였다. 저 자 등이 관찰한 CP의 평균 염기 변이율은 0.95%였 으며 변이된 장소에서의 평균 변이는 2.45±2.13개 로, 저자들32이 조사한 한국인에서 C유전자 부위의 변이율와 유사하였다. 본 연구에서 조사된 염기서열 의 특성을 보면, 염기 변이가 α-box와 복제 및 복사 의 initiator에 관련된 부위의 전방에서 주로 일어나, α-box와 BCP의 전반부에 매우 다양하다는 보고들
3,30과도 일치하였다. 또한 DR1과 TBP의 부위에서 는 변이 없이 잘 보존되고 있는 부위로 다른 보고30 와도 일치하였는데, DR 부위와 교차 활성화 부위인 TBP 부위에서 변이가 적은 것은 이 부위가 HBV의 특징을 보이는 부위일 가능성을 시사하는 것으로 생각된다.
CP 영역에서의 변이 다발 위치는 1653, 1762, 1764번이었고 1753번에서도 변이가 많은 경향이었 으며, 표준 염기형을 기준으로 관찰한 결과도 일치 하였다. BCP부위에서 1762번(A→T)과 1764번(G→
A)의 변이가 많음은 국내외의 여러 보고들과 일치 하였는데, 만성 간질환에서 주로 일어나는 것도 일 치하였다.3,25,26,35 이러한 염기 변이율이 많은 부위 (hotspot; 1653, 1762, 1764)에서 상호간의 관계를 관 찰한 결과 1762번(A→T)과 1764번(G→A)의 변이는 매우 밀접한 동반 변이로 존재하였고, 이것은 여러 저자들3,29,30,35에 의하여도 확인되었다. 이러한 동반 변이는 1명의 만성 간염 환자의 혈청에서 HBeAg가 anti-HBe로 전환되는 시기에 1764번의 G→A로 변이 가 일어난 후 1762번이 A→T로 전환됨을 관찰하여, 1764번에서 먼저 전환되고 뒤이어 1762번이 전환된 다고 하였으며,36 1997년 실험적으로 입증된 바24 있 다. 이러한 결과는 국내에서도 이를 간접적으로 입 증하는 보고들25,26이 있으며, 저자들의 연구에서도 유사하여 이와 같은 일련의 변화는 확실한 것 같다.
1762번과 1764번의 동반 변이와 1653번 변이와의 관련성은 1762/1764번의 동반 변이시에는 29명 중 9 명에서 1653번에서의 변이가 일어났지만, 1762/1764 번의 동반 변이가 없었던 유형에서는 관찰되지 않 아, 만성 간질환시에 1762번과 1764번의 변이가 호 발하며 1653번에서 C→T로의 변이가 다발한다는 다 른 보고들3,10과도 일치하는 소견이었다. 본 연구에서 1762/1764번의 동반 변이시에 유의하게 1653번의 변이가 일어남이 관찰되어 상호 관련성이 있을 것으 로 추정되었다.
HBeAg의 발현은 pre-C의 1896번이나 1899번의 염기 변이에 의해 결정되는데, 1762/1764번의 동반 변이는 precore의 전사를 감소시키고 genomic mRNA 의 전사를 증가시킴으로써, HBeAg의 발현에 영향을 주어서 HBeAg가 억제된다는 보고가 있다. 이러한 Fig. 6. Mutation rate of nucleotide 1653 according to
HBeAg status in liver cirrhosis group. Nucleotide 1653 mutation was significantly higher in HBeAg (-) cirrhotic patients (p<0.05).
640 대한소화기학회지 : 제 34 권 제 5 호 1999
이유로 1762/1764 동반 변이는 만성 HBV 보유자에 서 HBeAg 양성인 경우보다 anti-HBe 양성인 경우 에 보다 흔하고, 또한 무증상 HBV 감염자에서는 HBeAg 음성인 경우에 더욱 흔하다고3,29,30,35 하였다.
본 연구에서도 1762/1764번의 동반 변이는 HBeAg 가 음성인 경우에 많았으나 통계학적 의의는 찾지 못하였다. 하지만 1653번의 변이는 HBeAg 음성인 간경변증인 경우에 유의하게 많았다. BCP는 EnII에 의해 영향을 받으며,3,14-17 β box는 BCP에 직접적인 영향이 적으나 α-box의 기능을 강화시켜주므로 BCP에 대한 영향은 주로 α-box에 의한다는 보고로 미루어보아14 이들간에 상호 관련성이 존재할 것이 라 생각할 수 있는데, HBeAg 음성인 간경변증 환자 에서 α- box의 1653번의 변이가 유의하게 동반되어 HBeAg의 발현에 중요할 것이라는 본 연구 결과와 일치하는 바로 생각된다.
본 연구에서 무증상의 만성 HBV 보유자와 만성 간염 환자군의 사이에는 연령에는 차이가 없으나 1762/1764번의 동반 변이에 차이가 있음은 다른 보 고들26,28과 일치하는 소견으로서, 이는 1762/1764번 의 동반 변이가 만성 간질환과 밀접한 관계가 있을 것으로 생각된다. 간경변증군과 만성 간염군간에는 1762/1764번의 동반 변이에는 차이가 없고 1653번 변이는 차이가 있어, 최근 국내에서 간경변증 환자 군에서 1653번에 변이가 많이 발견된다는 보고37는 본 연구와 일치하는 소견으로 생각된다. 간경변증군 과 만성 간염군 간에는 1653번의 염기 변이와 연령 이 차이가 있는 것은 HBV에 의한 국내의 간질환이 대부분 모자 수직감염에 의한 것임을 고려할 때, 1653번의 변이가 질병의 자연 경과에 의한 HBV의 변화인지, 혹은 바이러스의 자체에 발병기전인지는 향후 연구 조사되어야 할 것으로 생각된다.
유전자형으로 분류 관찰하면서 BCP 부위에서 1751-1755번 사이가 변이 다발 부위(mutation clus- tering region)라고 보고된 바 있으며,30 국내의 보고 에서도 이를 언급하지는 않았지만 변이가 많음이 관 찰되었는데,26 본 연구에서도 1753번에서 변이가 많 은 경향을 보였다. 이 부위에서의 변이가 1762번과 1764번의 동반 변이시는 간질환이 유의하게 심하다 고 하였는데,30 본 연구에서도 간경변증군에서 많은
빈도를 보였다. 그러나 1753번의 변이와 1762/1764 번의 동반 변이와의 유관성은 조사된 예가 적어서 논하기는 어려웠다.
본 연구에서 CP 부위의 염기서열을 분석하여 이 를 연령, HBeAg과 anti-HBe의 발현 양태 및 간질환 군별로 분석하였으며, 이들간의 상호관계를 조사하 였다. 본 연구에서는 대상 환자 수가 적지는 않았으 나, 질환별이나 기타의 군간으로 분석시 일부에서는 대상이 적어 통계학적인 문제가 대두되는 경우도 있 어 더 많은 예들에서 확증되어야 할 것으로 생각되 었다. 분석 결과, 연령에서는 무증상의 HBV 보유자 와 만성 간염군간에는 차이가 없었으나, 1762/ 1764 번의 동반 변이는 주로 만성 간질환군에서 호발하 며, 1762/1764번의 동반 변이시 1653번의 변이가 자 주 발현하였다. 이러한 변이는 HBeAg 음성인 간경 변증군에서 많이 발견되었으나, 향후 많은 증례와 HBV 유전자형에 따라 검색되어야 할 것으로 생각 된다.
요 약
목적: B형 간염 바이러스의 증식은 core promo- tor (CP)의 영향을 받는 precore mRNA와 pregeno- mic mRNA의 전사에 의한다고 알려져 있으며, CP 의 변이는 HBeAg과 core 항원 발현에 영향을 미쳐, 간질환의 활성도를 변화시킬 것으로 추정하고 있다.
CP는 upstream regulatory sequence (URS)와 basic core promotor (BCP)로 구성되며, BCP는 URS의 와 -box 등에 의하여 영향을 받는다고 한다. 저자들 은 만성 B형 간염 바이러스 보유자에서 CP 부위의 염기서열의 변이와 이들간의 상호 관련성, 간질환의 중증도 및 HBeAg의 발현 양태에 미치는 영향 등을 관찰하고자 하였다. 대상 및 방법: 만성 HBV 보유 자로서 무증상 보유자 7명(HBeAg 양성 4명, 음성 3 명), 만성 간염 환자 9명(HBeg 양성 7명, 음성 2명) 및 간경변증 환자 20명(HBeAg 양성 14명, 음성 6명) 을 대상으로 하였다. 이들의 HBV DNA를 염기 분 석하여, CP 부위를 표준의 염기서열과 비교하였다.
결과: 연령은 무증상의 HBV 보유자군과 만성 간염 군보다 간경변증군에서 높았다. BCP 부위에서의 변
김용성 외 4인. 만성 B형 간염 바이러스 보유자에서 Core Promoter의 다양성 641
이는 10곳에서 관찰되었는데, 1753번은 변이의 발 현 빈도가 높은 경향만 보였으나, 1762번(A→T)과 1764번(G→A) 변이의 빈도는 유의하게 높았고 동반 변이를 보였다. URS 부위에서는 19곳에서 하나 이 상의 변이가 관찰되었는데, -box 부위인 1653번에 변이가 많이 관찰되었고 1762/1764번 동반 변이시 유의하게 관찰되었다. HBeAg의 발현 양태에 따른 1762/1764번 동반 변이 및 간질환의 중증도와는 연 관성이 없었지만, HBeAg 음성인 환자들에서는 1653 번 변이가 유의하게 높았다. 결론: 본 연구의 대상에 서 HBeAg 유무에 따른 간질환의 중증도의 차이는 없었으며, 연령은 간경변증군에서 높았다. 만성 간 질환군은 무증상군에 비하여 1762/1764번 동반 변이 가 흔히 관찰되었고, 이 동반 변이는 -box 부위인 1653번 변이와 밀접한 관련성이 있는 것으로 추정되 며, 1653번 변이가 1762/1764번 동반 변이보다 HBeAg 발현에 더 중요한 역할을 할 것으로 생각된다.
참 고 문 헌
1. Ganem D, Varmus HE. The molecular biology o the hepatitis B viruses. Annu Rev Biochem 1987;56 651-693.
2. Schaller H, Fisher M. Transcriptional control of hepadnavirus gene expression. Curr Top Microbiol Immunol 1991;168:21-39.
3. Nishizono A, Hiraga M, Kohno K, et al. Mutation in the core promoter/enhancer Ⅱ regions of naturally occurring hepatitis B virus variants and analysis o the effects on transcription activities. Intervirology 1995;38:290-294.
4. Sato S, Suzuki K, Akahane Y, et al. Hepatitis B virus strains with mutations in the core promoter in patients with fulminant hepatitis. Ann Intern Med 1995;122:241-248.
5. Günther S, Piwon N, Iwanska A, Schilling R, Mei sel H, Will H. Type, prevalence, and significance o core promoter/enhancer II mutations in hepatitis B viruses from immunosuppressed patients with severe liver disease. J Virol 1996;70:8318-8331.
6. Fukuda R, Ishimura N, Kushiyama Y, et al. Hepa
titis B virus with X gene mutation is associated with the majority of serologically "silent" non-B, non-C chronic hepatitis. Microbiol Immunol 1996;40:481- 488.
7. Bock ST, Tillman HL, Maschek HJ, Manns MP Trautwein C. A preS Mutation isolated from a patient with chronic hepatitis B infection leads to virus retention and misassembly. Gastroenterology 1997;113:1976-1982.
8. Tacheuchi M, Fujimoto J, Niwamoto H, Yamamoto Y, Okamoto E. Frequent detection of hepatitis B virus X-gene DNA in hepatocellular carcinoma and adjacent liver tissue in hepatitis B surface antigen negative patients. Dig Dis Sci 1997;42:2264-2269.
9. Moriarty AM, Alexander H, Lerner RA, Thornton GB. Antibodies to peptides detect new hepatitis B antigen: serological correlation with hepatocellular carcinoma. Science 1985;227:429-433.
10. Kaneko M, Uchida T, Moriyama M, et al. Probable implication of mutations of the X open reading frame in the onset of fulminant hepatitis B. J Med Virol 1995;47:204-208.
11. Suzuchi K, Uchida T, Shikata T, et al. Expression o pre-S1, pre-S2, S and X peptides in relation to vira replication in livers with chronic hepatitis B. Live 1990;10:355-364.
12. Uchida T, Saitoh T, Shinzawa H. Mutations of the X region of hepatitis B virus and their clinical implica tions. Pathol Int 1997;47:183-193.
13. Arii M, Tkada S, Koike K. Identification of three essential regions of hepatitis B virus X protein fo trans-activation function. Oncogene 1992;7:397-403.
14. Yuh CH, Ting LP. The genome of hepatitis B viru contains a second enhancer: cooperation of two ele ments within this enhancer is required for its function. J Virol 1990;64:4281-4287.
15. Yuh CH, Ting LP. C/EBP-like proteins binding to the functional box- and box- of the second enhan cer of hepatitis B virus. Mol Cell Biol 1991;11 5044-5052.
16. Yuh CH, Ting LP. Transcriptional regulation of pre core and pregenomic RNAs of hepatitis B virus.
642 The Korean Journal of Gastroenterology : Vol. 34, No. 5, 1999
Virol 1992;66:4073-4084.
17. Fourel G, Ringeisen F, Flajolet M, Tiollais P Buendia MA. Functional analysis of ground squirre hepatitis virus enhancer II. J Virol 1998;72:1616- 1622.
18. Wasylyk B, Chambon P. A T to A base substitution and small deletions in the conalbumin TATA box drastically decrease specific in vitro transcription.
Nucleic Acids Res 1981;9:1813-1824.
19. Junker-Niepmann M, Bartenschlager R, Schaller H A short cis-acting sequence is required for hepatitis B virus pregenome encapsidation and sufficient for packing of foreign RNA. EMBO J 1990;9:3389- 3396.
20. Tong SP, Li JS, Vitvitski L, Alan K, Christian T Evidence for a base-paired region of hepatitis B virus pregenome encapsidation signal which influ ences the patterns of precore mutations abolishing HBe protein expression. J Virol 1993;67:5651-5655 21. Kim YH, Kang SK, Lee YI. Functional analysis o hepatitis B virus transactivator X: implication of the leucine zipper-like region and C-terminal seven conserved amino acids in functional regions. Bio chem Biophys Res Commun 1993;197:894-903.
22. Wang GH, Zoulim F, Leber EH, Kitson J, Seeger C Role of RNA in enzymatic activity of the reverse transcriptase of hepatitis B viruses. J Virol 1994;68 8437-8442.
23. Chen IH, Huang CJ, Ting LP. Overlapping initiato and TATA box functions in the basal core promote of hepatitis B virus. J Virol 1995;3647-3657.
24. Buckwold VE, Xu Z, Yen TS, Ou JH. Effects of frequent double-nucleotide basal core promoter mu tation and its putative single-nucleotide precursor mutations on hepatitis B virus gene expression and replication. J Gen Virol 1997;78:2055-2065.
25. 김연수, 김성일, 황성규 등. HBeAg 양성인 만성 HBV 보유자에서 HBV core promoter 변이의 임상적 의의 대한소화기학회지 1997:30:769-778.
26. 정성택, 신용준, 김영수, 김진홍, 조성원. B형 만성 간 질환에서 core promoter 변이의 양상. 대한소화기학 회지 1997;31:780-788.
27. Laskus T, Rakela J, Tong MJ, Nowicki MJ, Mosley JW, Persing DH. Naturally occurring hepatitis B virus mutants with deletions in the core promoter region. J Hepatol 1994;20:837-841.
28. Okamoto H, Tsuda F, Akahane Y, et al. Hepatitis B virus with mutations in the core promoter for an antigen-negative phenotype in carriers with antibody to e antigen. J Virol 1994;68:8102-8110.
29. Takahashi K, Aoyama K, Ohno N, et al. The pre core/core promoter mutant (T1762A1764) of hepatitis B virus: clinical significance and an easy method fo detection. J Gen Virol 1995;76:3159-3164.
30. Kidd Ljunggren K, Oberg M, Kidd AH. Hepatitis B virus X gene 1751 to 1764 mutations: implications for HBeAg status and disease. J Gen Virol 1997;78 1469-1478.
31. 박중원, 윤정환, 황유진, 이효석, 김정룡. B형 간염바 이러스 표면항원과 항체가 동시에 발현된 만성 간염 환자에서 표면항원 “a" 결정기 유전자의 변이. 대한소 화기학회지 1997;29:182-191.
32. 송우건, 서검석, 김용성, 김학철. 한국인에서 B형 간 염 바이러스 유전자형 -핵산 배열로 관찰한 B형 간염 바이러스의 C 유전자 부위에서-. 대한간학회지 1998;
4(suppl):S22.
33. Kwok S, Higuchi R. Avoiding false positives with PCR. Nature 1989;339:237-238.
34. Moriyama K, Okamoto H, Tsuda F, Mayumi M Reduced precore transcription and enhanced core- pregenome transcription of hepatitis B virus DNA after replacement of the precore-core promoter with sequences associated with e antigen-seronegative persistent infections. Virology 1996;226:269-280.
35. Fukuda R, Nguyen XT, Ishimura N. et al. X gene and precore region mutations in the hepatitis B viru genome in persons positive for antibody to hepatitis B e antigen: comparison between asymptomatic
"healthy" carriers and patients with severe chronic active hepatitis. J Infect Dis 1995;172:1191-1197.
36. Laskus T, Rakela J, Nowicki MJ, Persing DH. He patitis B virus core promoter sequence analysis in fulminant and chronic hepatitis B. Gastroenterology 1995;109:1618-1623.
Kim, et al. Nucleotide Variability of Core Promoter Region in Type B Chronic Liver Diseases 643
37. 이진우, 김연수, 김영석 등. 만성 B형 간질환에서 HBV enhancer II의 변이. 대한소화기학회지 1998;32 (suppl):222.