556
©The Korean Society of Food Science and Technology
잠재적인 생균제제 Lactobacillus plantarum P1201을 이용한
항산화 활성이 증진된 두유-요구르트 제조
황정은·안민주·이희율·이병원
1·김현태
1·고종민
1·백인열
1·서원택·조계만*
경남과학기술대학교 식품과학부, 1농촌진흥청 국립식량과학원 기능성작물부
Potential Probiotic
Lactobacillus plantarum P1201 to
Produce Soy-yogurt with Enhanced Antioxidant Activity
Chung Eun Hwang, Min Ju An, Hee Yul Lee, Byong Won Lee1, Hyun Tae Kim1, Jong Min Ko1, In Youl Baek1, Weon Taek Seo, and Kye Man Cho* Department of Food Science, Gyeongnam National University of Science and Technology 1Deparment of Functional Crop, National Institute of Crop Science, Rural Development Administration
Abstract In this study, we evaluated changes in physicochemical properties, total phenol and isoflavone contents, and antioxidant activity during soymilk fermentation by a potential probiotic Lactobacillus plantarum P1201. The P1201 strain showed survival rates of 58.14% and 62.22% after 4 h exposure to acid and artificial gastric acid conditions (pH 2.5), respectively. The optimal conditions for soy-yogurt fermentation by L. plantarum were determined to be as follows: temperature of 35oC, seed volume of 5.0%, and sucrose content of 10.0%. The total phenol and isoflavone contents and antioxidant activity were higher in soy-powder milk (SPM) yogurt than soymilk yogurt. In particular, the highest levels of isoflavone-aglycones, such as daidzein, glycitein, and genistein (91.50, 12.34, and 61.39µg/g, respectively), were observed after 48 h of SPM fermentation. Thus, these results suggest that the soy-yogurt extract could be used as a potential source of natural antioxidants in food.
Kewwords: probiotic, Lactobacillus plantarum P1201, soy-yogurt, antioxidant, isoflavone
서
론
생균제제(probiotics)라 함은 인체의 척박한 환경인 위를 통해 장까지 도달하는 살아있는 생균을 뜻한다. 이러한 생균제제는 보 통 자연계에 널리 존재하는 젖산균이며, 이들을 사용하여 다양한 발효식품에 이용되어져 왔으며 오랜 기간에 걸친 섭취 이력을 통 해 일반적으로 안전하다고 인식되는 미생물로 알려져 있다(1). 생 균을 섭취함으로서 젖산균이 분비하는 효소 및 nisin 등의 항균 물질에 의해 위해미생물 억제, 정장작용, 면역증가, 배변활동 개 선 등 여러 효능이 있다(2). 콩은 양질의 단백질, 불포화 지방산 및 식이섬유소가 많아 영 양학적으로 우수한 식량자원이며 영양인자들이 항암 등 여러 생 리활성이 있다고 알려지면서 콩에 대한 관심이 증가하였다(3). 콩 의 단백질은 영양소의 40% 이상을 차지하며 대장암, 간암 및 폐 암의 성장을 억제하는 것으로 보고가 되어있다(4). 콩에는 다양 한 생리활성 물질이 함유되어 있으며, 그 중 대표적으로 isoflavone 은 glycosides (25%), malonylglycosides (70-80%), acetylglycosides (5%) 및 aglycone (2%)의 형태로 존재한다(5). 발효식품이 아닌 경우 소화 흡수 및 생리활성이 높은 aglycone 함량은 약 2% 정 도이지만 발효 중에는 식품에 따라서 30-80%까지 증가한다고 알 려져 있다(6). 두유 요구르트는 기존의 우유 요구르트에 비해 가격이 저렴할 뿐만 아니라 영양도 우수하여 국민 건강기능 향상에 크게 기여 할 수 있다(7). 그러나 콩 특유의 불쾌취와 비소화성 과당류가 함 유되어 있어 섭취 시 소화가 잘되지 않는 단점이 있다. 따라서 풍미와 소화장애 개선을 위해 여러 젖산균을 이용한 두유 발효 연구가 수행되었으며 젖산균 종류에 따라 풍미 및 비소화성 과 당류의 분해가 크게 차이나는 것으로 보고되었다(8). Ko 등(9)은 Lactobacillus plantarum은 비소화성 과당류를 분해하며 생균제로 서의 능력이 우수하다고 보고하였다. Wang 등(10)도 생균제로 쓰 이는 젖산균 중 L. plantarum은 높은 산 생성력으로 두유 요구르 트의 맛과 영양을 배로 늘려줄 뿐 아니라 장내균총의 정상화 등 의 효과가 있다고 보고하였다. 따라서 본 연구에서는 생균제제능이 뛰어난 L. plantarum P1201 을 선발 및 동정하였다. 부가적으로 L. plantarum P1201를 이용 하여 두유 요구르트를 제조하고 발효 중 총 phenolics, isoflavone 함량 및 항산화 활성 변화를 측정하여 기능성 두유 요구르트 제 조에 관한 기초 자료 확보를 위해 수행하였다.*Corresponding author: Kye Man Cho, Department of Food Sci-ence, Gyeongnam National University of Science and Technology, Jinju, Gyeongnam 660-758, Korea
Tel: 82-55-751-3272 Fax: 82-55-751-3279 E-mail: [email protected]
Received May 20, 2014; revised August 31, 2014; accepted September 1, 2014
재료 및 방법
재료, 시약 및 배지
국내산 늘찬콩은 2011년 경상남도 밀양시 농촌진흥청 국립식 량과학원 기능성작물부에서 재배 및 수확된 것을 공급받아 사용 하였다. 미생물 배지는 Lactobacill MRS broth (MRS, Difco Lab-oratories, Detroit, MI, USA)를 사용하였다. 12개의 isoflavone 중 3종의 aglycone (daidzein, glycitein 및 genistein)은 Sigma-Aldrich 사(Saint Louis, MO, USA)에서 구입하였고 glycosides (daidzin, glycitin 및 genistin)는 Indofine (Hillsborough, NJ, USA)사에서 구 입하였다. 3종의 malonylglycosides 및 acetylglycosides (malonyl-daidzin, malonylglycitin, malonylgenistin, acetyl(malonyl-daidzin, acetylgly-citin, 및 acetylgenistein)는 LC Laboratories (Woburn, MA, USA) 사에서 구입하였다. Glacial acetic acid, folin-cialteu reagent, 2,2-diphenyl-1-picrydrazyl (DPPH), 2,4,6-azinobis(3-ethyl-benzothiazo-line-6-sulfonic acid) (ABTS) 및 2,4,6-tripyridyl-s-triazine (TPTZ) 는 Sigma-Aldrich사 제품을 사용하였다. Methanol, acetonitrile 및 water는 J. T. Baker사(Philipsbug, NJ, USA)에서 구입하여 사용하 였고 그 외 나머지 기타 시약은 필요에 따라 특급 혹은 1급을 구입하여 사용하였다. 젖산균 분리 및 생균제제능 확인 젖산균 분리는 김치 등 발효식품을 멸균생리식염수로 단계별 희석하여 분리하였다. 순수 분리된 젖산균은 MRS 액체배지에 접 종하여 30oC에서 48 h 배양하였고 20% (v/v) glycerol를 첨가하여 −70oC 냉동고에서 보관하며 필요에 따라 활성화시켜 사용하였다. 산 및 인공위액산 내성 측정은 Kang과 Lee(1)의 방법을 변형 하여 수행하였다. 산 내성은 pH를 2.0-3.0으로 조정한 50 mL MRS 액체배지를 사용하였으며 인공위액산 내성은 pH를 2.0-3.0 으로 조정한 후 펩신 1.0%를 첨가한 50 mL MRS 액체배지를 사 용하였다. 각각의 배지에 종균 배양액을 106 CFU/mL 농도로 접 종한 후 37oC에서 4 h 배양시켰으며 각각 0, 2, 및 4 h째 MRS 평판배지를 이용하여 생균수를 측정하였다. 생존율은 초기 균의 농도와 MRS 평판배지 상에 형성된 젖산균의 집락수 비율을 비 교하여 계산하였다. 담즙산 내성은 Ha 등(11)의 방법을 변형하 여 측정하였다. MRS 액체배지에 30oC에서 48 h 배양한 균체(106 CFU/mL)를 다른 농도(1.0-4.0% w/v)의 담즙을 함유한 MRS 평 판배지 상에 spotting 하였다. 그 후 37oC에서 48 h 배양하였으며 젖산균에 대한 담즙산의 최소억제농도는 spot을 육안검사로 판 정함으로써 spot에서 균 생장을 전적으로 저해시키는 최소농도 로 결정하였다. 균주 P1201의 동정 균주 동정은 Cho 등(12)의 방법에 준하여 형태학적, 생리학적, 세포 지방산 조성, 생화학적 및 분자유전학적 특성으로 동정하였 다. 형태학적 특성은 그람염색 후 광학 현미경으로 관찰하였다. 생리학적 특성은 배양온도(10-50oC), pH (3-11) 및 NaCl
농도(0-10%)에서 확인하였다. 세포 지방산 조성은 MIDI Microbial Iden-tification System (Hewlett Packard, Palo Alto, CA, USA) 방법에 따라 전처리 후 지방산을 분석하였다. 지방산 분석은 gas chro-matography (model 7890, Hewlett Packard) 및 phenylmethyl silicone fused silica capillary column (25 m×0.2 mm, Hewlett Packard)을 사 용하였다. 생화학적 특성은 API50 NE kit (BioMeriux, Marcy-I'Eltoile, France)를 사용하여 당 이용성을 측정하였다. 한편 균주 P1201은 MRS 액체배지에서 24 h 배양한 후 Intron Genomic
DNA Purification kit (Intron Botechnology Co., Suwon, Korea) 를 사용하여 genomic DNA를 분리하고 이를 주형으로 PCR을 수 행하여 16S rDNA 염기서열을 결정하였다. PCR 반응은 95oC에
서 denaturation 5 min, 49oC에서 annealing 30 s, 72oC에서
exten-sion 1 min 30 s 동안 30 cycle을 수행하였다. Primer 제작은 세균 의 16S rDNA conserved region에서 약 1,500 bp 정도가 되게 제 작을 하였다. Forward oligonucelotide primer는 5'-CGGAGAGTTT GATCCTGG-3', reverse oligonucelotide primer는 5'-TACGGCTAC CTTACGAC-3'을 사용하였다(12). PCR 반응이 끝난 후 전기 영 동하여 16S rRNA 단편을 확인하고 Total Fragment DNA Purifi-cation Kit (Intron Biotechnology)의 기술된 방법에 따라 gel에서 DNA를 분리하였다. 정제한 16S rRNA 단편을 주형으로 염기서 열을 결정하였다. 결정된 염기서열 16S rRNA는 GenBank data-base (http://www.ncbi.nlm.nih.gov)로부터 얻은 또 다른 균주의 16S rRNA와 비교 분석하였다. 16S rRNA 유사성 값은 DNAMAN analysis system (Lynnon Biosoft, Quebec, Canada)을 사용하여 alignments 및 evolutionary distance로부터 계산하였다. Phyloge-netic tree는 neighbour-joining method와 distance matrix data를 사 용하여 확인하였다.
두유 제조 및 젖산발효
일반두유(soymilk, SM)는 정선 및 수세한 콩을 실온에서 12 h 수침하고 30 min 동안 물기를 제거한 후 수침콩 400 g에 정제수 1,000 mL를 가하고 휴롬 원액기(HUROM HU-400/HU-400G, Hurom Group Co., Ltd., Gimhae, Gyeongnam, Korea)를 이용하 여 액상과 비지를 걸러낸 후 두유 1,000 mL에 sucrose 100 g을 첨가한 후 250 mL 유리병에 담아 121oC에서 15 min 살균하여 제
조하였다. 콩-분말 두유(soy-powder milk, SPM)는 콩을 초밀도 분 쇄기(HD07026-5003, Hyundae Household Appliances Co., Ltd., Seoul, Korea)로 분쇄하여 80 mesh의 체에 걸러 콩 분말을 얻었 다. 콩 분말 100 g에 정제수 1,000 mL를 혼합한 후 sucrose 100 g 을 첨가하여 250 mL 유리병에 담아 121oC에서 15 min 살균하여 제조하였다. 살균된 두유는 상온으로 냉각하여 종균 배양액을 5.0% (v/v) 접종하여 35oC에서 48 h 발효하였으며 발효 중 12 h 간격으로 시료를 채취하여 실험에 사용하였다. SM 및 SPM의 건 조중량(고형분) 함량은 선행연구를 통해 동일한 양으로 조정하였다. pH, 총산 및 생균수 변화
pH는 pH meter (model 3510, Jenway, Essex, UK)를 사용하여 측정하였다. 총산은 중화적정법으로 시료 1 mL를 pH 8.2±0.1까 지 중화시키는데 소비된 0.1 N NaOH의 양을 구하고 젖산(lactic acid)양으로 환산하였다. 생균수는 멸균생리식염수로 단계별 희 석하여 MRS 평판배지에 도말한 후 30oC에서 48 h 배양 후 나 타난 집락을 계수하였다. 각 실험은 3회 반복하여 평균값으로 나 타내었다. 추출물 제조 시료 추출은 Hwang 등(13)의 방법에 준하여 추출하였다. 즉, 동결건조 분말 1 g에 10배의 50% 메탄올을 가하고 상온에서 12 h 추출하였다. 추출물은 0.45 µm membrane filter (Dismic-25CS, Toyoroshikaisha Ltd., Tokyo, Japan)로 여과하여 총 phenolics와 isoflavone 함량 및 항산화 활성 측정을 위한 시료로 사용하였다. 총 phenolics 함량 측정
희석한 추출물 0.5 mL를 시험관에 분주하고 25% Na2CO3 용액
0.5 mL를 첨가하여 3 min 정치시켰다. 다시 2 N folin-ciocalteu 시 약 0.25 mL를 첨가하여 혼합한 다음 30oC에서 1 h 동안 정치시
켜 발색시켰다. 발색된 청색을 750 nm에서 분광광도계(Spectronic 2D, Thermo Electron Co., Waltham, MA, USA)를 사용하여 흡광 도를 측정하였다. 이때 총 phenolics 함량은 gallic acid를 이용하 여 작성한 표준곡선으로부터 함량을 구하였다. 각 실험은 3회 반 복하여 평균값으로 나타내었다.
Isoflavone 함량 분석
Isoflavone 함량 분석은 Cho 등(15)의 방법에 준하여 high per-formance liquid chromatography (HPLC, Agilent 1200 series, Agilent Co, Forest City, CA, USA)로 분석하였다. 분석 column은 Lichrophore 100 RP C18 column (4.6×250 mm, 5µm, Merck, Germany)을 사용하였고 이동상 용매는 0.2% glacial acetic acid in water (solution A)와 0.2% acetonitrile in glacial acetic acid (solution B)로 분석하였다. 이동상 조건은 A 용매 기준으로 0 min-100%, 15 min-90%, 25 min-80%, 35 min-75%, 45 min-65% 및 50 min-65%로 유지하였다. 시료는 20 µL를 주입하였으며, 이동상 속도는 30oC에서 1 mL/min으로 유지하였다. 검출기는 diode array
detector를 사용하여 UV 254 nm에서 검출하였다. 각 실험은 3회 반복하여 평균값으로 나타내었다. DPPH 라디칼 소거 활성 DPPH 라디칼 소거 활성은 Hwang 등(13)의 방법을 변형하여 측정하였다. 1.5×10-4 MDPPH 에탄올 용액 0.8 mL과 적당히 희 석한 여과액 0.2 mL를 가하고 10초간 균질화 시켜 암실에서 30 min 방치 후 분광광도계(Spectronic 2D)를 사용하여 525 nm에서 흡광도를 측정하였다. 음성대조구 실험은 시료대신에 추출용매 0.2 mL를 취하여 실험하였으며 실험구와 음성대조구의 흡광도를 구하여 백분율(%)로 나타내었다. 각 실험은 3회 반복하여 평균값 으로 나타내었다. ABTS 라디칼 소거 활성 ABTS 라디칼 소거 활성은 Hwang 등(13)의 방법에 준하여 측 정하였다. 7 mM ABTS5 mL과 2.45 mM K2S2O8 5 mL를 섞어 암 실에서 12-16 h 방치 시킨 후 메탄올과 1:88 비율로 섞어 732 nm 에서 대조구의 흡광도 값이 0.7±0.02가 되도록 조절한 ABTS+ 용 액을 사용하였다. 적당히 희석된 여과액 0.1 mL과 ABTS+ 용액 0.9 mL를 첨가하여 혼합한 후 3 min 정치 시켜 732 nm에서 분광 광도계를 사용하여 흡광도를 측정하였다. 음성대조구 실험은 시 료대신에 추출용매 0.1 mL를 취하여 실험하였으며, 실험구와 음 성 대조구의 흡광도를 구하여 백분율(%)로 나타내었다. 각 실험 은 3회 반복하여 평균값으로 나타내었다.
Ferric reducing ability of plasma (FRAP) assay
FRAP는 Hwang 등(13)의 방법에 준하여 측정하였다. Acetate buffer (30 mM, pH 3.6), TPTZ 시약(10 mM in 40 mM HCl), 및 FeCl3 용액(20 mM in DW)을 10:1:1 (v/v/v)의 비율로 혼합하여 FRAP 측정 시약을 조제 하였다. 37oC 항온기에서 15 min 예비반 응 후 시료 50 µL와 FRAP 시약 950 µL를 시험관에 분주하여 37oC에서 15 min 반응시키고 분광광도계를 사용하여 590 nm에서 흡광도를 측정하였다. 음성대조구 실험은 시료대신에 추출용매를 사용하였다. 각 실험은 3회 반복하여 평균값으로 나타내었다. 통계처리
실험 결과는 SPSS 12.0 package (SPSS Institute, Chicago, IL, USA)를 사용하여 분산 분석을 수행하였고 평균±표준편차로 나 타내었다. 각 시료 분석 결과에 대한 유의성 검정은 분산 분석 후 p<0.05 수준에서 Duncan’s multiple range test를 실시하였다.
결과 및 고찰
젖산균의 생균제제능 확인 김치 등과 같은 발효식품으로부터 약 300여 종의 다양한 젖산 균을 순수분리한 후 산, 인공위액산 및 담즙산 내성이 우수한 10 종의 젖산균을 선발한 결과 Table 1과 같았다. 산 내성은 젖산균 중 S48 및 P1201 균주가 가장 우수하였다. S48 균주는 pH 2.5에 서 2 및 4 h 경과 후 91.26 및 63.25%의 생존율을 나타내었고 P1201 균주는 84.63 및 58.14%의 생존율을 나타내어 S48 균주가 우수한 것으로 나타났다. 인공위액산 내성 역시 S48 및 P1201 균 주가 가장 우수하였으며 S48 균주는 pH 2.5에서 2 및 4 h 경과 Table 1. Survival of lactic acid bacteria isolated under acidic and artificial gastric acidic conditionsIsolates
Survival rate1) (%)
Acid conditions (pH 2.5) Artificial gastric conditions (pH 2.5) Incubation time (h) Incubation time (h)
2 4 2 4
S48 91.26±4.52a 63.25±3.16a 88.52±4.43a 59.57±2.98a
S52 80.51±4.02b 52.31±2.62ab 59.66±2.98 44.21±1.78b
S56 78.92±3.16b 50.34±2.52b 48.22±1.93d 30.33±1.82c
S64 66.74±2.67c 47.45±1.90bc 46.62±2.33d 27.99±1.40d
S65 64.53±3.22c 45.21±2.11bc 72.33±3.62bc 49.41±2.47b
P1201 84.63±2.53ab 58.14±2.33a 92.56±4.63a 62.22±3.73a
K23 77.24±3.09b 50.46±3.03b 60.45±2.42c 42.25±2.11b
K24 79.23±3.17b 51.33±3.08b 74.98±3.00bc 47.32±2.37b
D56 63.88±3.19c 42.31±1.69c 80.45±4.02b 53.47±2.67ab
D58 81.69±4.08b 53.65±2.68ab 84.22±4.21ab 55.67±3.34ab
1)Each isolated strains was tested in triplicate for its tolerance in acidified and artificial gastric acidified MRS broth. Means with different lowercase letters (a, b, and c) indicate significant differences of incubation times by Tukey’s multiple range test (p<0.05).
후 88.52 및 59.57%의 생존율을 나타내었고 P1201 균주는 92.56 및 62.22%의 생존율을 나타내어 S48 균주보다 우수한 것으로 나 타났다(Table 1). 한편 담즙산 내성은 배양 후 형성된 spot을 육 안으로 살펴본 결과 10종의 균주 모두 약간의 차이는 있었으나 대부분 생장 하는 것으로 나타났다. 특히 3.0-4.0% 농도의 경우 담즙을 첨가하지 않은 대조구와 비교 시 큰 차이를 나타내지 않 아 균주 모두 담즙산에 의해 사멸되지 않고 성장이 저해되는 것 으로 판단되었다(자료 제시하지 않음). 인간의 위장관에 자연적으로 존재하는 생균제제는 인체에서 영 양소가 부족한 위, 췌장, 및 십이지장 등의 환경 속에서 생존율 이 뛰어나다. 생균제와 관련된 몇몇 연구에서는 생균제제가 인간 의 건강과 장수에 긍정적인 영향을 미치는 것으로 나타났고(16) 장내에 많이 분포하는 젖산균들은 여러 스트레스 조건에서 생존 하기 위해 경쟁적으로 증식하는 것으로 알려져 있다(17,18). Liu 등(19)은 L. plantarum 균주가 L. casei 균주보다 산 및 담즙산 내 성이 우수하다고 보고한 바 있으며 인공 산성 환경에서 3 h 경 과 후 잔존 균수는 107-108 CFU/mL 수준이었고, 인공 담즙산(1-2%)은 16 h 경과 105.5 CFU/mL 수준까지 증식이 가능함을 보여 주었다. Nagata 등(20)도 L, plantarum 균주가 Leuconostoc 속보다 인공위액산 내성이 우수함을 보여주었고 본 연구 결과와도 유사 하였다. 이러한 이전 연구 결과들과 김치 발효 후기(pH 2-3)에 L. plantarum 균주가 존재한다는 점(21)으로 보아 내산성이 뛰어난 균주이며 일반적으로 산, 인공위액산 및 담즙산에 대해 다른 미 생물보다 생존율이 뛰어난 것으로 판단되었다. 선발 P1201 균주의 확인 균주 P1201의 동정은 형태학적, 생리학적, 세포 지방산 조성, 생화학적, 및 분자유전학적 특성으로 동정을 한 결과 Fig. 1 및 Table 2와 같았다. 형태학적 특성은 그람양성 간균이었고 생리학적 특성은 온도 10-40oC, pH 5-11, 및 NaCl 4%까지 생육이 가능하였다(Table 2). 세포 지방산 조성은 Lactobacillus속에 많이 분포하는 C16:0 (37.81%)이 주요 지방산인 것으로 나타났다(자료 제시하지 않음). 한편 당 이용성 결과 L-arabinose를 비롯하여 lactose, D-galactose, Table 2. Phenotypic characteristics of L. plantarum P1201 strain
Characteristics Reaction Characteristics Reaction
Morphology Ribose +
Shape Rod D-Xylose
-Gram stain +1) L-Xylose
-Cell dimension 2-8 µm Adonitol -Physiological properties β-Methyl-xyloside -Aerobic growth + Galactose +
Growth at D-Glucose + 10oC w D-Fructose + 20oC + D-Mannose + 25oC + L-sorbose -30oC + Rhamnose -35oC + Dulcitol -40oC + Inositol -50oC - Mannitol +
Growth in NaCl Sorbitol +
0% + a Methyl-D-mannoside -2% + a Methyl-D-glucoside -4% w N Acetyl glucosamine + 6% - Amygdaline + 8% - Arbutine + 10% - Esculine + pH Salicine + 3 - Cellobiose + 5 + Maltose + 7 + Lactose + 9 w Melibiose + 11 w Saccharose + Carbohydrates Trehalose + Control - Inuline + Glycerol - Melezitose + Ertythritol - D-Raffionse -D-Arabinose - Amidon -L-Arabinose + Glycogene -Xylitol - L-Fucose -β Gentiobiose + D-Arabitol -D-Turanose + L-Arabitol -D-Lyxose - Gluconate + D-Tagatose - 2 ceto-gluconate -D-Fucose - 5 ceto-gluconate -1)Symbol: +, positive reaction; -, negative reaction; w, weak reaction. Fig. 1. Phylogenetic relationships of the starin P1201 and other
closely related Lactobacillus plantarum species based on 16S rDNA gene sequences. Numbers above each node are confidence levels (%) generated from 1,000 bootstrap trees.
D-glucose, D-fructose 및 D-mannose 등을 이용할 수 있었다(Table 2). 정확한 균주 동정을 위하여 16S rRNA 염기서열 분석을 수행 하였다. P1201 균주의 염기서열을 결정하여 phylogenetic tree 분 석 결과 L. plantarum WCFS1 (AL935263)와 99.0%로 가장 유사 성이 높았다(Fig. 1). 형태학적, 생리학적, 세포 지방산 조성, 생화 학적 및 분자유전학적 특성을 비교 검토하여 최종적으로 L. plantarum P1201로 명명하였다. 두유 요구르트 발효 중 이화학적 특성 변화 배양온도(20-35oC), 종균 접종량(1.0-5.0%, v/v), 당 종류(sucrose,
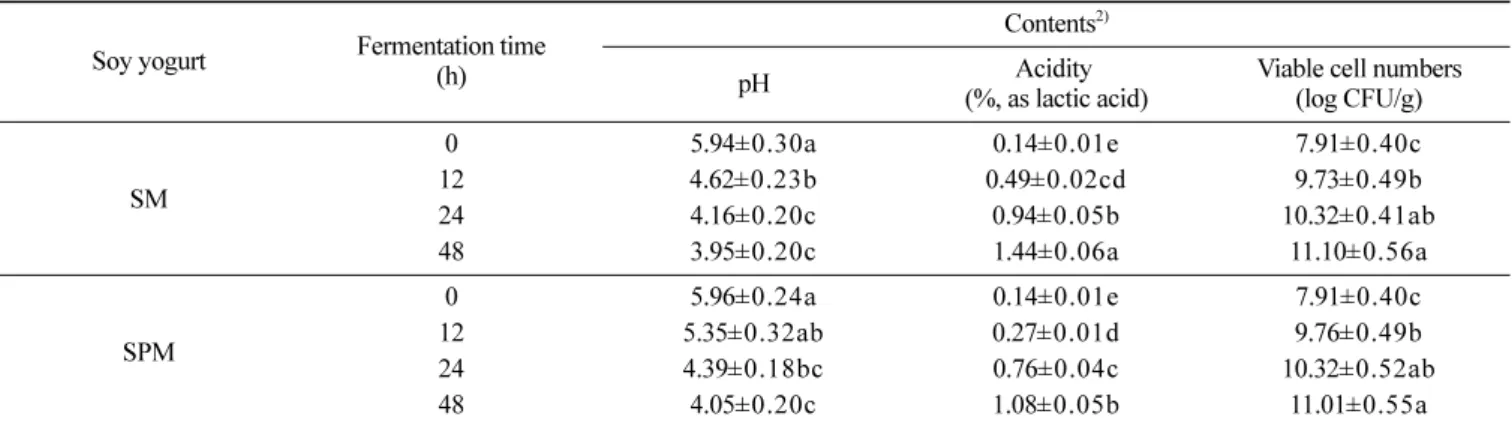
lactose, fructose 및 glucose) 및 당 농도(0-15.0%, w/v)에 따른 두 유 요구르트 발효조건을 검토한 결과 향, 맛, 질감 등의 관능적 성질, 이화학적 특성 및 항산화 활성이 우수한 조건은 배양온도 35, 종균 접종량 5.0% 및 10% sucrose 이었다(자료 제시하지 않 음). 최적 조건에 따라 제조된 SM 및 SPM 요구르트 발효 중 pH, 총산 및 생균수 변화는 Table 3과 같았다. SM 요구르트는 발효 12 h 경과 후 pH가 급격히 감소하였고 SPM 요구르트는 발효가 진행됨에 따라 서서히 감소하는 경향을 보였으나, 발효종기 생균 수는 유사한 수준이었다. 발효 초기의 pH는 5.94 (SM) 및 5.96 (SPM) 수준이었고 발효 48 h 경과 후 3.95 (SM) 및 4.05 (SPM) 로 감소하였다. 한편 총산은 pH 감소에 상응하여 발효 48 h 경 과 후 1.44 (SM) 및 1.08% (SPM)로 증가하였다(Table 3). 적절한 산의 농도는 발효두유의 품질을 높이는 데 가장 중요 한 요소 중 하나로 알려져 있다(22). Chun 등(23)은 Streptococcus 속과 Weissella 속을 혼합 배양한 두유 요구르트 발효 시 12 h만 에 pH가 4.19로 감소하였고 총산은 0.64%까지 도달하였다. Pyo 등(24)은 젖산균(L. plantarum, L. delbrueckii, Bifidobacterium breve, 및 B. thermophilum) 단독 배양 시 발효 24 h만에 pH 4.4-5.8 이었고 총산은 0.76-1.12%이었다. 이러한 연구 결과들은 본 연구의 산 생성력과 유사하였고 시판두유의 pH가 4.2-4.4 수준 (25)인 점에 의거하면 본 연구에 사용된 L. plantarum P1201 균 주는 산 생성능이 우수한 것으로 판단된다. 두유 요구르트 발효 중 총 phenolics 함량 SM 및 SPM 요구르트 발효 중 총 phenolics 함량 변화는 Fig. 2와 같았다. 발효가 진행됨에 따라 전체적으로 총 phenolics 함량 은 증가하였고 SPM 요구르트는 SM 요구르트보다 높은 함량을 나타내었다. SM 요구르트의 총 phenolics 함량은 발효 초기 1.18 mg/g, 발효 12 h 경과 1.24 mg/g, 발효 24 h 경과 1.27 mg/g 및 발 효 48 h 경과 1.36 mg/g으로 증가하였다. 한편 SPM 요구르트는 발효 초기 1.33 mg/g, 발효 12 h 경과 1.40 mg/g, 발효 24 h 경과 1.49 mg/g 및 발효 48 h 경과 1.51 mg/g을 나타내었다(Fig. 2). Phenolics 화합물은 보통 식물계에 널리 존재하고 수산기와 당 이 결합한 형태로 인체에 다양한 생리활성을 나타내며(13) 항산 화 활성에 관여하는 것으로 알려져 있다(26). Mccue와 Shetty(27) 는 Keflr를 첨가한 두유 발효 중 총 phenolics 함량과 항산화 활 성이 발효 24 h까지 증가하였지만 48 h에는 총 phenolics 함량만 감소한다고 하여 본 연구 결과와는 차이가 있었다. Wang 등(28) 은 발효가 진행됨에 따라 두유의 항산화 활성 및 환원력이 증가 한다고 보고하여 본 연구결과와 유사하였다. 한편 Cho 등(29)은 이와 유사한 콩 발효식품인 청국장에서 총 phenolics 함량이 증 가한다고 보고하였는데 본 연구와는 함량 차이가 비교적 크게 나 타났다. 이러한 차이는 원료 콩, 발효과정, 배양환경, 및 발효미 생물의 효소 활성차이에 의한 것이라 판단되었다.
Fig. 2. Changes of total phenolic contents during soy milk (SM) and soy powder milk (SPM) fermentations at 35oC for 48 h. All values are presented as the mean±SD of triplicate determination. Means with different lowercase letters (a and b) indicate significant differences of fermentation times by Tukey’s multiple range test (p<0.05).
Table 3. Changes of pH, acidity and viable cell numbers during soy milk (SM) and soy powder milk (SPM) fermentations1)
Soy yogurt Fermentation time (h)
Contents2)
pH Acidity
(%, as lactic acid)
Viable cell numbers (log CFU/g) SM 0 5.94±0.30a 0.14±0.01e 7.91±0.40c 12 4.62±0.23b 0.49±0.02cd 9.73±0.49b 24 4.16±0.20c 0.94±0.05b 10.32±0.41ab 48 3.95±0.20c 1.44±0.06a 11.10±0.56a SPM 0 5.96±0.24a 0.14±0.01e 7.91±0.40c 12 5.35±0.32ab 0.27±0.01d 9.76±0.49b 24 4.39±0.18bc 0.76±0.04c 10.32±0.52ab 48 4.05±0.20c 1.08±0.05b 11.01±0.55a
1)SM and SPM were fermented at 35 for 48 h
2)All values are means of determinations in three independent experiments. Means with different lowercase letters (a, b, and c) indicate significant differences of fermentation times by Tukey’s multiple range test (p<0.05).
두유 요구르트 발효 중 isoflavone 함량 변화
SM 및 SPM 요구르트 발효 중 HPLC 크로마토그램 및 isofla-vone 함량 변화는 Fig. 3 및 Table 4와 같았다. HPLC 크로마토 그램은 총 11개의 isoflavone 유도체(daidzin, glycitin, genistin, malonyldaidzin, malonylglycitin, malonygenistin, acetyldaidzin, acetylglycitin, daidzein, glycitein 및 genistein)가 검출되었다. SM 및 SPM 요구르트의 발효 초기 공통적인 피크로는 배당체(glyco-sides)인 diadzin (peak 1), glycitin (peak 2) 및 genistin (peak 3) 이 검출되었고 발효과정 중 isoflavone 피크 변화는 glycosides가 줄어들면서 비배당체(aglycone)인 daidzein (peak 9), glycitein (peak 10) 및 genistein (peak 11)의 함량이 증가하였다(Fig. 3).
한편 총 isoflavone 함량은 약간 감소하였으나 SPM 요구르트 가 SM 요구르트보다 약 2배정도 높았으며 aglycone 함량 또한 높은 경향을 나타내었다(Table 4). SM 요구르트의 총 isoflavone
함량은 발효 초기(0 h) 524.5 µg/g에서 발효 48 h 후 약간 감소하 여 502.2 µg/g이었고 isoflavone-aglycone 형태(daidzein, glycitein 및 genistein)는 35.79 µg/g에서 약 3배 증가한 109.8 µg/g이었다. 또한, SPM 요구르트의 총 isoflavone 함량은 1,161 µg/g(발효 0 h) 에서 1,083 µg/g(발효 48 h)으로 약간 감소하였고 isoflavone-agly-cone 형태 역시 발효 초기 85.23 µg/g에서 발효 종기(48 h) 165.2 µg/g으로 약 2배 정도 증가하였다(Table 4). 이 결과로부터 비배 당체인 aglycone 전환율은 SM 방식이 SPM 방식보다 높은 것을 알 수 있었으며, 이는 SPM 발효 방식보다는 SM 발효 방식이 산 생성력이 더 우수하기 때문인 것으로 판단되었다(Table 3).
일반적으로 콩의 isoflavone은 약 90%가 glycoside, malonylgly-coside, 및 acetylglycoside 형태로 존재하며 2-10%가 aglycone으로 존재한다(30). 이러한 화합물들은 발효과정 중 미생물이 생성하 는 산이나 β-glucosidase 활성에 의해 aglycone으로 전환된다(15). Fig. 3. Comparison of HPLC chromatogram of soy yogurt isoflavone. Samples: soy yogurt fermented at 35oC for 48 h with SM and SPM. A, SM 0 h; B, SM 12 h; C, SM 24 h; D, SM 48 h; E, SPM 0 h; F, SPM 12 h; G, SPM 24 h; H, SPM 48 h. 1, daidzin; 2, glycitin; 3, genistin; 4, malonyldaidzin; 5, malonylglycitin; 6, acetyldaidzin; 7, acetylglycitin; 8, malonylgenistin; 9, daidzein; 10, glycitein; 11, genistein
한국 식품과 학회지 제 46 권 제 5 호 (2014)
Table 4. Changes of isoflavone contents during soy milk (SM) and soy powder milk (SPM) fermentations1)
Soy-yogurt
Fermentation time (h)
Isoflavone contents2) (µg/g)
Glycosides Malonylglycosides Acetylglycosides Aglycones
Totals
Daidzin Glycitin Genistin Daidzin Glycitin Genistin Daidzin Glycitin Genistin Daidzein Glycitein Genistein
SM 0 113.50 ±5.68cd 97.30 ±3.90d 170.83 ±8.54d 28.14 ±1.41c 13.11 ±0.79c 64.14 ±3.85ab nd 3) 1.66 ±0.08cd nd 12.83 ±0.64e 10.19 ±0.51c 12.77 ±0.64e 524.47 12 123.44 ±4.94c 128.17 ±4.93c 219.26 ±10.96c 21.89 ±1.31d 13.11 ±0.66c 54.06 ±2.16c nd 2.01 ±0.12c nd 35.79 ±2.15d 9.51 ±0.48c 22.62 ±1.36d 629.86 24 79.96 ±4.00d 121.89 ±6.09c 168.99 ±10.14d 24.95 ±1.25cd 13.66 ±0.55c 60.30 ±3.62ab nd 1.90 ±0.09c nd 52.47 ±2.62c 6.93 ±0.42cd 33.95 ±1.70c 565.00 48 53.92 ±2.70e 106.71 ±5.34cd 131.21 ±6.56e 26.84 ±1.07c 13.25 ±0.66c 59.05 ±2.36b nd 1.46 ±0.07d nd 62.38 ±3.12b 4.50 ±0.23d 42.88 ±2.14b 502.20 SPM 0 237.77 ±9.51a 269.57 ±10.78ab 389.51 ±15.58a 52.34 ±2.62a 33.90 ±2.03a 87.74 ±4.39a tr4) 5.32 ±0.32a nd 27.14 ±1.63d 29.30 ±1.47a 28.79 ±1.44cd 1161.38 12 225.00 ±11.25a 296.75 ±11.87a 393.25 ±23.60a 39.26 ±1.94b 28.41 ±1.42ab 66.60 ±2.66ab tr 4.85 ±0.24a nd 36.48 ±1.46d 26.61 ±1.33a 31.09 ±1.87c 1148.30 24 174.03 ±6.96b 252.16 ±10.09b 347.63 ±17.38b 35.21 ±1.76bc 25.47 ±1.53b 60.88 ±3.04ab tr 4.13 ±0.20a nd 60.68 ±3.64b 20.06 ±1.20b 44.42 ±2.22b 1024.67 48 141.80 ±7.09bc 320.47 ±19.23a 336.60 ±16.83b 31.90 ±1.60bc 25.79 ±1.29b 57.62 ±3.46b tr 3.63 ±0.22b nd 91.50 ±3.66a 12.34 ±0.62c 61.39 ±3.68a 1083.04
1)SM and SPM were fermented at 35 for 48 h
2)All values are presented as the mean±SD of triplicate determination. Means with different lowercase letters (a, b, c, d, and e) indicate significant differences of fermentation times by Tukey’s multiple range test (p<0.05)
3)nd: not detected 4)tr: trace (< 0.002 µg/g).
특히, Donkor와 Shah(31)는 L. acidophilus, L. casei 및 Bifido-bacterium lactis를 이용한 두유발효 중 β-glucosidase 활성에 비례 하여 aglycone 형태는 증가하고 glycoside 형태는 감소한다고 보
고하였다. 젖산균을 이용한 여러 두유발효 연구에서도 일반적으 로 β-glucosidase 활성에 의해 aglycone 형태로 전환된다는 점을 강조하였다(31-33). 한편 Marazza 등(34)과 Wei 등(35)은 β-glu-cosidase 활성을 나타내진 않으나 젖산균이 생성한 젖산에 의해 glycoside 형태가 aglycone 형태로 전환된다고 하였다. 본 연구에 서 두유 요구르트 발효 중 aglycone 함량이 증가한 것은 균주 P1201이 생성한 산에 의해 배당체 isoflavone이 비배당체 aglycone 으로 전환된 것으로 판단되었으며, 앞으로 발효 중 β-glucosidase 활성 연구가 필요할 것으로 판단되었다. 두유 발효 중 항산화 활성 변화 SM 및 SPM의 요구르트 발효 중 DPPH와 ABTS 라디칼 소거 활성 및 환원력은 Fig. 4와 같았다. SM 및 SPM 요구르트의 DPPH 라디칼 소거활성은 발효초기 각각 28.10% 및 34.40% 이었 으며 발효가 진행됨에 따라 점차적으로 증가하여 발효 48 h 경과 각각 36.53% 및 41.61%를 나타내었다(Fig. 4A). ABTS 라디칼 소 거활성도 발효초기 각각 40.59% 및 50.35%에서 발효 48 h 경과 각각 57.80% 및 60.67%로 활성이 증가하였다(Fig. 4B). 한편 환 원력 역시 라디칼 소거활성과 유사하게 발효초기 각각 0.34 및 0.40에서 발효 48 h 경과 0.39 및 0.50으로 증가하였다(Fig. 4C). 발효 중 항산화 활성은 SPM 요구르트가 SM 요구르트보다 높 았고 이는 콩 수침 시 수용성 phenolics 등의 소실에 기인한 것 으로 판단되었고 수용성 phenolics뿐만 아니라 영양성분의 감소 도 SM 요구르트가 SPM 요구르트보다는 클 것으로 사료되었다. Tyug 등(36)은 콩 분말, 두유분말 및 간장분말을 비교한 결과 콩 분말이 전체적으로 총 phenolics와 isoflavone 함량 및 항산화 활 성이 우수하였다. 그 이유는 콩 껍질부분에 생리활성 물질들이 많이 존재하기 때문이라 보고하였다. Erdil 등(37)도 콩을 물과 함 께 마쇄할 경우 수용성 비타민 및 여러 항암성분들이 소실되므 로 콩 분말을 첨가하는 것이 여러 생물학적관점에서 우수하다고 보고하였다. 이렇듯 두유발효에 관한 여러 연구 결과에서는 대체 적으로 분말형태의 첨가가 색, 조직, 질감 및 항산화 활성이 우 수한 것으로 보고하였다(38-40). DPPH 및 ABTS 라디칼 소거활 성은 phenolics 함량과 서로 상관관계가 있으며 일반적으로 phe-nolics 함량이 높을수록 항산화 활성이 증가하는 것으로 알려져 있다(41-42). Free radical을 효과적으로 억제하는 flavonoid의 활 성은 생리활성 물질 구조와 직접적인 연관이 있는 것으로 알려 져 있다(43). 한편 flavonoid류인 isoflavone은 생리활성이 높은 물 질이며 특히, aglycone 형태인 genistein 및 daidzein은 높은 항산 화 활성을 지닌 것으로 알려져 있다(44). 본 연구에서 SPM 요구 르트의 경우 콩 껍질에서 유래된 총 phenolics, genistein 및 daid-zein 함량에 기인하여 항산화 활성이 SM 요구르트보다 높은 것 으로 판단되었다. 본 연구에서는 생균제제능이 우수한 L. plantarum P1201을 이 용한 SPM 요구르트는 높은 산 생성력으로 품질을 높일 뿐 아니 라 상당량의 aglycone이 생성되어 잠재적인 천연 항산화제 식품 으로 사용 가능하며 향후 상업적인 측면에서 제조 원가 절감 및 공정의 간편화 등 여러 효과가 있을 것으로 사료된다. 한편 본 연구에서 개발된 두유 요구르트 제조에 관한 연구는 현대인의 장 건강에 도움이 되는 새로운 기능성 제품 개발을 위한 기초 자료 로 활용될 수 있을 것이다.
요
약
잠재적인 생균제제 L. plantarum P1201의 두유 발효 중 이화 Fig. 4. Changes of DPPH (A), ABTS (B) radical activities andFRAP assay (C) during soy milk (SM) and soy powder milk (SPM) fermentations at 35oC for 48 h. All values are presented as the mean±SD triplicate determination. Means with different lowercase letters (a, b, c, and d) indicate significant differences of fermentation times by Tukey’s multiple range test (p<0.05).
학적 특성, 총 phenolics과 isoflavone 함량 및 항산화 활성 변화 를 연구하였다. 균주 P1201는 pH 2.5의 산 및 인공위액산 환경 에서 4 h 후 각각 58.14%와 62.22%의 생존율을 나타내었다. 두 유 요구르트의 최적 발효 조건은 배양온도 35oC, 종균 접종량 5.0% 및 10%의 sucrose이었다. 한편 총 phenolic와 isoflavone 함 량 및 항산화 활성은 SM 요구르트보다 SPM 요구르트가 우수하 였다. 특히 SPM 발효 48 h 후, daidzein, glycitein 및 genistein의 함량은 각각 91.50, 12.34 및 61.39 µg/g이 검출되었다. 이러한 결과로 본 연구의 두유 요구르트 추출물은 잠재적인 천연 항산 화제로 식품에서 사용할 수 있을 것으로 판단되었다.
감사의 글
본 연구는 농촌진흥청 공동연구사업(과제번호: PJ008461012013) 의 지원에 의하여 연구되었습니다.문
헌
1. Kang KM, Lee SH. Physiological characteristics of starter iso-lated from Kimchi and fermentation of tofu with isoiso-lated starter. Korean J. Soc. Food Sci. Nutr. 41: 1626-1631 (2012)
2. Paik HD, Jung MY, Jung HY, Kim WS, Kim KT. Characteriza-tion of Bacillus polyfermenticus SCD for oral bacteriotherapy gastrointestinal disorders. Korean J. Food Sci. Technol. 34: 73-78 (2002)
3. Jang SY, Sin KA, Park NY, Bang KW, Jeong YJ. Protein changes in soymilk and whole soymilk due to enzymatic hydrolysis. Korean J. Food Preserv. 15: 903-908 (2008)
4. Yang M, Kwak JS, Jang SR, Jia YN, Park IS. Antioxidant activ-ity of soybean yogurt added to tomato extract by Bacillus subtilis and Lactobacillus plantarum. Korean J. Food Nutr. 26: 280-286 (2013)
5. Lee JH, Choung MG. Determination of optimal acid hydrolysis time of soybean isoflavones using drying oven microwave assisted methods. Food Chem. 129: 577-582 (2011)
6. Choi YB, Woo JG, Noh WS. Hydrolysis of β-glycosidic bonds of isoflavone conjugates in the lactic acid fermentation of soy milk. Korean J. Food Sci. Technol. 31: 189-195 (1999)
7. Bae HC, Nam MS. Fermentation properties of the mixed yogurt prepared with bovine milk and soybean milk. Korean J. Food Sci. Ani. Resour. 25: 483-493 (2005)
8. Choi SY, Yoon S. pH, titratable acidity, glucose content, viable cell counting and sensory evaluation of Bifidobacterium longum ATCC 15707 containing milk and soymilk during cold storage. Korean J. Food Sci. Technol. 29: 115-119 (1997)
9. Ko JL, Oh CK, Oh MC, Kim SH. Isolation and identification of lactic acid bacteria from commercial kimchi. J. Korean Soc. Food Sci. Nutr. 38: 732-741 (2009)
10. Wang CY, Wu SC, Ng CC, Shyu YT. Effect of Lactobacillus-fer-mented adlay-based milk on lipid metabolism of hamsters fed cholesterol-enriched diet. Food Res. Int. 43: 819-824 (2010) 11. Ha CG, Cho JK, Chai YG, Heo KC. Isolation and identification
of lactic bacteria containing superior activity of the bile salts deconjugation. Korean J. Sci. Ani. Resour. 24: 164-170 (2004) 12. Cho KM, Ahn BY, Seo WT. Lactic acid fermentation of gamju
manufactured using medicinal herb decoction. Korean J. Food Sci. Technol. 40: 649-655 (2008)
13. Hwang CE, Seo WY, Cho KM. Enhanced antioxidant effect of black soybean by cheonggukjang with potential probiotic Bacillus subtilis CSY191. Korean J. Microbiol. 49: 391-397 (2013) 14. Folin O, Denis W. On phosphotungstic-phosphomolybdic
com-pounds as color reagents. J. Biol. Chem. 12: 239-243 (1912) 15. Cho KM, Lee JH, Yun HD, Ahn BY, Kim H, Seo WT. Changes
of phytochemical constituents (isoflavone, flavanols, and phenolic acids) during cheonggukjang soybeans fermentation using poten-tial probiotics Bacillus subtilis CS90. J. Food Comp. Anal. 24:
402-410 (2011)
16. Gu RX, Yang ZQ, Li ZH, Chen SL, Luo ZL. Probiotic properties of lactic acid bacteria isolated from stool samples of longevous people in regions of Hotan, Xinjiang and Bama, Guangxi, China. Anaeobe 14: 313-317 (2008)
17. Nielsen DS, Møller PL, Rosenfeldt V, Paerregarrd A, Michaelsen KF, Jakobsen M. Case study of the distribution of Mucosa asso-ciated Bifidobacterium species, Lactobacillus species, and other lactic acid bacteria in the human colon. Appl. Environ. Microbiol. 69: 7545-7548 (2003)
18. Eckburg PB, Bik EM, Bernstein CN, Purdom EB, Dethlefsen L, Sargent MC, Gill SR, Nelson KE, Relman DA. Diversity of the human intestinal microbial flora. Science 308: 1635-1638 (2005) 19. Liu XM, Liu WY, Zhang QI, Tian FW, Wang G, Zhang H, Chen
W. Screening of lactobacilli with antagonistic activity against enteroinvasive Escherichia coli. Food Control. 30: 563-568 (2013)
20. Nagata YK, Hashiguchi KJ, Kamimura YY, Yoshida MK, Gomyo TH. The garstrointestinal transit tolerance of Lactobacillus plan-tarum starin No. 14 depended on the carbon source. Biosci. Bio-technol. Biochem. 73: 2650-2655 (2009)
21. Kim KO, Kim WH. Changes in properties of kimchi prepared with different kinds and levels of salted and fermented seafoods during fermentation. Korean J. Food Sci. Technol. 26: 324-330 (1994)
22. Granata LA, Morr CV. Improved acid, flavour and volatile com-pound production in a high-protein and fiber soymilk yogurt-like product. J. Food Sci. 61: 331-336 (1996)
23. Chun JY, Kim JS, Kim JH. Enrichment of isoflavone aglycones in soymilk by fermentation with single and mixed cultures of Sterptococcus infantarius 12 and Weissella sp. 4. Food Chem. 109: 278-284 (2008)
24. Pyo YH, Lee TC, Lee YC. Enrichment of bioactive isoflavones in soymilk fermented with β-glucosidase-producing lactic acid bacteria. Food Res. Int. 38: 551-559 (2005)
25. Angeles AG, Marth EH. Growth and activity of lactic acid bacte-ria in soymilk. J. Milk. Food Technol. 34: 30-36 (1971)
26. Cho KM, Cho HK, Lee JY, Seo WT, Kim MK. Quality charac-teristics and antioxidant effects during makgeolli fermentation by purple sweet potato-rice nuruk. Korean J. Food Sci. Technol. 44: 728-735 (2012)
27. Mccue PP, Shetty KD. Phenolic antioxidant mobilization during yogurt production from soymilk using Kefir cultures. Process Biochem. 40: 1791-1797 (2005)
28. Wang YC, Yu RC, Chou CC. Antioxidant activities of soymilk fermented with lactic acid bacteria and Bifidobacteria. Food Microbiol. 23: 128-135 (2006)
29. Cho KM, Hong SY, Math RK, Lee JH, Kambiranda DM, Kim JM, Islam SMA, Yun MG, Cho JJ, Lim WJ, Yun HD. Biotrans-formation of phenolics (isoflavones, flavanols and phenolic acids) during the fermentation of cheonggukjang by Bacillus pumilus HY1. Food Chem. 114: 413-419 (2009)
30. Otieno DO, Ashton JF, Shah NP. Evaluation of enzyme potential for biotransformation of isoflavone phytoestrogen in soymilk by Bifidobacterium animals, Lactobacillus acidophilus and Lactoba-cillus casei. Food Res. Int. 39: 394-407 (2006)
31. Donkor ON, Shah NP. Production of β-glucosidase and hydrolysis of isoflavone phytoestrogenes by Lactobacillus acidophilus, bifi-dobacterium lactis, and Lactobacillus casei in soymilk. J. Food Sci. 73: 15-20 (2008)
32. Otieno DO, Ashton JF, Shah NP. Role of microbial strain and storage temperatures in the degradation of isoflavone phytoestro-gen in fermented soymilk with selected β-glucosidase producing Lactobacillus casei strains. Food Res. Int. 40: 371-380 (2007) 33. Ewe JA, Wan-Abdullah WN, Alias AK, Liong MT. Effects of
ultrasound on growth, bioconversion of isoflavone and probiotic properties of parent and subsequent passages of Lactobacillus fer-mentum BT 8633 in biotin-supplemented soymilk. Ultrason Sonochem. 19: 890-900 (2012)
34. Marazza JA, Garro MS, de Giori GS. Aglycone production by Lactobacillus rhamnosus CRL981 during soymilk fermentation. Food Microbiol. 26: 333-339 (2009)
35. Wei QK, Chen TR, Chen JT. Using of Lactobacillus bifidobacte-rium to product the isoflavone algycones in fermented soymilk. Int. J. Food Microbiol. 117: 120-124 (2007)
36. Tyug TS, Prasad ND, Ismail AM. Antioxidant capacity, phenolics and isoflavones in soybean by products. Food Chem. 123: 583-589 (2010)
37. Erdil DN, Serventi L, Boyacioglu D, Vodovotz Y. Effect of soymilk powder addition on staling of soy bread. Food Chem. 131: 1132-1139 (2012)
38. Gujral HS, Khanna G. Effect of skim milk powder, soy protein concentrate and sucrose on the dehydration behaviour, texture, color and acceptability of mango leather. J. Food Eng. 55: 343-348 (2002)
39. Wang W, Zhou WB. Characterization of spray dried soy sauce powders using maltodextrins as carrier. J. Food Eng. 109: 399-405 (2012)
40. Wang JC, Wu RN, Zhang WY, Sun ZH, Zhao WJ, Zhang HP. Proteomic comparison of the probiotic bacterium Lactobacillus
casei Zhang cultivated in milk and soy milk. J. Dairy. Sci. 96: 5603-5624 (2013)
41. Choi JS, Kim HY, Seo WT, Lee JH, Cho KM. Roasting enhances antioxidant effect of bitter melon (Momordica charantia L.) increasing in flavan-3-ol and phenolic acid contents. Food Sci. Biotechnol. 21: 19-26 (2012)
42. Lee JW, Lee BW, Kim BL, Kim HT, Ko JM, Baek IY, Seo WT, Kang, YM, Cho KM. Changs in phenolic compounds(isoflavones and phenolic acids) and antioxidant properties in high-protein soybean (Glycine max L., cv. Saedanbaek) for different roasting conditions. J. Korean Soc. Appl. Biol. Chem. 56: 605-612 (2013) 43. Marazza JA, Nazareno MA, de Giori GS, Garro MS.
Enhance-ment of the antioxidant capacity of soymilk by ferEnhance-mentation with Lactobacillus rhamnosus. J. Funct. Food. 4: 594-601 (2012) 44. Heim KE, Tagliaferro AR, Bobilya DJ. Flavonoid antioxidants:
chemistry, metabolism and structure-activity relationships. J. Nutr. Biochem. 13: 572-584 (2002)