학 술 논 문
109
3T3-L1 지방전구세포에서 멜라토닌과 유체전단응력의 영향
이정근 · 이영훈 · 박채림 · 김지현
연세대학교 보건과학대학 의공학부
Effects of Melatonin and Fluid Shear Stress on 3T3-L1 Preadipocytes
Jeongkun Lee, Yeong Hun Lee, Chae Lim Park and Chi Hyun Kim
Department of Biomedical Engineering, Yonsei University, Korea
(Manuscript received 29 March 2018 ; revised 24 April 2018 ; accepted 18 May 2018)
Abstract: Obesity is a worldwide disease caused by the excessive proliferation of adipocytes. Multiple factors, includ- ing melatonin and physical loading, are involved in the control of obesity. Melatonin has been shown to induce apop- tosis on preadipocytes while physical loading such as fluid shear stress (FSS) affects the proliferation and differentiation of adipocytes. Here, we studied the combined effects of melatonin and FSS on 3T3-L1 preadipocytes.
For physical loading, preadipocytes were stimulated with a maximum dynamic fluid shear stress of 1 Pa at 1 Hz for 2 hours with/without melatonin. The experiment conditions were divided into four groups: (1) control, (2) 1 mM mel- atonin treatment, (3) FSS, and (4) combined 1 mM melatonin and FSS. All groups had a fixed duration time of 2 hours.
ERK, p-ERK, COX-2, C/EBPβ, PPARγ, osteopontin, Bax, caspase-3 and caspase-8 proteins were assessed by West- ern blot analysis. GAPDH was used as a control. Results showed that combined melatonin and FSS treatment acti- vated the ERK/MAPK pathway but not COX-2. Furthermore, combined melatonin and FSS treatment significantly decreased C/EBP β and PPARγ compared to other groups. However, caspase-3 and caspase-8 did not result in sig- nificant changes. In summary, combined melatonin and FSS appears to have the potential to inhibit adipogenesis and treat obesity.
Key words: Obesity, Melatonin, Fluid Shear Stress(FSS), Mechanobiology, Adipogenesis
I. 서 론
비만은 과도한 에너지 섭취로 인한 지방 조직의 비정상적 인 발달을 특징으로 하는 대사 질환이다. 이는 제 2형 당뇨, 심혈관계 질환 등 심각한 합병증을 유발하며[1,2] body mass index (BMI) 를 기준으로 30 이상일 시 비만으로 정 의한다[3]. 이 뿐만 아니라 선진국형의 정적인 생활습관과
육류 위주의 식습관으로 인해 비만 인구는 꾸준히 증가하는 추세에 있다[4]. 비만으로 인한 대사 불균형의 직접적인 원 인으로는 지방세포의 과잉 증식 및 성장을 꼽을 수 있는데 [5], 특히 지방세포의 직경이 커질 경우 인슐린 민감도가 저 하되어 혈당 조절에 문제가 생겨 항상성 유지에 있어 지방 의 역할을 잃어버리게 된다[6]. 따라서 비만을 치료하고 증 상을 완화하기 위해 과도한 지방생성(adipogenesis)을 억제 하고 지방세포의 세포자살(apoptosis)을 유발할 수 있는 방 법을 찾기 위한 연구가 이루어지고 있다[7].
지방생성은 지방세포로 분화하기 전 단계인 지방전구세포 (preadipocyte) 가 성숙한 지방세포로 분화하는 일련의 과정 으로, 세포 내 다양한 성장 인자들과 사이토카인(cytokine), 호르몬의 영향을 받는다[8,9]. 지방생성을 조절하기 위한 실 험에서 주로 mouse embryo 유래의 3T3-L1 지방전구세포 Corresponding Author : Chi Hyun Kim
Department of Biomedical Engineering, College of Health Sciences, Yonsei University, Wonju, Gangwon-do, 26493, Korea
TEL: +8233-760-2785 E-mail: [email protected]
이 연구는 교육부의 재원으로 한국연구재단의 지원을 받아 수행된
기초연구사업임 (NRF-2015R1D1A1A01060699).
110
가 사용되며[10], 지방전구세포의 세포신호전달에서 열량 보 존이 주 역할인 백색 지방의 생성을 조절하는 것으로 알려 진 주요 전사 인자들로 peroxisome proliferator-activated receptor gamma (PPAR γ)와 CCAAT/enhancer-binding proteins (C/EBPs) 가 있다[11]. 실험을 통해 위 인자들을 정 량 분석하여 3T3-L1 지방전구세포에서 지방생성이 촉진되 었는지 억제되었는지 알 수 있다.
Apoptosis 는 세포 괴사(necrosis)와는 구분되는 pro- grammed cell death 로서, 세포 위축, 세포막 blebbing, 염 색질 응축 및 DNA 분절 등의 특징들을 나타내며 원하지 않는 세포 및 조직을 비교적 안전하게 제거하는 방법으로 연구되고 있다[12]. 세포 내에서 apoptosis를 유발하는 경 로는 세포막에 존재하는 수용체를 통한 경로와 미토콘드리 아에 의해 유발되는 경로가 있다[13]. 두 경로 모두 단백질 분해 효소의 일종인 caspase family가 관여하고 특히 caspase-3 는 apoptosis의 기질 내 최종 인자로 작용한다 [14].
지방 조직의 과도한 증식을 억제하기 위한 연구는 크게 화학 물질이나 호르몬을 활용한 분자생물학적 접근과[15, 16] 지방세포에 다양한 물리적 자극을 직접적으로 가하는 기계생물학(mechanobiology)적 접근으로 나눌 수 있다 [17]. 특히 호르몬의 경우 생체 주기와 관련한 연구가 지속 되고 있으며, 그 중 송과선(pineal gland)에서 분비되어 생 물의 일주기를 조절하는 멜라토닌(melatonin)과 지방세포 와의 연관성에 대한 연구가 있다. 특히 멜라토닌은 처리 시 간, 농도, 대상 세포의 종류에 따라 세포의 apoptosis와 증 식을 억제하는지 촉진시키는지 달라지기 때문에[18-21] 보 다 심도 있는 연구가 필요하다. 지방생성의 측면에서도 멜 라토닌이 3T3-L1 지방전구세포에서 투여 농도 및 시간에 따라 세포의 증식 및 지방생성을 촉진하며, 지방세포의 생 존에는 크게 기여하지 않는다는 연구결과가 있는 반면[22, 23], fibroblast와 마찬가지로 지방세포로 분화가 가능한 간 엽줄기세포(mesenchymal stem cell)에 투여했을 시 지방 생성을 억제한다는 연구결과 또한 존재한다[24]. 따라서 멜 라토닌과 지방 조직 간의 뚜렷한 상관관계를 규명하기 위하 여 보다 다양한 연구가 이루어져야 한다.
기계생물학적 측면에서 지방 조직은 항상성 유지의 역할 뿐만 아니라 물리적 하중을 견뎌내는 구조물로서도 역할을 수행하며, 각각의 세포는 다양한 물리적 자극에 적응하기 위 해 이에 반응하는 수용체들을 가지고 있다. 이에 비만 치료 의 일환으로 지방 조직과 물리적 자극 간의 상관관계를 규 명하기 위한 시도도 계속되고 있다. 지방세포에 가해지는 자 극은 힘의 크기가 고정된 정적인(static) 자극과 일정한 주 기로 순환하는 동적인(dynamic) 자극으로 나눌 수 있다[17, 25]. 같은 종류의 자극이라 할 지라도 힘의 크기 변화가 정적
인지 동적인지에 따라 세포에 미치는 영향이 달라지는데, 3T3- L1 에 하중을 가하는 실험에서 정적 신축(static stretching) 자 극의 경우 지방생성이 촉진되었지만[26] 정적 압축(static compression) 자극이나 일반적인 동적 자극에는 힘의 방향 에 상관 없이 지방생성이 억제되었다[27,28]. 자극의 종류에 따라 경유하는 신호전달체계(signaling pathway) 또한 차 이가 있는데[17], 지방생성을 억제하는 동적 신축(dynamic stretching) 자극은 공통적으로 cyclooxigenase-2 (COX-2) 와 extracellular signal-regulated kinase 1/2 (EKR 1/2) 를 활성화시킨다는 연구 결과가 있다[27,28].
세포에 동적인 물리적 자극을 가하는 방법 중의 하나로, 세포를 슬라이드 위에 배양한 뒤 그 위로 얇은 공간을 조성 하고 유체가 흐르게 하여 전단응력을 발생시키는 유체전단 응력(fluid shear stress, FSS)이 있다[29]. 주로 내피세포 (endothelial cell) 나 조골세포(osteoblast), 간엽줄기세포와 같은 부착세포를 대상으로 실행하며[30], 간엽줄기세포가 뼈 세포로 분화하도록 유도하는 한편 조골세포의 뼈 생성을 유 도한다[31]. 하지만 지금까지 지방세포와 유체전단응력 간 의 상관관계에 대한 연구는 활발히 이루어지지 않았으며, 물 리적 자극을 통한 연구에서 이를 유추하는 정도로 진행되었다.
우리는 지방생성의 억제 및 지방세포의 apoptosis를 유 발하는 기전을 다방면으로 분석하기 위하여, 3T3-L1 지방 전구세포에 멜라토닌 처리와 유체전단응력을 동시에 가하여 지방생성인자의 변화 및 apoptosis 인자의 변화를 살펴보 았다. 또한 각각의 개별적인 영향과도 비교하기 위하여 실 험 군을 네 개로 나누어 각각 대조군, 멜라토닌, 유체전단응 력, 멜라토닌과 유체전단응력을 동시에 가한 조건으로 실험 을 진행하였다.
II. 실험 방법
1. 세포 배양
세포는 지방 조직 및 세포 실험에 주로 사용되는 mouse- derived 3T3-L1 지방전구세포를 사용, 1주일에 한 번 confluence 가 80%이상 도달했을 때 계대 배양을 수행하였다.
사용한 배지는 Dulbecco’s modified eagle’s medium (DMEM) 에 calf serum (CS) 5%, penicillin/streptomycin 1% 를 혼합하여 사용하였다. 배지 교환은 새로 계대 배양한 지 3일 후에 실시하였다. 세포 배양에 사용된 모든 용액은 사 용 전 water bath를 이용하여 37
oC 로 맞춰 사용하였다. 세 포 배양기는 온도 37
oC 에 이산화탄소 농도 5%로 고정하였다.
2. 유체전단응력 실험
유체전단응력 실험에 쓰이는 도구는 멸균 후 70% 에탄올
로 소독하여 사용하였다. 그 후 충분히 배양된 3T3-L1을
111 trypsin 을 이용하여 떼어준 뒤 슬라이드에 6 × 10
5개씩 지
방전구세포를 배양하였다. 이를 자체 제작한 유체전단응력 기기와 chamber 를 사용하여, 진동수는 1 Hz에 1 Pa의 압 력으로 총 2시간 동안 가해주었다[29,32,33]. 전단응력에 사 용된 배지는 세포배양에 쓰인 것과 동일한 조건으로 맞추었 으며, 멜라토닌과 유체전단응력을 동시에 처리할 실험군은 멜라토닌을 투여한 배지를 사용하였다. 유체전단응력을 하 지 않는 실험군은 2시간 동안 실온에 두었다. 멜라토닌의 농 도는 1 mM로 고정해 주었다[23].
3. Western Blot
Gel 제작 시 acrylamide/bis 농도는 running gel 9%, stacking gel 5% 로 설정하였으며, 80 V의 전압으로 2시간 동안 전기영동을 실시하였다. 전기영동 후, 100 V의 전압에 서 50분 동안 단백질을 PVDF membrane으로 transfer한 후 shaker를 이용, membrane을 1시간 동안 5% skim milk로 blocking 하였다. 1차 항체는 BSA에 1:1000의 비 율로 희석하여 사용하였고, shaker에서 배양하였다. 2차 항 체는 5% skim milk에 1:5000의 비율로 희석하여 사용하 였고, 1시간 동안 shaker에서 배양하였다. 항체 처리의 각 과정 사이에는 TBS-T용액을 사용하여 10분씩 네 번, 총 40 분 간 membrane을 shaker로 washing하였다.
4. 통계 분석
감광하여 필름에 검출된 밴드는 Image J 프로그램을 이 용하여 밴드의 상대적 밝기를 측정 후, Graphpad Prism 5 프로그램을 이용하여 그래프를 산출하였고, 같은 프로그 램을 이용하여 one-way ANOVA를 수행하였다. 결과 값은 평균 ± 표준편차 (SD)로 표현하였고, 사후 검정은 Tukey’s test 를 사용하였다. 유의 수준은 0.05로 설정하였다.
III. 결 과
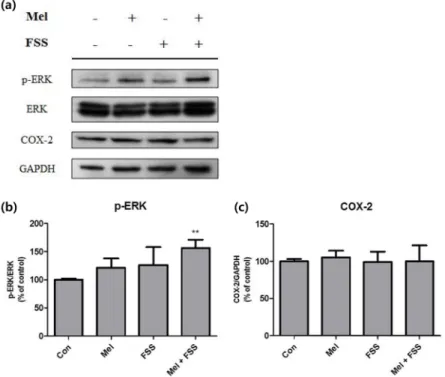
먼저 각각 멜라토닌과 유체전단응력, 혹은 두 자극을 동 시에 가해 주었을 때 세포의 신호전달체계 관련 인자들을 비교하였다(그림 1a). COX-2의 경우 어떤 자극에도 유의한 변화를 보이지 않았다(그림 1b). ERK는 단일 자극만 가하 였을 때 멜라토닌과 유체전단응력 실험군 사이에 큰 차이를 보이지 않았으나, 두 자극을 함께 주었을 때 대조군은 물론 단일 자극 실험군에 비해서도 높은 발현을 보였다(그림 1c).
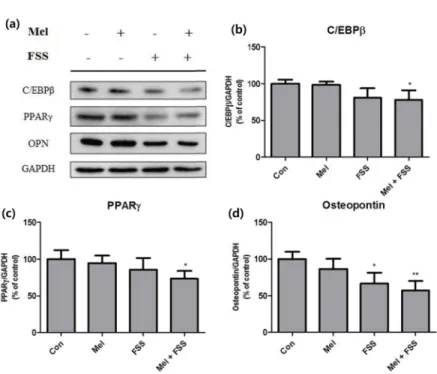
지방생성 인자는 전체적으로 자극에 의해 대조군 대비 활 성이 감소하였다. 지방생성 시 C/EBPα의 선행 단계에서 관 여하는 C/EBPβ의 경우 유체전단응력 실험군과 두 자극을 동시에 처리한 실험군에서 유의하게 감소하였다(그림 2b).
또한 지방생성의 핵심 지표인 PPARγ도 C/EBPβ와 유사한 변화 양상을 보였다(그림 2c). 마지막으로 지방 조직에서
그림 1. 3T3-L1에 웨스턴 블롯을 실시하여 대조군 대비 세포의 신호전달체계 활성화 정도를 분석한 것. (a) 웨스턴 블롯 결과, (b) p-ERK, (c) COX-2. (n = 4, **P < 0.01).
Fig. 1. Signaling pathway activity of treated 3T3-L1 preadipocytes measured with western blot analysis. (a) Bands of western blot analysis, (b) p-ERK, (c) COX-2. (n = 4, **P < 0.01).
112 cleaved form 으로 분비되며 비만 환자에게서 염증을 유발 하는 osteopontin (OPN)은 멜라토닌만을 투여했을 때 cleaved form 의 분비가 소폭 감소하였으며 유체전단응력을 단독으로 가하였을 때와 멜라토닌과 함께 처리하였을 때 큰 폭으로 감소하였다(그림 2d).
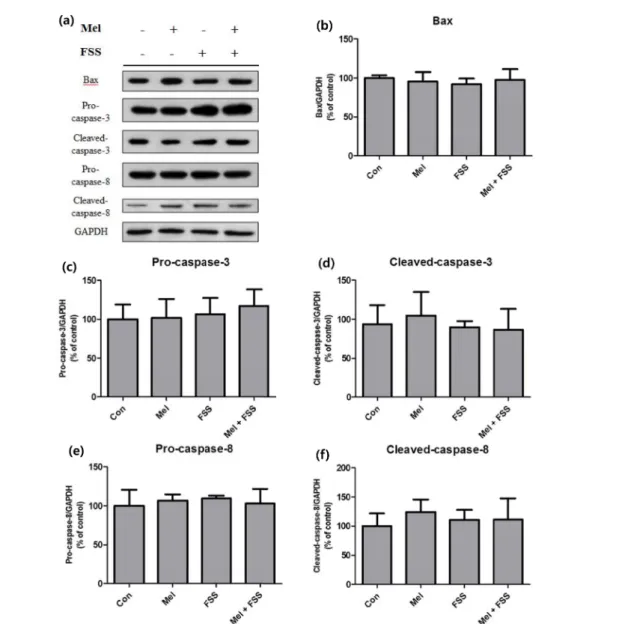
Apoptosis 관련 인자의 경우 전체적으로 유의한 차이가 나타나지 않았다. 미토콘드리아를 통하여 apoptosis를 조절 하는 Bax는 모든 실험군에서 거의 변화하지 않았다(그림 3b). Cell death 수용체에 의해 활성화되는 caspase-3와 caspase-8 은 멜라토닌만 단독으로 투여해 주었을 때 cleaved form 의 활성 변화가 통계적으로 유의하지 않았고, FSS를 처 리해 주었을 때는 멜라토닌의 유무와 상관없이 대조군과 비 슷한 수준의 활성을 보여주었다(그림 3d, 3f).
IV. 고찰 및 결론
본 실험은 지방 조직의 증식을 억제하고 지방세포의 지방 형성을 억제하는 방법을 찾기 위하여 기존의 생화학적 접근 법과 물리적 접근법을 융합하였다. 지방생성과 apoptosis에 관련이 있다고 알려진 멜라토닌과 유체전단응력을 3T3-L1 에 동시에 가하여, 두 자극을 개별적으로 가하였을 때와 비 교하여 차이가 있는지, 지방 감소에 효과가 있는지 분석하 였다. 본 실험에서 사용된 멜라토닌의 농도는, 지방세포에
멜라토닌을 투여한 기존의 여러 연구들과[22-24], 멜라토닌 과 유체전단응력을 동시에 처리한 이전 실험방법을 근거로 설정하였다[33].
먼저 신호전달체계의 활성화 정도를 비교하였다. 실험 결 과 ERK의 활성화 형태인 p-ERK는 멜라토닌과 유체전단 응력 두 자극에 모두 반응하여 활성화되었으며 특히 두 자 극을 동시에 가해 주었을 때 큰 폭으로 증가하였다. 이를 통 해 두 자극을 동시에 가해 주었을 때 외부자극에 의한 신호 전달이 더 활발히 이루어졌음을 알 수 있다. 선행연구에서 COX-2 는 지방세포가 정적 압축(static compression) 자극 이나 동적 신축(dynamic stretching) 자극을 받을 때 활성 화되어 지방생성을 억제하는 역할을 하였다[27,28]. 본 실 험결과에서 COX-2는 모든 실험군에서 유의한 차이를 보이 지 않았다. 유체전단응력만으로는 지방전구세포의 COX-2발 현을 활성화하기에 부족하거나, 유체전단응력의 세포 자극 이 기존연구의 신축(stretching) 자극과는 다른 유형의 물 리적 자극인 것으로 사료된다.
지방생성 인자의 경우 유체전단응력에 의하여 C/EBPβ와 PPAR γ의 발현이 감소하였다. C/EBPβ의 경우 유체전단응 력만 단독으로 처리했을 때와 멜라토닌을 함께 처리했을 때 큰 차이가 없었으나 PPARγ는 두 자극을 함께 가하였을 때 더 큰 영향을 받는 것을 확인할 수 있었다. 이를 통하여 지 방생성 억제 효과는 두 자극을 단독으로 가할 때보다 함께
그림 2. 3T3-L1에 웨스턴 블롯을 실시하여 대조군 대비 세포의 지방생성 인자의 활성화 정도를 분석한 것. (a) 웨스턴 블롯 결과, (b) C/EBPβ, (c) PPARγ, (d) OPN. (n = 4, *P < 0.05; ** P < 0.01).
Fig. 2. Adipogenesis factors of treated 3T3-L1 preadipocytes measured with western blot analysis. (a) Bands of western blot analysis, (b) C/EBPβ, (c) PPARγ, (d) OPN. (n = 4, *P < 0.05; **P < 0.01).
113
가하였을 때 보다 효과적이며, 유체전단응력이 멜라토닌보 다 지방생성 억제에 주는 영향이 보다 크다는 것을 확인할 수 있었다. 또한 OPN cleavage의 변화도 분석하였는데, C/
EBPβ의 변화 양상과 유사하게 대조군, 멜라토닌, 유체전단 응력, 동시 자극 순서대로 cleaved form의 발현이 줄어들 었다. OPN은 지방 조직에서 cleaved form으로 분비되어 비만환자에게서 염증을 유발하고, 또한 체지방률이 높을수 록 그 분비량이 증가한다[34]. OPN이 직접적으로 지방생 성에 기여하는지는 명확히 밝혀지지 않았지만, 간접적으로 멜라토닌과 유체전단응력에 의한 분비량 변화로 인하여 비 만 증상의 개선에 효과가 있을 것으로 사료된다.
마지막으로 apoptosis를 유도하는 인자들을 분석하였다.
모든 실험군에서 Bax는 유의미한 변화를 보이지 않았으므로 멜라토닌과 유체전단응력은 미토콘드리아를 통한 apoptosis 에 영향을 주지 않았음을 확인하였다. 반면에 caspase-3와 caspase-8의 경우 통계적 유의성은 검정되지 않았지만 멜 라토닌을 투여해 주었을 때 대조군에 비해 발현이 약간 증 가하는 양상을 보였다. 멜라토닌은 암세포에서 apoptosis조 절을 통하여 종양 증식을 억제한다는 연구결과가 보고되었 다[19]. 또한 본 실험결과에서 멜라토닌과 유체전단응력을 함께 가해 주었을 때, 유체전단응력이 멜라토닌의 영향을 상 쇄하거나 apoptosis를 억제하였을 수 있다는 것을 caspase 의 발현 감소로 유추해 볼 수 있었다. Osteoblast에 종양 괴사 인자(tumor necrosis factor)를 활용한 이전 연구에서
그림 3. 3T3-L1에 웨스턴 블롯을 실시하여 대조군 대비 세포의 apoptosis 관련 인자의 활성화 정도를 분석한 것. (a) 웨스턴 블롯 결과, (b) Bax, (c) Pro-caspase-3, (d) Cleaved-caspase-3, (e) Pro-caspase-8, (f) Cleaved-caspase-8. (n = 4).Fig. 3. Apoptotic factors of treated 3T3-L1 preadipocytes measured with western blot analysis. (a) Bands of western blot analysis, (b) Bax, (c) Pro-caspase-3, (d) Cleaved-caspase-3, (e) Pro-caspase-8, (f) Cleaved-caspase-8. (n = 4).
114
는 약물과 유체전단응력을 세포에 동시에 처리해 주었을 때 종양 괴사 인자에 의한 세포의 apoptosis가 억제되었고, 동 일하게 caspase-3의 발현이 억제되는 결과가 나타났다[35].
이를 통해 유체전단응력은 cell death 수용체를 통한 apoptosis 에 부적합하며 오히려 멜라토닌을 단독으로 투여 하는 것이 더 효과적이라고 사료된다.
본 연구를 통해 멜라토닌과 유체전단응력이 3T3-L1의 지 방생성과 apoptosis에 주는 영향을 실험하였다. 지방생성 억 제의 경우 멜라토닌과 유체전단응력을 동시에 처리해 주었 을 때 각각의 효과가 증진되어 보다 유의한 효과를 보였으 나, 반대로 apoptosis는 두 자극이 상충하였다. 본 실험 결 과와 유사하게, 멜라토닌과 유체전단응력을 osteoblast에 동 시에 가해준 이전 실험에서 osteoblast의 동화작용과 관련 된 인자들의 변화가, 자극의 조합에 따라 상쇄되거나 증가 한 것을 알 수 있었다[33]. 이는 세포에 대한 생화학적 자 극과 물리적 자극이 동시에 가해질 때, 각 자극의 영향과 특 성에 따라 효과가 달라질 수 있음을 시사한다. 본 실험에서 지방생성 억제의 경우 단독으로 처리한 결과를 비교했을 때 유체전단응력이 더 효과적이었던 반면 지방세포의 apoptosis 의 경우 멜라토닌이 효과적일 수 있는 양상을 보였다.
결론적으로, 이 실험을 통하여 지방세포에 멜라토닌 또는 유체전단응력을 단독으로 가할 때보다 함께 가하였을 때 지 방 증식 억제 효과가 보다 강하다는 것을 알 수 있었다. 이 러한 결과는 비만 관리를 위한 운동 시 혈중 멜라토닌 농도 가 높은 밤이 낮보다 더 효과적임을 제안할 수 있다.
참고문헌
[1] P.G. Kopelman, “Obesity as a medical problem,” Nature, vol.
404, pp. 635-643, 2000.
[2] X. Formiguera, and A. Cantón, “Obesity: epidemiology and clinical aspects,” Best Pract. Res. Clin. Gastroenterol., vol.
18, no. 6, pp. 1125-1146, 2004.
[3] K.M. Flegal, M.D. Carroll, and B.K. Kit, “Prevalence of obe- sity and trends in the distribution of body mass index among US adults, 1999-2010,” Jama., vol. 307, no. 5, pp. 491-497, 2012.
[4] N.S. Wellman, and B. Friedberg, “Causes and consequences of adult obesity: health, social and economic impacts in the United States,” Asia Pac. J. Clin. Nutr., vol. 11, no. 8, pp.
705-709, 2002.
[5] T. Furuyashiki, H. Nagayasu, Y. Aoki, H. Bessho, T.
Hashimoto, K. Kanazawa, and H. Ashida, “Tea catechin sup- presses adipocyte differentiation accompanied by down-reg- ulation of PPARγ2 and C/EBPα in 3T3-L1 cells,” Biosci.
Biotechnol. Biochem., vol. 68, no. 11, pp. 2353-2359, 2004.
[6] C. Weyer, J.E. Foley, C. Bogardus, P.A. Tataranni, and R.E.
Pratley, “Enlarged subcutaneous abdominal adipocyte size, but not obesity itself, predicts type II diabetes independent of insulin resistance,” Diabetologia, vol. 43, no. 12, pp. 1498- 1506, 2000.
[7] Y.W. Wang, and P.J.H. Jones, “Conjugated linoleic acid and obesity control: efficacy and mechanisms,” Int. J. Obes., vol.
28, no. 8, pp. 941-955, 2004.
[8] D.B. Hausman, M. DiGirolamo, T.J. Bartness, G.J. Hausman, and R.J. Martin, “The biology of white adipocyte prolifera- tion,” Obes. Rev., vol. 2, no. 4, pp. 239-254, 2001.
[9] O.A. MacDougald, and S. Mandrup, “Adipogenesis: forces that tip the scales,” Trends Endocrinol. Metab., vol. 13, no. 1, pp. 5-11, 2002.
[10] H. Green, and O. Kehinde, “Sublines of mouse 3T3 cells that accumulate lipid,” Cell, vol 1, no. 3, pp. 113-116, 1974.
[11] M.I. Lefterova, and M.A. Lazar, “New developments in adi- pogenesis,” Trends Endocrinol. Metab., vol. 20, no. 3, pp.
107-114, 2009.
[12] K. Vermeulen, D.R.V Bockstaele, and Z.N. Berneman,
“Apoptosis: mechanisms and relevance in cancer,” Ann.
Hematol., vol. 84, no.10, pp. 627-639, 2005.
[13] K.J. Woo, Y.J. Jeong, J.W. Park, and T.K. Kwon, “Chrysin- induced apoptosis is mediated through caspase activation and Akt inactivation in U937 leukemia cells,” Biochem. Biophys.
Res. Commun., vol. 325, no. 4, pp. 1215-1222, 2004.
[14] M.O. Hengartner, “The biochemistry of apoptosis,” Nature, vol. 407, pp. 770-776, 2000.
[15] C.L. Hsu, and G.C. Yen, “Effects of capsaicin on induction of apoptosis and inhibition of adipogenesis in 3T3-L1 cells,” J.
Agric. Food Chem., vol. 55, no. 5, pp. 1730-1736, 2007.
[16] S. Rayalam, J.Y. Yang, S. Ambati, M.A. Della-Fera, and C. A.
Baile, “Resveratrol induces apoptosis and inhibits adipogen- esis in 3T3?L1 adipocytes,” Phytother. Res., vol. 22, no. 10, pp. 1367-1371, 2008.
[17] N. Shoham, and A. Gefen. “Mechanotransduction in adipo- cytes,” J. Biomech., vol. 45, no. 1, pp. 1-8, 2012.
[18] R.M. Sainz, J.C. Mayo, C. Rodriguez, D.X. Tan, S. Lopez- Burillo, and R.J. Reiter, “Melatonin and cell death: differen- tial actions on apoptosis in normal and cancer cells,” Cell Mol. Life Sci., vol. 60, no. 7, pp. 1407-1426, 2003.
[19] D.E. Blask, L.A. Sauer, R.T. Dauchy, E.W. Holowachuk, M.S.
Ruhoff, and H.S. Kopff, “Melatonin inhibition of cancer growth in vivo involves suppression of tumor fatty acid metabolism via melatonin receptor-mediated signal transduc- tion events,” Cancer res., vol. 59, no. 18, pp. 4693-4701, 1999.
[20] J.A. Roth, T. Rosenblatt, A. Lis, and R. Bucelli, “Melatonin- induced suppression of PC12 cell growth is mediated by its Gi coupled transmembrane receptors,” Brain res., vol. 919, no. 1, pp. 139-146, 2001.
[21] D.L. Drazen, D. Bilu, S.D. Bilbo, and R.J. Nelson, “Mela- tonin enhancement of splenocyte proliferation is attenuated by luzindole, a melatonin receptor antagonist,” Am. J.
Physiol. Regul. Integr. Comp. Physiol., vol. 280, no. 5, pp.
R1476-R1482, 2001.
[22] K. Zwirska-Korczala, J. Jochem, M. Adamczyk-Sowa, P.
Sowa, R. Polaniak, E. Birkner, M. Latocha, K. Pilc, and R.
Suchanek, “Influence of melatonin on cell proliferation, anti- oxidative enzyme activities and lipid peroxidation in 3T3-L1 preadipocytes-an in vitro study,” J. Physiol. Pharmacol., vol.
56, pp. 91-99, 2005.
[23] H. Kato, G. Tanaka, S. Masuda, J. Ogasawara, T. Sakurai, T.
Kizaki, H. Ohno, and T. Izawa, “Melatonin promotes adipo- genesis and mitochondrial biogenesis in 3T3-L1 preadipo-
115
cytes,” J. Pineal. Res., vol. 59, no. 2, pp. 267-275, 2015.[24] L. Zhang, P. Su, C. Xu, C. Chen, A. Liang, K. Du, Y. Peng, and D. Huang, “Melatonin inhibits adipogenesis and enhances osteogenesis of human mesenchymal stem cells by suppressing PPARγ expression and enhancing Runx2 expres- sion,” J. Pineal. Res., vol. 49, no. 4, pp. 364-372, 2010.
[25] E. Linder-Ganz, N. Shabshin, Y. Itzchak, and A. Gefen,
“Assessment of mechanical conditions in sub-dermal tissues during sitting: a combined experimental-MRI and finite ele- ment approach,” J. Biomech., vol. 40, no. 7, pp. 1443-1454, 2007.
[26] Y. Hara, S. Wakino, Y. Tanabe, M. Saito, H. Tokuyama, N.
Washida, S. Tatematsu, K. Yoshioka, K. Homma, K. Hase- gawa, H. Minakuchi, K. Fujimura, K. Hosoya, K. Hayashi, K. Nakayama, and H. Itoh, “Rho and Rho-kinase activity in adipocytes contributes to a vicious cycle in obesity that may involve mechanical stretch,” Sci. Signal., vol. 4, no. 157, pp.
ra3, 2011.
[27] Y. Tanabe, M. Koga, M. Saito, Y. Matsunaga, and K.
Nakayama, “Inhibition of adipocyte differentiation by mechanical stretching through ERK-mediated downregula- tion of PPARγ2,” J. Cell. Sci., vol. 117, no. 16, pp. 3605- 3614, 2004.
[28] Y. Tanabe, Y. Matsunaga, M. Saito, and K. Nakayama,
“Involvement of cyclooxygenase-2 in synergistic effect of cyclic stretching and eicosapentaenoic acid on adipocyte dif- ferentiation,” J. Pharmacol. Sci., vol. 106, no. 3, pp. 478- 484, 2008.
[29] H. Huang, R.D. Kamm, and R.T. Lee. “Cell mechanics and
mechanotransduction:pathways, probes, and physiology,”
Am. J. Physiol. Cell. Physiol., vol. 287, no. 1, pp. C1-C11, 2004.
[30] T.M. Maul, D.W. Chew, A. Nieponice, and D.A. Vorp,
“Mechanical stimuli differentially control stem cell behavior:
morphology, proliferation, and differentiation,” Biomech.
Model. Mechanobiol., vol. 10, no. 6, pp. 939-953, 2011.
[31] L. Liu, W. Yuan, and J. Wang, “Mechanisms for osteogenic differentiation of human mesenchymal stem cells induced by fluid shear stress,” Biomech. Model. Mechanobiol., vol. 9, no. 6, pp. 659-670, 2010.
[32] C.H. Kim, L. You, C.E. Yellowley, and C.R. Jacobs, “Oscil- latory fluid flow-induced shear stress decreases osteoclasto- genesis through RANKL and OPG signaling,” Bone, vol. 39, no. 5, pp. 1043-1047, 2006.
[33] C.H. Kim, and Y.M. Yoo, “Fluid shear stress and melatonin in combination activate anabolic proteins in MC3T3-E1 osteo- blast cells,” J. Pineal Res., vol. 54, no. 4, pp. 453-461, 2013.
[34] M. Zeyda, K. Gollinger, J. Todoric, F.W. Kiefer, M. Keck, O.
Aszmann, G. Prager, G.J. Zlabinger, P. Petzelbauer, and T.M.
stulnig, “Osteopontin is an activator of human adipose tissue macrophages and directly affects adipocyte function,” Endo- crinology, vol. 152, no. 6, pp. 2219-2227, 2011.
[35] F.M. Pavalko, R.L. Gerard, S.M. Ponik, P.J. Gallagher, Y. Jin, and S.M. Norvell, “Fluid shear stress inhibits TNF-α-induced apoptosis in osteoblasts: A role for fluid shear stress-induced activation of PI3-kinase and inhibition of caspase-3,” J. Cell.
Physiol., vol. 194, no. 2, pp. 194-205, 2003.