합성된 쿼럼 신호 유사 물질에 의한 녹농균 쿼럼 센싱 및 생물막 형성의 제어
김수경1·김철진2·윤제용3·이준희1*
1부산대학교 약학대학 미생물학 연구실
2Departments of Chemistry, Princeton University, Princeton, New Jersey 08544, USA
3서울대학교 화학생물공학부
Received: December 22, 2010 / Revised: January 14, 2011 / Accepted: January 24, 2011
Inhibition of Quorum Sensing and Biofilm Formation by Synthetic Quorum Signal Analogues in Pseudomonas aeruginosa. Kim, Soo-Kyoung1, Cheoljin Kim2, Jeyong Yoon3, and Joon-Hee Lee1*.
1Department of Pharmacy, College of Pharmacy, Pusan National University, Busan 609-735, Korea,
2Departments of Chemistry, Princeton University, Princeton, New Jersey 08544, USA, 3School of Chemical and Biological Engineering, College of Engineering, Seoul National University, Seoul 151-742, Korea − Pseudomonas aeruginosa is an opportunistic pathogen that causes various infections on urinary track, cornea, respiratory track, and burn wound site, and mainly relies on quorum sensing (QS) for its virulence. To control the infectivity of P. aeruginosa, we previously synthesized the structural analogues of a major QS signal, N-3-oxododecanoyl homoserine lactone (3OC12-HSL) to use as a QS inhibitor. Two of them (5b and 5f) had been confirmed to have an inhibitory effect on LasR, a major QS signal receptor of P. aeruginosa in the screening by the recombinant Escheri- chia coli reporter. To further evaluate these compounds, we tested their efficacy to control the QS and virulence of P. aeruginosa. Unlike the result from E. coli reporter, both 5b and 5f failed to affect the LasR activity in P. aerugi- nosa, but instead they selectively affected the activity of QscR, another 3OC12-HSL receptor of P. aeruginosa.
Interestingly, their effect on QscR was complex and opposite to what we obtained with E. coli system. Both 5b and 5f enhanced the QscR activity at the low concentration range (< 10µM), but high concentration of 5f (≈1 mM) strongly inhibited QscR. While 5b and 5f didn’t affect the production of proteases, the key virulence factor, they significantly reduced the biofilm formation that is important in mediating chronic infections. Especially, 5f inhibited the initial attachment of P. aeruginosa, rather than the biofilm maturation. Based on our results, we suggest that 5f can be applied for an anti-biofilm agent without increasing virulence of P. aeruginosa.
Key words: Pseudomonas aeruginosa, quorum sensing, Acyl-homoserine lactone, inhibitor, biofilm, QscR
서 론
그람음성 간균인 녹농균(Pseudomonas aeruginosa)은 면 역이 약화된 사람들에게 특히 치명적인 기회감염성 병원균 이다[25]. 각막, 요도, 화상 부위 등의 국소 감염을 일으키며, 낭포성 섬유증(cystic fibrosis) 등의 호흡기 질병의 진행에 깊이 관여한다[25], 여기에 관여하는 병독인자(virulence factors)에는 편모, 섬모, lipopolysaccharide, 독소(exotoxin A, exoenzyme S), 각종 프로테아제[elastase (LasB), LasA protease, alkaline protease, protease IV], 용혈소(hemolysin), 계면활성 성분(rhamnolipid) 등이 알려져 있다[1, 2, 5, 6, 16, 24, 36]. 특히 ventilator나 카테터(catheter)와 같은 장비
를 삽입한 환자들에서 흔히 일어나는 병원 교차감염에 매우 중요한 원인균이며, 여기에는 이러한 장비의 표면에 형성되 는 생물막(biofilm)이 매우 중요한 역할을 하는 것으로 알려 져 있다[13, 35].
이러한 병독인자들 중에는 감염시 주변 환경에 따라 특이 적으로 발현되는 것들이 많기 때문에 여기에 관여하는 주변 환경 인지 및 병독인자 발현 조절 단백질들은 병원균의 병 원성을 제어하기 위한 중요한 타겟으로 여겨져 왔으며, 이 들의 활성을 억제, 교란, 조절 할 수 있는 물질들이 새로운 항생 전략의 일환으로 많이 연구되어 왔다[14]. 이 중 가장 많이 연구된 것이 세포밀도 인식 기전으로 처음 발견되어 이 후 여러 세균에서 병독인자들의 발현, 생물막 형성, 운동성, 항생제 내성 등과 같은 병원성 발현에 매우 중요한 조절 기 전으로 밝혀진 쿼럼 센싱(quorum sensing; QS)이다[30, 37, 38]. 다양한 병원균들에서 QS는 병원성을 제어하기 위한 좋 은 타겟으로 여겨지고 있으며, 다양한 QS 억제 물질들이 광
*Corresponding author
Tel: +82-051-510-2821, Fax: +82-051-513-6754 E-mail: [email protected]
범위하게 탐색되어 왔다[3, 12, 17, 19, 25]. 더욱이 QS은 전체 만성감염의 65% 이상을 매개한다고 추정되는 생물막 에 의한 감염에서도 중요한 역할을 한다고 알려져 있어[8, 39], QS가 세균의 병원성과 감염력을 억제하기 위한 좋은 타겟임을 말해준다.
QS 조절 기전은 일반적으로 확산이 가능한 신호 물질과 이를 합성하는 효소, 그리고 이를 수용하는 수용체 단백질로 구성된다. QS 신호물질은 세균에 따라 그 구조와 종류가 매 우 다양한데, 녹농균의 경우, N-3-oxododecanoyl homoserine lactone(3OC12-HSL)과 N-buturyl homoserine lactone(C4- HSL), 2-heptyl-3-hydroxy-4-quinolone(Pseudomonas quinolone signal; PQS) 등을 주된 QS 신호물질로 생성한다[11, 31].
그런데 C4-HSL과 PQS의 생성은 3OC12-HSL 신호에 의존 적이어서 QS 관련 병독 인자들의 대부분이 3OC12-HSL에 의해 발현이 조절된다[10]. 3OC12-HSL은 LasR과 QscR이 라는 두 개의 수용체 단백질에 의해 수용되는데, 이중 LasR 이 가장 초기에 3OC12-HSL을 수용하여 이후 C4-HSL과 PQS 신호전달 시스템을 활성화 시키기 때문에 병원성 제어 를 위한 가장 근본적인 타겟으로 생각되어 왔으며, 대부분 의 QS 억제 물질들은 LasR의 3OC12-HSL 수용을 경쟁적 으로 방해하는 길항제(antagonist)로써 발견된 것들이다[17, 19, 33]. 지금까지 발견된 QS 억제 물질 중 가장 주목할 만 한 것은 furanone 유도체이나[4, 18, 23], 여전히 병원성 세 균에 특이적으로 작용하는 보다 나은 QS 저해제가 요구되 고 있는 실정이다.
이전 연구에서 본 연구팀은 in silico 모델링을 이용하여 녹농균의 LasR에 3OC12-HSL보다 더 잘 결합할 수 있을 것 으로 예상되는 구조 유사체들을 예측하고 이를 합성하여 그 활성을 대장균 리포터 균주를 이용하여 스크리닝한 바 있다 [17, 20]. 특히 acyl tail 부분에 변화를 준 furanone 화합물
들 중 5b와 5f가 효과적으로 대장균 리포터 균주에서 QS 반 응을 저해한 바 있었다(Fig. 1)[20]. 본 연구에서는 이 물질 들이 실제 녹농균에서도 효과적으로 QS 반응을 억제할 수 있는지, LasR과 QscR중 어느 수용체 단백질을 통해 이러한 QS 저해 효과가 나타나는지 조사하였으며, 이들이 병독소 (virulence factor)의 발현 및 생물막 형성과 같은 병원성 매 개 인자들에 실제 어떻게 영향을 미치는지 분석해 보았다.
재료 및 방법 녹농균 균주 및 배양 조건
본 실험에서는 녹농균 야생형 균주로 PAO1을 사용하였다 [34]. 대부분의 실험에서 녹농균은 Luria-Bertani(LB) 배지에 서 37oC, 호기적 진탕배양을 통해 배양되었다. 실험에 사용 된 플라스미드를 포함하는 녹농균의 선택적 배양을 위해서 는 LB 배지에 carbenicillin이 150 µg/ml의 농도로 첨가되었 다. 프로테아제 활성 측정과 생물막 형성 실험을 위해서는 다른 조성의 배지가 사용되었는데, 이에 대하여는 아래에 따 로 설명하였다.
녹농균에서 QS 신호 수용체 단백질의 활성 측정
녹농균의 주된 QS 신호물질인 3OC12-HSL에 대한 수용 체 단백질인 LasR과 QscR의 활성을 리포터 유전자를 이용 하여 측정하였다. LasR 활성 측정을 위하여 3OC12-HSL에 의해 활성화된 LasR이 특이적으로 결합하여 발현을 증가시 키는 lasI 유전자에 lacZ 유전자를 리포터로 결합시킨 pSC11 플라스미드를 사용하였으며[7], QscR 활성 측정을 위하여는 3OC12-HSL에 의해 활성화된 QscR이 특이적으로 발현을 유 도하는 PA1897 유전자의 프로모터에 lacZ를 결합시킨 pJL101 플라스미드를 사용하였다[21]. 리포터 유전자인 lacZ 로부터 발현되는 β-galactosidase의 양은 각각 LasR과 QscR 의 활성을 의미한다[21]. 이 플라스미드들을 각각 녹농균 야 생형 균주인 PAO1에 형질전환을 통해 도입한 후, 이를 LB (Luria-Bertani) broth에 접종하여 그림에서 표시된 것과 같 이 다양한 농도의 저해제들과 함께 37oC에서 배양하였다. 배 양 시간에 따라 200 µl씩 배양액을 채취하여 아래에 기술한 방법에 따라 세포의 OD(optical densities)와 β-galactosidase 활성을 측정하였다
β-galactosidase 활성 측정
β-galactosidase 활성 측정은 Galacto-Light PlusTM kit (Applied Biosystems, Foster City, CA, USA)를 이용하여 이전에 보고된 바와 같이 수행되었다[21, 26]. 배양 시간에 따라 200 µl씩 덜어낸 녹농균 배양액 중 100 µl로 600 nm 에서 OD를 측정하였다(Tristar LB941, Berthold, 독일). 한 편 나머지 100 µl에는 CHCl3을 10 µl 넣고 voltex를 이용 해 강하게 섞어준 후, 15분간 가만히 놓아두어 첨가해 준 Fig. 1. The molecular structures of 5b and 5f. These com-
pounds were suggested as QS inhibitors in the previous study [20].
CHCl3을 완전히 분리시키고 상층의 10 µl만을 취하여 96- well plate에 옮겼다. 여기에 Galacto-light plusTM 기질 0.6 µl를 diluents buffer 100 µl에 희석한 용액을 각 well에 넣 고 어두운 곳에서 1시간 동안 반응시켰다. 이어 accelerator2 용액 150 µl을 각 well에 넣고 발생되는 luminescence를 측정 하였다(Tristar LB941, Berthold, 독일). 측정된 luminescence 값을 앞에서 측정한 OD600 값으로 표준화(luminescence/
OD600)하여 제시하였다[1, 21].
프로테아제 활성 측정
프로테아제 활성은 skim milk agar(0.5% skim milk, 0.5% peptone, 0.1% glucose, 1.5% agar)를 이용하여 측정 되었다. Skim milk agar plate위에 지름 0.5 cm의 여과지를 놓고 그 위에 그림에 표시된 농도의 저해제 물질을 포함하 는 LB에서 OD600=4.0까지 배양된 녹농균을 5 µl씩 접종하 였다. 이를 37oC에서 18-24시간 배양하면서 여과지 주위에 skim milk의 단백질이 분해되어 생기는 투명환의 직경을 통 해 프로테아제 활성을 측정하였다.
생물막 형성 측정
녹농균 생물막은 서로 다른 두 방법을 통해 측정되었다.
정지 생물막 측정(static biofilm assay)은 96-well plate에서 M63 최소배지[M63 salt(12 g/L KH2PO4, 28 g/L K2HPO4, 8 g/L (NH4)SO4), 1 mM MgSO4, 0.5% casamine acid, 0.2%
citrate]를 이용해 수행되었다. 두 개의 96-well plate에 그림 에 표시된 농도의 저해제를 넣고 M63 최소배지를 150 µl씩 분주하였다. 여기에 16시간 이상 충분히 배양된 PAO1 세포 를 2% 접종하여 37oC에서 48시간 배양하였다. 두 plate중 하나로는 OD600 값을 측정하고 또 다른 하나는 crystal violet을 염색을 통해 생물막 형성 정도를 측정하였다. Plate 에 있는 배양액을 모두 버리고 물에 2회 가볍게 씻어 well 표면에 붙어 자란 생물막 만을 남긴 후, 0.1% crystal violet 용액을 180 µl씩 분주하여 7분 동안 흔들면서 염색하였다.
남은 과량의 crystal violet 용액을 모두 버리고 absolute ethanol 200 µl로 30분 동안 흔들어 생물막 속에 염색된 crystal violet을 녹여낸 후 600 nm에서 흡광도를 측정하였 다. 측정된 A600값을 미리 측정한 OD600값으로 표준화 하 였다. 적하 유동 생물막 측정(drip-flow biofilm assay)은 녹 색 형광 단백질(GFP; green fluorescence protein)을 발현하 는 플라스미드(pAB1)를 도입한 녹농균[20]을 drip flow-cell 내에서 흘리면서 배양함에 의해 수행되었다. 우선 형광 발 현용 플라스미드인 pAB1을 녹농균 PAO1에 형질전환을 통 해 도입한 후, 이를 16시간 이상 충분히 배양하였다. Drip flow-cell내에서 생물막을 형성하기 위하여 M63 최소배지에 표시된 농도의 저해제를 넣고, 앞서 배양된 형광 발현 녹농 균을 1% 접종하여 OD600= 0.5 정도로 맞춘 다음, slide glass를 넣어둔 drip flow-cell속으로 1 mL/min의 유속으로
흘려 주었다. 초기 부착이나 생물막의 성숙 등 관찰하고자 하는 단계에 도달할 정도의 시간동안 흘려준 후 slide glass 를 꺼내어 형광 현미경(Leica MZ16F)으로 생물막이 형성된 정도를 관찰하였다.
결과 및 고찰
녹농균에서 QS 저해제들이 3OC12-HSL 수용체 단백질의 활성에 미치는 효과
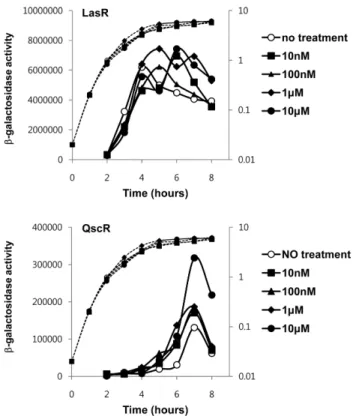
본 실험에 사용된 QS 저해 물질들인 5b와 5f의 구조는 Fig. 1에 제시되어 있다. 5b와 5f는 모두 3OC12-HSL의 수 용체인 LasR의 신호물질 결합부위에 3OC12-HSL보다 더 높 은 결합에너지(docking score)를 가지는 구조로 in silico 모 델링 분석에서 예측되었으며, 실제 대장균 QS 리포터 균주 를 이용한 실험에서 LasR의 활성을 억제하는 것이 밝혀진 물질이다[20]. 이 물질들을 실제로 녹농균에 처리했을 때, 녹 농균의 QS 반응을 주도하는 신호물질인 3OC12-HSL의 수 용체인 LasR과 QscR의 활성이 어떻게 변하는지 알아보고 자 녹농균의 성장에 따른 LasR과 QscR의 활성 변화를 측정 해 보았다. 5b의 경우 대장균 리포터 균주를 이용한 결과와 는 달리 성장 단계 전체에 걸쳐 LasR의 활성을 억제하지 못
Fig. 2. The inhibition of the LasR and QscR activities by 5b.
The lasIp-lacZ and PA1897p-lacZ fusions (pSC11 and pJL101) were introduced into the PAO1 wild type cells to monitor the activity of LasR and QscR, respectively, and cultivated with 5b at the indicated concentrations, and assayed for β-galactosidase activ- ity along the growth.
하였다(Fig. 2). LasR보다 늦게 활성이 증가하는 QscR의 경 우 예상과는 달리 5b에 의해 오히려 활성이 증가하는 것을 관찰할 수 있었다(Fig. 2). 5f의 경우에도 실험에 사용된 모 든 농도범위에서 LasR의 활성에는 영향을 주지 않는 반면, QscR의 활성은 농도 범위에 따라 증가, 혹은 감소시키는 것 으로 관찰되었다(Fig. 3). 흥미롭게도 5f는 10 µM 이하의 낮 은 농도로 처리하였을 때는 QscR의 활성을 증가시킨 반면, 그 이상에서는 QscR의 활성을 증가시키는 정도가 줄어들기 시작해서 100 µM 근방에서는 5f를 넣지 않았을 때와 비슷 한 수준이 되며, 1 mM 수준의 높은 농도에서는 QscR의 활 성을 강하게 억제하였다. 5b에서는 이러한 높은 농도에서의 QscR 활성 억제현상이 관찰되지 않았다(data not shown). 실 험에 사용된 모든 농도 범위에서 두 저해제 모두 녹농균의 성장에는 큰 영향을 미치지 않았다(Fig. 2, 3).
이 결과는 몇 가지 중요한 사실을 시사해 준다. 첫째, 대 장균을 이용한 실험에서는 5b와 5f가 모두 대장균에서 발현 된 LasR의 활성을 억제할 수 있었는데, 녹농균에서는 LasR 의 활성을 억제하지 못했다는 것은 대장균과 녹농균에서의 LasR의 접힘(folding)이 다소 다르게 일어날 가능성이 있다 는 것을 시사한다. LasR의 경우 자신의 신호물질인 3OC12- HSL과 강하게 결합하고 있어, 보통의 투석 방법으로는 LasR 로부터 3OC12-HSL을 유리시킬 수 없다는 것이 보고되어
있다[32]. 이러한 현상은 LasR 단백질이 처음 접힘을 시작 할 때 3OC12-HSL이 결합하여 들어가면서 접힘을 유도하기 때문으로 생각된다. 즉, 3OC12-HSL이 없이는 적절한 접힘 이 일어나지 않으며, 일단 3OC12-HSL과 함께 접힘이 일어 나고 나면 3OC12-HSL은 가역적으로 빠져 나오지 않는 것 으로 생각할 수 있다. 이와는 반대로 QscR의 경우는 같은 3OC12-HSL을 신호물질로 쓰면서도 3OC12-HSL의 결합과 유리가 가역적임이 밝혀져 있다[21]. 이는 QscR은 3OC12- HSL이 없이도 적절한 접힘이 일어날 수 있으며, 접힘을 유 지한 상태에서 3OC12-HSL이 다시 빠져나올 수 있음을 의 미한다. 대장균의 경우는 스스로 acyl-HSL을 만들지 못하므 로, QS 수용체를 발현시키거나 리포터를 이용한 QS 반응 측정시에 합성된 3OC12-HSL를 외부에서 첨가 하여야만 한 다. 따라서 저해제와 3OC12-HSL가 모두 외부에서 들어오 기 때문에, 처음 LasR 단백질이 합성되어 접힐 때 3OC12- HSL보다 저해제가 먼저 들어갈 수 있다. 하지만 녹농균은 스스로 3OC12-HSL을 포함한 여러 acyl-HSL을 합성하므로 3OC12-HSL은 내부에서 합성되어 있고 저해제는 외부에서 들어오기 때문에, 배양시 저해제를 빨리 넣더라도 LasR의 접힘 과정에서 일정 정도 3OC12-HSL가 우선적으로 들어갈 수 밖에 없다. LasR의 경우 일단 접힘이 일어나고 나면 그 구조가 매우 견고해서 3OC12-HSL이 빠져나올 수 없으므로 저해 효과도 나타나지 않을 수 있다고 생각된다. QscR의 경 우는 LasR의 경우처럼 3OC12-HSL이 우선적으로 접힘 과 정에 들어가더라도 다시 빠져나올 수 있으므로, 만약 저해 제가 3OC12-HSL에 비해 QscR에 대한 결합력이 더 크다면 저해제가 3OC12-HSL을 대체해 들어갈 수 있을 것이다. 따 라서 QscR에 대해서는 녹농균에서도 이들 저해물질들이 효 과를 나타낼 수 있었을 것으로 생각된다.
또 한가지 중요한 시사점은 5f의 경우 QscR에 대해서 농 도에 따라 활성을 증가시키기도, 혹은 억제하기도 했다는 점 이다. 사실 저해제도 3OC12-HSL처럼 수용체 단백질에 결 합하여 그 구조를 변화시킬 수 있지만, 3OC12-HSL의 경우 타겟 프로모터에 대한 결합과 전사활성화에 적합한 구조로 LasR이나 QscR의 구조를 변화시키는 반면, 저해물질은 프 로모터 결합이나 전사활성에 부적절한 쪽으로 수용체 단백 질의 구조를 변화시키는 것이라고 생각할 수 있다. 만약 그 구조가 프로모터 결합이나 전사활성에 적합한 형태로 수용 체 단백질의 구조를 변화시킨다면 그 물질은 오히려 활성 물 질이 될 수도 있을 것이다. 따라서 5b의 경우 QscR에 결합 하여 유도하는 구조 변화가 타겟 프로모터에 대한 결합이나 전사활성을 증가 시켰다고 생각할 수 있으며, 5f의 경우는 농도에 따라, 혹은 결합하는 5f 분자 수에 따라 QscR이 구 조가 단계적으로 변할 수 있는 경우라고 생각된다.
녹농균에서 저해제들의 프로테아제 생성에 미치는 영향 5b와 5f가 LasR과 QscR을 구별하여 농도 의존적으로 작 Fig. 3. The inhibition of the LasR and QscR activities by 5f.
The experiment was carried out with the treatment of 5f in the same way as in Fig. 2.
용하였기 때문에 이들이 QS과 관련된 중요한 표현형에 어 떻게 작용할지 판단하기 어려웠으므로, 이를 알아보기 위해 QS와 관련된 중요한 생리적 특징인 병독인자(virulence
factor)의 생산에 이들이 미치는 영향을 조사하였다. 5b와 5f 를 녹농균에 다양한 농도로 처리한 후, 대표적 병독인자인 프로테아제 활성을 skim milk 배지법을 이용하여 측정해 보
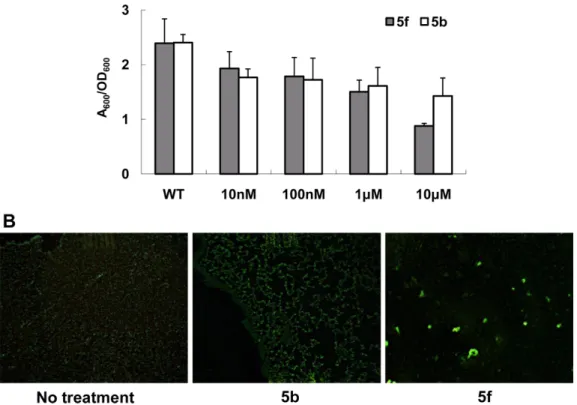
Fig. 4. Total protease activity assay. The total activity of secreted proteases was measured on skim milk plates. Either of 5b (A) or 5f (B) was treated at the different concentration as indicated.
Fig. 5. The static biofilm assay (A) and the microscopic observation of the initial attachment of PAO1 cells (B). Either of 5b or 5f was treated at the indicated concentration. The biofilm formation was measured by the absorbance at 600 nm after the crystal violet staining (A). The initial attachment of PAO1 cells that produce GFP was observed by fluorescence microscope after the treatment of 5b and 5f (B).
았을 때, 5b와 5f 모두 측정한 모든 농도 범위에서 프로테아 제 활성에 의미있는 영향을 미치지 못하였다(Fig. 4AB). 지 금까지 알려진 많은 병독인자들이 LasR의 직, 간접적인 조 절을 받는다는 점에서, 이는 앞서 5b와 5f가 LasR의 활성을 저해하지 못했다는 결과와 부합되는 결과라 할 수 있다.
QscR의 경우는 직접 조절하는 유전자중에 병독인자로 알려 진 유전자는 아직 없으나, LasR이나 RhlR등 다른 QS 수용 체 단백질들에 의해 조절되는 유전자들 중 일부의 발현을 억 제한다는 것이 보고된 바 있다[7, 22]. 본 실험과 앞서의 실 험 결과들을 종합할 때 5b나 5f에 의하여 변화된 QscR의 활 성 정도로는 LasR의 활성이나 녹농균의 병독성에 영향을 주 지 못함을 알 수 있다.
저해제들이 녹농균의 생물막 형성에 미치는 영향
저해제들이 녹농균의 생물막 형성에 미치는 영향을 알아 보기 위해 생물막 형성 정도를 조사하였다. 여러 농도로 5b 와 5f를 처리한 후, 정지 생물막 측정법(static biofilm assay) 을 이용하여 측정하였을 때, 두 저해제 모두 의미있게 녹농 균의 생물막 형성을 저해하였다. 특히 5f를 10 µM 처리한 경우 처리하지 않은 것에 비해 50% 이하로 생물막 형성이 감소하였다(Fig. 5A). 흥미롭게도 이들이 QS에 영향을 줄 때 와는 달리, 생물막 억제 효과는 낮은 농도 범위에서도 나타 났다. 이는 이 두 물질들이 생물막 억제제로 쓰일 수 있음을 의미하는 동시에, 이들에 의한 생물막 형성의 저해가 QS 저 해의 결과로 나타난 것이 아닐 수도 있음을 시사하는 것이 었다. 이를 보다 정확히 살펴보기 위해 생물막 형성 과정 동 안 시간에 따른 생물막의 성숙 정도를 현미경을 통해 관찰해 보았다. 이를 위해 녹색 형광 단백질(GFP; green fluorescence protein)을 발현하는 플라스미드(pAB1)를 도입한 녹농균[20]
을 drip flow-cell 내의 유동 배지 속에서 흘리면서 생물막을 형성시키는 적하 유동 생물막 형성 방법(drip-flow biofilm formation)과 이를 형광 형미경으로 관찰하는 방법을 통해 이 물질들이 어느 단계에서 생물막 형성을 저해하는지 조사 해 보았다. 5b의 경우 생물막 형성을 특이적으로 억제하는 특정 단계가 뚜렷이 관찰되지 않았으나, 흥미롭게도 5f의 경 우는 녹농균의 초기 부착단계를 저해하였으며(Fig. 5B), 일 단 부착이 일어난 다음 성숙하는 단계에서는 의미있는 영향 을 미치지 않았다(data not shown).
오랫동안 녹농균에서 QS과 생물막 형성이 밀접한 관계가 있다는 것이 많이 보고되어 왔음에도 불구하고[9, 29], 최근 에는 생물막 형성에 QS이외의 다른 조절인자들도 영향을 미 친다는 보고들도 많아서[15, 27], 생물막 형성은 QS와 독립 적으로도 여러 주변 인자에 의해 다양하게 조절 받을 수 있 다고 생각된다. 특히 생물막의 성숙은 QS에 의해 영향을 많 이 받을 수 있으나, 초기 부착과 같은 단계에서는 cyclic di- GMP와 같은 세포내 신호전달 물질들의 영향을 많이 받는 다는 것이 알려져 있다[28]. 본 실험에서 5b와 5f가 QS에 영
향을 주는 농도보다 훨씬 낮은 농도에서 생물막 형성을 억 제한 것이나, 5f의 경우 생물막의 성숙이 아닌 초기 부착을 억제하였다는 점에서 이들이 QS와는 독립적인 경로로 생물 막 형성을 억제하였을 가능성이 매우 높다고 생각된다.
본 연구는 세균 QS 저해제 개발을 위한 지속적 연구의 일 환으로 이전 연구에서 in silico 모델링을 이용한 스크리닝을 통해 QS 저해제로 발굴된 두 가지 물질에 대한 보다 심층 적인 평가를 위한 것이었다. 결론적으로 이들 화합물들이 대 장균 리포터 균주에서는 3OC12-HSL 수용체중 LasR의 활 성을 억제한 것과는 달리, 녹농균에서는 LasR보다는 녹농균 의 또 다른 3OC12-HSL 수용체인 QscR의 활성에 주된 영 향을 주며, 그 영향도 단순한 활성 억제라기 보다는 5b의 경 우 활성 증가이거나, 혹은 5f의 경우처럼 농도에 따른 활성 억제였음을 알 수 있었다. 이러한 차이는 대장균과 녹농균 두 균주간에 단백질의 접힘이 다르게 나타나거나, 혹은 5b 나 5f의 대사 과정이 달라 서로 다른 물질로 변환되기 때문 일 수 있다. 이에 대한 보다 자세한 기전 연구가 필요하다고 생각된다. 또한 이들 물질이 녹농균의 생물막 억제 효과가 있음을 새로이 발견하였는데, 이들 물질들이 QS에 영향을 주더라도 대표적 병독소인 프로테아제의 발현에 영향을 주 지는 못하였으므로, 녹농균의 병독성을 증가시키지 않으면 서 생물막을 억제 할 수 있는 물질로 이들을 개발할 수 있 을 가능성이 있다고 생각된다.
요 약
그람음성 간균인 녹농균(Pseudomonas aeruginosa)은 비 뇨기, 각막, 호흡기, 화상부위 등에 광범위하게 감염하는 기 회감염성 병원균으로, 병원성의 발현에 세균의 세포밀도 인 식 기전인 쿼럼 센싱(quorum sensing)이 매우 중요하게 관 여한다. 사전 연구에서 녹농균 감염력을 제어하기 위한 방 법으로 쿼럼 센싱의 주 신호물질인 N-3-oxododecanoyl-HSL (3OC12-HSL)의 분자 구조가 변형된 물질들을 합성하여 쿼 럼 센싱 억제물질로 사용하고자 하였으며, 그 중 두 개의 물 질들(5b, 5f)이 대장균을 이용한 스크리닝을 통해 녹농균의 주요 쿼럼 센싱 수용체 단백질인 LasR의 활성을 억제할 수 있음을 확인하였었다. 본 연구에서는 이 물질들의 효과를 보 다 면밀히 분석하기 위하여 실제 녹농균에서 이 물질들이 쿼 럼 센싱과 병독성을 억제할 수 있는지 분석해 보았다. 대장 균을 이용한 리포터 분석에서와는 달리, 5b와 5f 모두 녹농 균에서 직접 처리하였을 때는 LasR의 활성에 영향을 주지 못하였다. 대신 이 물질들은 녹농균의 또다른 쿼럼 센싱 수 용체 단백질인 QscR의 활성에 선택적으로 영향을 주었다.
흥미롭게도 이 물질들의 효과는 대장균에서 얻어진 결과와 는 달랐으며 다소 복잡하였다. 두 물질 모두 낮은 농도 범위 (<10 µM)에서 QscR의 활성을 증가시켰으며, 높은 농도의 5f(≈1 mM)는 QscR을 강하게 억제하였다. 두 물질 모두 중
요한 병독인자인 프로테아제 활성에는 영향을 주지 않으면 서도, 만성감염을 매개하는데 중요한 생물막의 형성은 의미 있게 감소시켰다. 특히 5f는 생물막의 성숙단계 보다는 녹농 균 세포의 초기 부착을 억제하였다. 이러한 결과들을 바탕 으로, 5f의 경우 독성의 증가 없이 생물막 형성을 억제할 수 있는 물질로 응용이 가능하다고 제안한다.
Acknowledgements
This work was supported by Korea Ministry of Environ- ment as "The Eco-technopia 21 project" (102-081-067) and the Korea Science and Engineering Foundation (KOSEF) grant funded by the Korea Government (MOST) (R01-2007- 000-20732-0). This work was also supported by the Korea Research Foundation Grant funded by the Korean Govern- ment (MOEHRD) (KRF-2007-331-C00222) and National Foundation of Korea Gront funded by the Korean Govern- ment (2010-0015901).
REFERENCES
1. Abdel-Mawgoud, A. M., F. Lepine, and E. Deziel. 2010.
Rhamnolipids: Diversity of structures, microbial origins and roles. Appl Microbiol Biotechnol. 86: 1323-1336.
2. Alionte, L. G., B. M. Cannon, C. D. White, A. R. Caballero, R. J. O'Callaghan, and J. A. Hobden. 2001. Pseudomonas aeruginosa lasa protease and corneal infections. Curr Eye Res. 22: 266-271.
3. Balaban, N., A. Giacometti, O. Cirioni, Y. Gov, R. Ghiselli, F. Mocchegiani, C. Viticchi, M. S. Del Prete, V. Saba, G.
Scalise, and G. Dell'Acqua. 2003. Use of the quorum- sensing inhibitor rnaiii-inhibiting peptide to prevent biofilm formation in vivo by drug-resistant staphylococcus epidermidis. J Infect Dis. 187: 625-630.
4. Baveja, J. K., G. Li, R. E. Nordon, E. B. Hume, N. Kumar, M. D. Willcox, and L. A. Poole-Warren. 2004. Biological performance of a novel synthetic furanone-based antimicrobial.
Biomaterials. 25: 5013-5021.
5. Blackwood, L. L., R. M. Stone, B. H. Iglewski, and J. E.
Pennington. 1983. Evaluation of pseudomonas aeruginosa exotoxin a and elastase as virulence factors in acute lung infection. Infect Immun. 39: 198-201.
6. Bruno, T. F., D. E. Woods, and C. H. Mody. 2000.
Exoenzyme s from pseudomonas aeruginosa induces apoptosis in t lymphocytes. J Leukoc Biol. 67: 808-816.
7. Chugani, S. A., M. Whiteley, K. M. Lee, D. D'Argenio, C.
Manoil, and E. P. Greenberg. 2001. Qscr, a modulator of quorum-sensing signal synthesis and virulence in pseudomonas aeruginosa. Proc Natl Acad Sci U S A. 98: 2752-2757.
8. Cos, P., K. Tote, T. Horemans, and L. Maes. 2010. Biofilms:
An extra hurdle for effective antimicrobial therapy. Curr Pharm Des. 16: 2279-2295.
9. Davies, D. G., M. R. Parsek, J. P. Pearson, B. H. Iglewski, J.
W. Costerton, and E. P. Greenberg. 1998. The involvement of cell-to-cell signals in the development of a bacterial biofilm. Science. 280: 295-298.
10. Dietrich, L. E., A. Price-Whelan, A. Petersen, M. Whiteley, and D. K. Newman. 2006. The phenazine pyocyanin is a terminal signalling factor in the quorum sensing network of pseudomonas aeruginosa. Mol Microbiol. 61: 1308-1321.
11. Diggle, S. P., P. Cornelis, P. Williams, and M. Camara. 2006.
4-quinolone signalling in pseudomonas aeruginosa: Old molecules, new perspectives. Int J Med Microbiol. 296: 83- 91.
12. Dong, Y. H. and L. H. Zhang. 2005. Quorum sensing and quorum-quenching enzymes. J Microbiol. 43 Spec No, 101- 109.
13. Francolini, I. and G. Donelli. 2010. Prevention and control of biofilm-based medical-device-related infections. FEMS Immunol Med Microbiol. 59: 227-238.
14. Hentzer, M., H. Wu, J. B. Andersen, K. Riedel, T. B.
Rasmussen, N. Bagge, N. Kumar, M. A. Schembri, Z. Song, P. Kristoffersen, M. Manefield, J. W. Costerton, S. Molin, L.
Eberl, P. Steinberg, S. Kjelleberg, N. Hoiby, and M. Givskov.
2003. Attenuation of pseudomonas aeruginosa virulence by quorum sensing inhibitors. EMBO J. 22: 3803-3815.
15. Heydorn, A., B. Ersboll, J. Kato, M. Hentzer, M. R. Parsek, T. Tolker-Nielsen, M. Givskov, and S. Molin. 2002. Statistical analysis of pseudomonas aeruginosa biofilm development:
Impact of mutations in genes involved in twitching motility, cell-to-cell signaling, and stationary-phase sigma factor expression. Appl Environ Microbiol. 68: 2008-2017.
16. Johnson, M. K. and D. Boese-Marrazzo. 1980. Production and properties of heat-stable extracellular hemolysin from pseudomonas aeruginosa. Infect Immun. 29: 1028-1033.
17. Kim, C., J. Kim, H. Y. Park, J. H. Lee, H. J. Park, C. K.
Kim, and J. Yoon. 2009. Structural understanding of quorum-sensing inhibitors by molecular modeling study in pseudomonas aeruginosa. Appl Microbiol Biotechnol. 83:
1095-1103.
18. Kim, C., J. Kim, H. Y. Park, R. J. McLean, C. K. Kim, J.
Jeon, S. S. Yi, Y. G. Kim, Y. S. Lee, and J. Yoon. 2007.
Molecular modeling, synthesis, and screening of new bacterial quorumsensing antagonists. J Microbiol Biotechnol.
17: 1598-1606.
19. Kim, C., J. Kim, H. Y. Park, H. J. Park, C. K. Kim, J. Yoon, and J. H. Lee. 2009. Development of inhibitors against trar quorum-sensing system in agrobacterium tumefaciens by molecular modeling of the ligand-receptor interaction. Mol Cells. 28: 447-453.
20. Kim, C., J. Kim, H. Y. Park, H. J. Park, J. H. Lee, C. K.
Kim, and J. Yoon. 2008. Furanone derivatives as quorum- sensing antagonists of pseudomonas aeruginosa. Appl Microbiol Biotechnol. 80: 37-47.
21. Lee, J. H., Y. Lequette, and E. P. Greenberg. 2006. Activity of purified qscr, a pseudomonas aeruginosa orphan quorum- sensing transcription factor. Mol Microbiol. 59: 602-609.
22. Lequette, Y., J. H. Lee, F. Ledgham, A. Lazdunski, and E. P.
Greenberg. 2006. A distinct qscr regulon in the pseudomonas aeruginosa quorum-sensing circuit. J Bacteriol. 188: 3365- 3370.
23. Manefield, M., T. B. Rasmussen, M. Henzter, J. B. Andersen, P. Steinberg, S. Kjelleberg, and M. Givskov. 2002.
Halogenated furanones inhibit quorum sensing through accelerated luxr turnover. Microbiology. 148: 1119-1127.
24. Matsumoto, K. 2004. Role of bacterial proteases in pseudomonal and serratial keratitis. Biol Chem. 385: 1007- 1016.
25. Page, M. G. and J. Heim. 2009. Prospects for the next anti- pseudomonas drug. Curr Opin Pharmacol. 9: 558-565.
26. Park, S. J., H. Y. Chung, and J. H. Lee. Rapid in vivo screening system for anti-oxidant activity using bacterial redox sensor strains. J Appl Microbiol. 108: 1217-1225.
27. Parsek, M. R. and E. P. Greenberg. 2005. Sociomicrobiology:
The connections between quorum sensing and biofilms.
Trends Microbiol. 13: 27-33.
28. Pesavento, C. and R. Hengge. 2009. Bacterial nucleotide- based second messengers. Curr Opin Microbiol. 12: 170- 176.
29. Purevdorj, B., J. W. Costerton, and P. Stoodley. 2002.
Influence of hydrodynamics and cell signaling on the structure and behavior of pseudomonas aeruginosa biofilms.
Appl Environ Microbiol. 68: 4457-4464.
30. Reading, N. C. and V. Sperandio. 2006. Quorum sensing:
The many languages of bacteria. FEMS Microbiol Lett. 254:
1-11.
31. Schuster, M. and E. P. Greenberg. 2006. A network of networks: Quorum-sensing gene regulation in pseudomonas aeruginosa. Int J Med Microbiol. 296: 73-81.
32. Schuster, M., M. L. Urbanowski, and E. P. Greenberg. 2004.
Promoter specificity in pseudomonas aeruginosa quorum
sensing revealed by DNA binding of purified lasr. Proc Natl Acad Sci U S A. 101: 15833-15839.
33. Smith, K. M., Y. Bu, and H. Suga. 2003. Library screening for synthetic agonists and antagonists of a pseudomonas aeruginosa autoinducer. Chem Biol. 10: 563-571.
34. Stover, C. K., X. Q. Pham, A. L. Erwin, S. D. Mizoguchi, P.
Warrener, M. J. Hickey, F. S. Brinkman, W. O. Hufnagle, D.
J. Kowalik, M. Lagrou, R. L. Garber, L. Goltry, E. Tolentino, S. Westbrock-Wadman, Y. Yuan, L. L. Brody, S. N. Coulter, K. R. Folger, A. Kas, K. Larbig, R. Lim, K. Smith, D.
Spencer, G. K. Wong, Z. Wu, I. T. Paulsen, J. Reizer, M. H.
Saier, R. E. Hancock, S. Lory, and M. V. Olson. 2000.
Complete genome sequence of pseudomonas aeruginosa pao1, an opportunistic pathogen. Nature. 406: 959-964.
35. Suman, E., S. Varghese, and J. Jose. 2005. Gentamicin resistance in biofilm producing pseudomonas aeruginosa causing catheter associated urinary tract infections. Indian J Med Sci. 59: 214-216.
36. Tang, A., M. E. Marquart, J. D. Fratkin, C. C. McCormick, A. R. Caballero, H. P. Gatlin, and R. J. O'Callaghan. 2009.
Properties of pasp: A pseudomonas protease capable of mediating corneal erosions. Invest Ophthalmol Vis Sci. 50:
3794-3801.
37. Welch, M., H. Mikkelsen, J. E. Swatton, D. Smith, G. L.
Thomas, F. G. Glansdorp, and D. R. Spring. 2005. Cell-cell communication in gram-negative bacteria. Mol Biosyst. 1:
196-202.
38. Williams, P. 2007. Quorum sensing, communication and cross-kingdom signalling in the bacterial world. Microbiology.
153: 3923-3938.
39. Ymele-Leki, P. and J. M. Ross. 2007. Erosion from staphylococcus aureus biofilms grown under physiologically relevant fluid shear forces yields bacterial cells with reduced avidity to collagen. Appl Environ Microbiol. 73: 1834-1841.