A Functional Analysis of OsCPK11, a Calcium-dependent Protein Kinase (CDPK) Gene in Rice
Su-Hee Lee
1, Jeong-Eun Lee
2, Philip Day
3, Simon Gilroy
4and Sung-Ha Kim
5*
1Iwol Middle School, 753 Jingwang-ro, Iwol-myeon, Jincheon-gun, Chungbuk 27813, Korea
2Keoje Girls’ Middle School, 43 Geumyong-ro, Yeonje-gu, Busan 47529, Korea
3Department of Mathmatics and Natural Sciences, Penn State Altoona, 3000 Ivyside Park, Altoona, PA16601, USA
4Department of Botany, University of Wisconsin-Madison, 430 Lincoln Dr., Madison, WI53706, USA
5Department of Biology Education, Korea National University of Education, 250 TaesungTabyeon-ro, Gangnae-myeon, Heungdeok-gu, Cheongju-si, Chungbuk 28173, Korea
Received September 19, 2016 /Revised October 30, 2017 /Accepted October 31, 2017
CDPKs have pivotal roles in plant Ca
2+-mediated transduction signaling. A total of 29 CDPK genes have been identified in rice (Oryza sativa L.), but their key functions have not been completely noted.
This study focused on the OsCPK11 gene, which has not been studied, to determine its functional characteristics. A study of tissue-specific expressions revealed that the OsCPK11 gene is expressed in young leaves, mature leaves and flowers of rice. An expression of the gene was also confirmed in gibberellin-treated aleurone layers of rice. Regarding the phenotypic characteristics of Tos17-inserted OsCPK11 mutants, the heights of the mutants were not distinguishable from the heights of wild type plants, but the number of caryopses and the caryopses' weights were significantly statistically different. In addition, many grains of the mutants had white belly materials in their endosperm. The cDNA of the OsCPK11 was cloned, and an OsCPK11 protein of about 60.5 kD was obtained by using a GST affinity chromatography and an SDS-PAGE. An analysis of the amino-acid sequence of the pro- tein indicated that the OsCPK11 protein has the structural characteristics of typical CDPKs. The results provided useful information about the functions of the OsCPK11 gene and further noted the roles CDPKs have in Ca
2+-mediated signaling in plants.
Key words : Calcium-dependent protein kinase, gibberellins, OsCPK11, rice aleurone layers, white
belly materials
*Corresponding author
*Tel : +82-43-230-3738, Fax : +82-43-232-7176
*E-mail : [email protected]
This is an Open-Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Life Science 2017 Vol. 27. No. 11. 1233~1244 DOI : https://doi.org/10.5352/JLS.2017.27.11.1233
Introduction
All organisms control their metabolism and adapt to their environment through a network of signal transduction pathways. Among these pathways, Ca
2+plays a pivotal role as a universal second messenger [7, 42, 50]. In hormone-sen- sitive cells, neurons, muscle cells, and other cells that re- spond to extracellular signals, Ca
2+triggers the intracellular response. Among the processes triggered by Ca
2+are exocy- tosis in nerve and endocrine cells and contraction in muscle [30]. Cytosolic Ca
2+concentrations are normally kept low by the action of Ca
2+pumps in the endoplasmic reticulum, mi-
tochondria, and plasma membrane [10], but hormonal, neu- ral, or other stimuli can cause an influx of Ca
2+into the cell through specific Ca
2+channels in the plasma membrane or by the release of sequestered Ca
2+from the endoplasmic re- ticulum or mitochondria, raising cytosolic Ca
2+levels and triggering a cellular response [37].
In plants, when intracellular Ca
2+increases in response to a variety of stimuli such as light, hormones, pathogens, and stress [18, 21, 27, 43], Ca
2+sensors or Ca
2+-binding pro- teins recognize the change in Ca
2+levels; in turn, they induce downstream effects including altered protein phosphor- ylation and gene expression [42].
Many kinds of Ca
2+sensors are found in plants. Major plant Ca
2+sensors include calmodulin (CaM), calcineurin B-like proteins and Ca
2+-dependent protein kinases (CDPKs or CPKs) [12, 13, 28]. Of these three sensors, CDPKs are of special interest because they represent a novel type of Ca
2+sensors and effectors.
CDPKs consist of variable N-terminal domains and sev-
eral functional domains, including a protein kinase domain,
an autoinhibitory region and a CaM-like domain [11, 19, 20, 24]. The CaM-like domain generally contains four EF hands to which Ca
2+can be bound. Thus, the activity of CDPKs is directly dependent on Ca
2+, but not on CaM. When the CaM-like domain is not occupied by Ca
2+, the autoinhibitory domain which contains a pseudosubstrate sequence can in- teract with the active site of the kinase domain and inhibit enzyme activity [51]. The binding of Ca
2+to the CaM-like domain leads to structural changes in the autoinhibitory do- main, resulting in exposure of the active site of the kinase [14]. Activated CDPKs take part in different signal trans- duction pathways by phosphorylating specific substrates [23, 54]. Although the biochemical activities of the CDPK family have been characterized, the biological functions of in- dividual plant CDPKs are not known very well [33]. Putative functions of CDPKs include roles in growth, development and differentiation of plants, regulation of plant tran- scription, responses to phytohormones, plant-stress signal transduction and defense responses [35]. Tissue-specific ex- pression of CDPK isoforms suggests their involvement in early developmental processes including embryogenesis, seed development and germination [2, 3]. The participation of CDPK isoforms in tuber development was studied in po- tato (Solanum tuberosum L.) [39]. In addition, CDPKs modu- late Ca
2+-dependent plant responses caused by phyto- hormones such as gibberellins, cytokinins and auxins.
Activity of partially purified rice CDPK is 10-fold higher in GA-treated seeds than in untreated ones [1]. Moreover, GA treatment leads to an increase in the transcript levels of OsCDPK13, another CDPK, in rice seedlings [52].
Rice (Oryza sativa L.), one of the most important food plants in the world, feeds over half of the global population and has played a central role in human culture for the past 10,000 years [25]. Recently, the genome sequence of Oryza sativa L. ssp. japonica cv Nipponbare has been determined [25]. Rice is an important model plant in plant molecular biology because of its small genome size among cereals, dense genetic maps and relative ease of genetic trans- formation [44]. Therefore, these properties are expected to set the stage for a complete functional characterization of the rice genome along with the finished sequence and other tools under development.
Twenty-nine rice CDPK genes have been identified using a bioinformatics analysis of the entire rice genome, and rice CDPK genes are distributed among all the rice chromosomes except chromosome 6 [5]. Many of the rice CDPKs are in-
duced mainly by a variety of stimuli such as cold, drought, salt, elicitors and hormones [51]. For instance, OsCPK12 pos- itively regulates salt tolerance and negatively modulates blast disease resistance [6], OsCPK13 is involved in cold, salt and drought tolerance [41], and OsCPK21 is likely involved in the positive regulation of abscisic acid and salt signaling pathways [6]. Other CDPKs play roles in the development of each tissue in rice. For example, OsCPK23 affects seed development, especially during storage product accumu- lation [4], and eight CDPK genes including OsCPK11 are in- volved with pollen development [53]. Many investigators have studied rice CDPKs, but the exact functions of some rice CDPKs–especially OsCPK11–have not been identified.
Therefore, we focused on the functional characterization of the OsCPK11 gene. In this investigation, the expression of OsCPK11 gene from both rice tissues and hormone-treated rice aleurone layers was measured. Some phenotypic fea- tures of the Tos17–insertion mutants of OsCPK11 were also analyzed. Finally, a rice cDNA encoding OsCPK11 was cloned from flowers, and the OsCPK11 protein was partially characterized.
Materials and Methods
Plant materials and growth conditions
The National Institute of Agrobiological Science in Japan kindly provided 20 seeds each of two mutant rice lines (ND0001 and NF6012) in which retrotransposon Tos17 was inserted into the OsCPK11 gene. Nipponbare wild type seeds were also used. Seeds were surface-sterilized with 2% so- dium hypochlorite and 0.1% Tween-20. They were then ger- minated in distilled water at room temperature for 24 hr in the dark. Each germinated seed was planted in a plastic pot. They were watered daily and were grown in the green- house at 25~38°C for about 6 months. In order to investigate tissue-specific expression of the OsCPK11 gene, tissues such as young leaves, mature leaves, roots and flowers of wild type plants were collected when the plants reached a specific developmental stage. For the genotyping of the mutant lines, young leaves of each individual were collected. All of the samples were immediately frozen in liquid nitrogen and stored at -80°C until used.
Tissue-specific expression of the OsCPK11 gene in rice
Young leaves that looked yellowish green were obtained
at about 10 days after planting, when an individual's height reached 15 cm. Mature leaves that looked dark green were collected at about 8 weeks after planting, when an in- dividual's height reached 50 cm. Roots were obtained from seedlings that were grown in the dark for 2 weeks at 27~28°C, and flowers were collected right after blooming.
Aleurone layers were obtained by removing starch material from the grain endosperm.
Total RNA was extracted with TRIzol reagent following the manufacturer's instructions (Invitrogen Inc., USA).
About 100 mg of each tissue sample was ground into fine powder with liquid nitrogen, a mortar and pestle. Each ground sample was transferred to a 1.5 ml microtube with 1 ml of TRIzol reagent and vortexed vigorously. After cen- trifugation at 12,000x g for 10 min at 6°C , the RNA-contain- ing supernatant was transferred to a fresh tube and was in- cubated for 5 min at room temperature (20~25°C) to permit the complete dissociation of the nucleoprotein complexes.
Next, 0.2 ml of chloroform was added to the supernatant, and the tube was shaken vigorously by hand for 15 sec. It was incubated at room temperature for 2 to 3 min and then centrifuged at 12,000x g for 13 min at 6°C. Centrifugation, separated the mixture into a lower red phenol-chloroform phase, an interphase, and an upper colorless aqueous phase.
RNA remains exclusively in the aqueous phase, which was transferred to a fresh tube. For precipitation of the RNA from the aqueous phase, 0.5 ml of isopropyl alcohol was added. The tube was incubated at room temperature for 10 min and centrifuged at 12,000 xg for 9 min at 6°C. After removal of the supernatant, the RNA pellet was washed once with 1 ml of 75% ethanol and was dried at 37°C for 30 min. The dried RNA pellet was dissolved in 4~15 μl of diethyl pyrocarbonate-distilled water (DEPC-DW, Bioneer Inc., Korea), and then it was stored at -80°C until used.
cDNA was synthesized using DNaseⅠ and SuperscriptⅡ RT following the manufacturer's technical manual (Invitro- gen Inc., USA). Each resuspended RNA sample was diluted with DEPC-DW to make 1 μg/μl stock. Then, 1 μl of the 1 μg/μl RNA stock sample was combined with 1 μl 10X DNaseⅠ Reaction Buffer, 1 μl DNaseⅠ(1U/μl) and 7 μl DEPC-DW in an RNase-free microtube on ice. The mixture was incubated for 14 min at room temperature. To inactivate the DNaseⅠ, 1 μl of 25 mM EDTA solution was added to the mixture above, and the tube was heated for 10 min at 65°C. The RNA sample was ready to use in reverse tran- scription, prior to amplification. 1 μl of oligo-dT (0.25 μg/μl)
was added to the 11 μl volume from above, and the mixture was heated at 65℃ for 10 min and cooled at room temper- ature for 10 min. To prepare the Reverse Transcriptase(RT) Master Mix of total RNA samples, 4 μl 5X 1st Strand Buffer, 2 μl 0.1M dithiothreitol (DTT), 1.5 μl 10 mM dNTP mix, 1.5 μl (200 U/μl) SuperscriptⅡ RT, and 0.5 μl RNase Inhibitor per each RNA sample were mixed in 1.5 ml microtubes on ice. Then, 8 μl of RT Master Mix was added to each RNA sample, and this mixture was incubated at 42°C for 1 hr to synthesize the cDNA. The synthesis was then inactivated at 65°C for 15 min. Samples made by the process above were used in PCR. cDNA was amplified in a volume of 25 μl con- taining 17.5 μl sterilized DW, 2.5 μl 10X Taq buffer, 1 μl 10 mM dNTP mix, 1 μl forward primer (10 pmoles/μl), 1 μl reverse primer (10 pmoles/μl), 1 μl Taq DNA polymerase (5,000 U/ml) and 1 μl of cDNA RT product. Thermocycling for PCR was designed as follows: 94°C for 5 min for 1 cycle, 94°C for 1 min, 50°C for 1 min, 72°C for 1 min for 38 cycles, 72°C for 5 min for 1 cycle. The pairs of specific primer sets for the OsCPK11 gene and rice actin gene were constructed, and the specificity of primer sequences was determined us- ing the Basic Local Alignment Search Tool of the National Center for Biotechnology Information (NCBI). The primers used to confirm OsCPK11 transcripts were CF (CCTCTTCA TCCACCAGAAGG) and CR (TTCCGGT-ACTATGCTGACCC).
The primers used to confirm actin transcripts were AF (CATCTTGGCATCTCTCAGCAC) and AR (AACTTTGTCC ACGCTAATGAA). PCR products were separated on 0.8%
agarose gels and photographed under a UV light.
Expression of the OsCPK11 gene in hormone-treated rice aleurone layers
Rice aleurone layers were isolated using the methods de- scribed in Day [16]. To resolve the starchy endosperm, 20~30 ml of Endosperm Cellulase Solution (5% Onozuka R-10 cel- lulase, 1% PVP-K25, 10 mM CaCl
2, 5 mM MES, pH 5.5) was prepared. Peeled rice seeds were surface-sterilized with 99.9% ethanol in a 50 ml tube for 15 sec. The embryo and the opposite end of its grain were dissected out with a razor blade, and the remaining sections were sterilized with 2%
sodium hypochlorite and 0.1% Tween-20 in a 50 ml tube
for 5 min. Then, the sections were rinsed several times with
sterilized DW. Filtered Endosperm Cellulase Solution was
added to the rice sections, and they were transferred to a
sterilized petri dish. The petri dish was sealed with parafilm
and aluminum foil and was incubated in a 65 rpm shaker
at 32°C for 24 hr. The following day, the rice sections were softly pressed with a sterilized spatula and rinsed several times with sterilized DW. Rice aleurone layers prepared by this procedure were used for the hormone treatment.
Treatments with the plant hormones GA and/or abscisic acid (ABA) were performed in like manner as Day [16].
Groups of 30 rice aleurone layers were transferred to small petri dishes. To each dish, 2 ml of the filtered Rice Aleurone Buffer (10 mM CaCl
2, 5 mM MES, pH 5.5) with plant hor- mones (10 μM GA and/or 10 μM ABA) was added, and the dishes were incubated at 32℃ for 3 days. 50 μl samples from each petri dish were taken every 24 hr for 3 days and were stored at 4°C.
α-Amylase activity was assayed as described in Bush and Jones [9]. 20 ml of each stored sample was transferred to a fresh microtube with 200 ml of Starch Solution (0.15% pota- to starch, 44.1 mM KH
2PO
4, 20 mM CaCl
2). After the tubes were incubated at 37°C for 8~15 min, 200 μl of I
2+KI solution [1 ml iodine stock solution (47 mM I
2and 361 mM KI) in 99 ml 0.05 N HCl] was put into each tube. A 420 μl aliquot of each sample in the tube was placed into a cuvette contain- ing 1ml sterilized DW. The absorbance was determined at 625 nm.
To determine expression of the OsCPK11 gene in rice aleurone layers, aleurone layers with or without 72-hr ex- posure to plant hormones were used. They were tested in the same manner as that for tissue- specific expression of the OsCPK11 gene.
Genotyping and phenotyping of OsCPK11 mutants Genotypes of two OsCPK11 mutant lines were determined.
For DNA extraction from young leaves, a phenol-free DNA minipreparation method was used as described in Day [16].
About 50 mg of each tissue sample was ground using liquid nitrogen, mortar and pestle. The ground samples were trans- ferred to 1.5 ml microtubes containing 700 μl of pre-warmed (65°C) Extraction Buffer (2% CTAB, 1.42 M NaCl, 20 mM EDTA, 100 mM Tris-HCl, 2% PVP-40, 5 mM ascorbic acid, pH 8.0), vortexed briefly, and incubated at 65°C. Fifty Units of RnaseⅠ were added to each tube, with incubation at 65°C for 5 min. After addition of 570 μl chloroform:isoamyl alco- hol mixture (24:1), the mixture was centrifuged at full speed (13,000 rpm) at room temperature for 10 min. The upper phase containing DNA was transferred to a new microtube.
DNA was precipitated by adding 400 μl of isopropanol and spinning at full speed (13,000 rpm) for 5 min, and the super-
natant was removed. The DNA pellet was washed with 200 μl of 70% ethanol and air-dried. Then, it was resuspended in 15 μl of sterilized DW. PCR was performed as follows:
94°C for 4 min for 1 cycle, 94°C for 45 sec, 55.2°C(or 57°C) for 45 sec, 72°C for 90 sec for 31 cycles, 72°C for 5 min for 1 cycle. The primer pairs for genotyping were (TTACAAGT ACCCACCG-GCTC) as a forward primer and (GCATGCAT GATCCAAATCAG) as a reverse primer for ND0001, (ACCA TGTCATT-GTGGTTTGC) as a forward primer and (AACAT CAACATGAAAGCAGCC) as a reverse primer for NF6012, and (GAGAGCATCATCGGTTACATCTTCTC) for Tos17, respectively. The master-mix components for the DNA am- plification were 0.3 μl 10mM dNTP mix, 1.5 μl 10×Taq buffer, 0.15 μl Taq DNA polymerase, 0.75 μl 10 μM Tos17 primer, 0.75 μl 10 μM ND0001(or NF6012) forward primer, 0.75 μl 10 μM ND0001(or NF6012) reverse primer, 10.7 μl distilled H
2O and 0.1 μl of the DNA template.
The phenotypic features analyzed in both oscpk11 mutant lines and wild type plants were plant growth, caryopsis number and caryopsis weight. Plant growth was determined by measuring individual height every 5 days during the 50 days after planting. Caryopsis number per panicle was con- firmed by counting the number of caryopses in each of 10 panicles per individual. Caryopsis weight was examined by measuring the weight of 100 caryopses individually in each mutant line. In addition, the presence of white belly materi- als in cross sections of 100 grains of each mutant line was determined. One-way analysis of variance (ANOVA) was conducted using SPSS (version 12.0).
Cloning of full-length cDNA encoding OsCPK11 cDNA encoding OsCPK11 was amplified using primers whose sequences are F (AAAGGATTCATGGGCAACAACT GCGTCGG) and R(AAACTCGAGTTACCGAAGT-TTTAGC GCTTCC), with BamHⅠ and XhoⅠ sites for the insertion of 1,731 bp of OsCPK11 cDNA into a pET-41a(+) expression vector. A mixture of 28.5 μl sterilized DW, 10 μl 5X GC buf- fer, 4 μl 10 mM dNTP mix, 3 μl cDNA from flowers, 2.5 μl F/R primer (10 pmoles/μl), 1.5 μl dimethyl sulfoxide, and 0.5 μl of phusion DNA polymerase (5,000 U/ml) was used for PCR, performed as follows: 98°C for 1 min for 1 cycle, 98°C for 10 sec, 58°C for 30 sec, 72°C for 90 sec for 38 cycles, 72°C for 7 min for 1 cycle. PCR products were separated by an agarose gel, and the 1750 bp band was eluted using a Wizard SV gel and PCR clean-up system (Promega, USA).
pET-41a(+) DNA was prepared as a vector for cloning. This
vector was transformed into DH5α competent cells. The transformant was cultured in Luria-Bertani (LB) medium (Merck, Germany) with kanamycin, and the vectors were purified using a Wizard plus SV miniprep DNA purification system (Promega, USA).
The amplified cDNA and vector were double-digested with BamHⅠ and XhoⅠ, according to the manufacturer's manual (New England BioLabs, USA). The restriction en- zymes and the buffer were added, and the mixture was in- cubated at 37℃ for 2 hr. cDNA digested by restriction en- zymes was ligated into the pET-41a(+) vector at 4°C overnight. For this, 15 μl cDNA (95 ng/μl), 2 μl vector (121 ng/μl), 1μl T4 DNA polymerase (400 U/μl) and 2 μl 10×
reaction buffer were mixed according to procedure.
Transformation using Escherichia coli (E. coli) was carried out as described previously [15]. 10 μl of ligation product was added to a tube containing DH5α competent cells, and the tube was left on ice for 30 min. The tube was then trans- ferred to a rack placed in a heat block and stored for exactly 90 sec. 1ml of LB medium was added to the tube, and the mixture was incubated at 37°C for 1 hr to allow the bacteria to recover and to express the antibiotic resistance marker encoded by the plasmid. 100 μl of transformed competent cells were transferred onto agar LB medium with the appro- priate antibiotics, and the plate was stored at room temper- ature until all liquid was absorbed. The inverted plate was incubated at 37°C, and transformed colonies appeared in 12~16 hr.
Sixteen bacterial colonies from a plate were transferred to each tube along with 25 μl sterilized DW, and the tubes were mixed softly. Some solution of each colony was used for the PCR. PCR reactants included 0.2 μl 5X buffer, 0.5 μl F/R primer, 0.5 μl 10 mM dNTP mix, 2 μl Taq DNA poly- merase (5 U/μl), 2 μl of the colony solution and 4.8 μl steri- lized DW. This was kept at 95°C for 5 min and subjected to 20 cycles of 95°C for 30 sec, 55°C for 30 sec and 72°C for 90 sec. Afterward, the resulting mixture was kept at 72°C for 5 min to stabilize the amplified DNA.
After PCR, the positive transformed cells were cultured, and pET41a(+)-OsCPK11 complex DNA was extracted using the Wizard plus SV miniprep DNA purification system (Promega, USA). A part of purified plasmid DNA was used for the sequencing. Sequencing was performed by the Genome Research Facility at Seoul National University (Seoul, Korea), and the result was analyzed through multiple sequence alignment by Florence Corpet (http://bioinfo.gen
otoul.fr/multalin/).
Expression and purification of OsCPK11
After E. coli strain BL21 was retransformed with the pET41a(+)-OsCPK11, the transformed colony was inoculated in 5 ml LB medium containing 50 μg/μl kanamycin, and cultured overnight at 37°C to obtain a saturated culture. 500 μl of saturated culture was again inoculated with 500 ml LB medium containing 50 μg/μl kanamycin in a 2 l flask and incubated with shaking at 37°C until the culture has reached the mid-log phase of growth (A
600=0.5~1.0). Isopropyl- β-D-thiogalactopyranoside (IPTG) was added to reach a final concentration of 0.2 mM to express the OsCPK11 protein.
The cells were grown at 30°C for 5 hr, and they were har- vested by centrifugation at 4,200x g for 15 min at 4°C. The cell pellet was resuspended in 10ml PBS (137 mM NaCl, 10 mM Na
2HPO
4, 2.7 mM KCl, 2 mM KH
2PO
4, pH 7.4) per 100 ml of cell culture. The suspension was frozen and thawed several times to destroy the cells. Lysozyme was added to the cell suspension to reach a final concentration of 1 mg/ml, and this mixture was incubated on ice for 1 hr. After in- cubation, 10 ml of 0.2% Triton X-100 was added to the mix- ture, and the mixture was incubated at 4°C with rocking for 10 min. The insoluble debris was removed by cen- trifugation at 13,000x g for 1 hr at 4°C. The supernatant, which included the cell lysate, was collected in a fresh tube.
DTT was added to the tube to make a final concentration of 1 mM. The cell lysate was mixed with an appropriate amount of a 50% slurry of glutathione-agarose resin in PBS.
In order to combine the glutathione S-transferase (GST) fu- sion protein with the resin, the mixture was shaken gently overnight at 4°C. Then, it was loaded onto the poly-prep chromatography column (Bio-Rad, USA) under gravity flow.
Unbound proteins were washed away from the resin by add- ing 10 bed volumes of PBS to the pellet. Bound GST fusion protein was eluted from the resin using 1 bed volume of Glutathione Elution Buffer (10 mM reduced glutathione, 50 mM Tris-HCl, pH 8.0). All steps for the expression and puri- fication of the OsCPK11 were followed as described in Lee [31].
SDS-PAGE and sequence analysis
Discontinuous SDS-PAGE was performed according to Laemmli [29] using 10% acrylamide minislab gels, and the molecular weight of the OsCPK11-GST protein was determined.
Homology alignment and phylogenetic tree analyses of the
Fig. 1. Tissue-specific expression of OsCPK11. Total RNA was isolated from the flowers, young leaves, mature leaves, roots and aleurone layers of rice, and RT-PCR was performed. Primers for OsCPK11 were used in lanes 1, 3, 5, 7 and 9, and ones for rice actin were used for lanes 2, 4, 6, 8 and 10. M indicates DNA molecular weight markers.
A
B
C
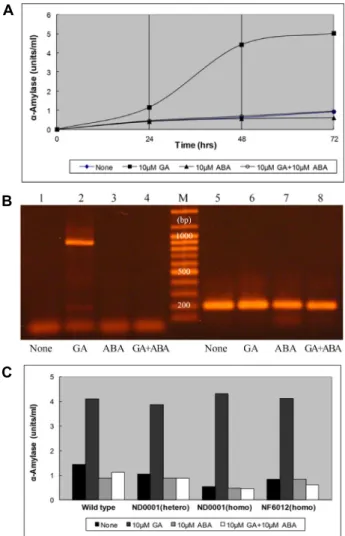
Fig. 2. (A) α-Amylase activity of GA- and/or ABA-treated rice aleurone layers. After each sample was incubated with starch solution at 37°C for 11 min, iodine solution was added, and sample absorbance was measured at 625 nm.
(B) OsCPK11 gene expression from rice aleurone layers treated with plant hormones. Total RNAs were extracted from rice aleurone layers treated for 72 hr with either no hormone (lanes 1 and 5), 10 μM GA (lanes 2 and 6), 10 μM ABA (lanes 3 and 7), or 10 μM GA + 10 μM ABA (lanes 4 and 8) as prepared in A. RT-PCR was per- formed afterwards. Primers for the OsCPK11 gene were used for lanes 1, 2, 3 and 4, and ones for the rice actin gene were used for lanes 5, 6, 7 and 8. M indicates DNA molecular weight markers. (C) GA-responsiveness of the aleurone layers from various oscpk11 mutants. Rice aleurone layers of wild type and three oscpk11 including a heterozygous mutant of ND0001, a homozygous mu- tant of ND0001 and a homozygous mutant of NF6012 were prepared as in A.
OsCPK11 with other CPKs were made using CLUSTAL methodology [49] and neighbor-joining phylogeny. CLUSTAL X (version 1.81) and Tree View (Win 32) were used, and the amino acids of CPKs were obtained from the NCBI gene bank (http://www.ncbi.nlm.nih.gov/).
Results
OsCPK11 gene expression in rice tissues and hormone-treated aleurone layers
Expression data is a good way to predict gene function [5]. Usually gene expression is detected by Northern blot or RT-PCR analysis [5, 46, 48, 51, 57]. For identification of the OsCPK11 transcript, RT-PCR was performed. In this study, the OsCPK11 gene expression was confirmed in flow- ers, young leaves and mature leaves, while the gene was not expressed in the roots or in the aleurone layers (Fig.
1).
To study OsCPK11 gene expression in hormone-treated rice aleurone layers, the effect of 10 μM GA and/or 10 μM ABA treatment on the layers was examined (Fig. 2A). The α-amylase activity of the 10 μM GA-treated aleurone layers was the highest at every time point from 24 hr to 72 hr, with activity at other time points almost similar to that of layers with no hormone treatment. Then, expression of the OsCPK11 gene in aleurone layers with or without hormone for 72 hr was tested (Fig. 2B). Presence of the OsCPK11 tran- script was confirmed in 10μM GA-treated aleurone layers.
In contrast, when layers were treated with 10 μM ABA only, 10 μM GA and 10 μM ABA together, or no hormone treat- ment, the OsCPK11 transcript was not present. OsCPK11
gene expression was induced by GA, while ABA prevented
its induction. Since we knew OsCPK11 was expressed in
GA-affected aleurone layers and would like to further look
at OsCPK11 participation in GA signaling, the GA-re-
Table 1. Caryopsis number and caryopsis weight of oscpk11 mu- tants
Genotype Caryopses/Panicle
(No.)
Caryopsis weight (mg) Wild type
ND0001 (heterozygous) ND0001 (homozygous) NF6012 (homozygous)
103.3±2.23 90.1±4.32* 77.4±6.16* 83.9±2.21*
25.9±0.15 24.3±0.18* 23.5±0.20* 23.2±0.23* Wild type and mutant line seedlings were harvested at 6 months after planting. The number of caryopses per panicle (average of 10 panicles in each individual) was counted and the caryopsis weight (average of 100 caryopses in each mutant line) were measured. Numbers are the mean ± SE and an aster- isk (*) indicates a statistical significance (p<0.01) compared to wild type.
A
B
Fig. 3. (A) Presence of white belly materials in the grains as a phenotypic feature. One hundred rice grains were husked and cut in half, and the number of grains with white belly materials was counted. (B) Presence of white belly materials in the grains of oscpk11 mutant lines. For each treatment group and wild type, 20 rice grains (Clockwise from top left: wild type, heterozygous ND0001, homozygous NF6012, homozygous ND0001) were husked and cut in half, and their cross sections were taken photographed using a digital camera.
sponsiveness of the aleurone layers from various oscpk11 mutants was examined (Fig. 2C). Aleurone layers of all test- ed mutant lines showed GA responsiveness of α-amylase activity.
Features of Oscpk11 mutants
Various mutant lines made by random insertion of the endogenous retrotransposon Tos17 into the rice genome are being used to study the rice genome [22]. To reveal the bio- logical function of OsCPK11, this study used two mutant lines (ND0001 and NF6012) in which Tos17 has been inserted into OsCPK11. In ND0001, Tos17 is reversely inserted into the sixth exon; in the case of NF6012, Tos17 is inserted into the third exon.
After genotyping the two mutant lines, the change in rice phenotypes - including plant height and caryopsis number and weight - by the Tos17 insertion was investigated. The heights of rice individuals of wild type and each mutant line were little different. However, there were statistically significant differences (p<0.01) in the number of caryopses per panicle and caryopsis weight (Table 1). Compared to wild type, the number of caryopses per panicle decreased to 13% in heterozygous mutants of the ND0001 line, 25%
in homozygous mutants of the ND0001 line and 19% in ho- mozygous mutants of the NF6012 line. Similarly, the car- yopsis weight from each mutant line also reduced to 6%(ND0001 - heterozygous), 9%(ND0001 - homozygous) and 10%(NF6012 - homozygous). In addition, the white belly materials in grains were confirmed to be 37% in hetero- zygous mutants of the ND0001 line, 49% in homozygous mutants of the ND0001 line and 55% in homozygous mu- tants of the NF6012 line (Fig. 3). Namely, all of the mutant
lines showed a distinct number of grains with white belly materials.
6cDNA cloning of OsCPK11 and partial charac- terization of OsCPK11 protein
In order to examine the function of OsCPK11 protein, it is necessary to reveal protein features. In this study, partial characterization of the OsCPK11 protein was performed.
In order to make the OsCPK11 protein, cDNA encoding
OsCPK11 was ligated into the pET-41a(+) vector, and DH5α
competent cells were transformed with the pET41a(+)-
OsCPK11 complex. Six transformants were identified out of
16 colonies, and the plasmid DNA of these colonies was
sequenced. Consequently, cDNAs inserted into plasmid
DNAs of three colonies completely matched the OsCPK11
cDNA sequence obtained from NCBI. Thereafter, the plas- mid of one colony was retransformed into BL21 competent cells, and production of OsCPK11 protein was induced by IPTG. The fusion protein expressed from the pET-41a(+) vec- tor contains a GST moiety. So, the OsCPK11-GST protein was purified by GST affinity chromatography and found to be a single polypeptide based on SDS-PAGE, consistent with the predicted molecular weight of the fusion protein (Fig.
4A). The molecular mass of OsCPK11-GST and GST were about 95.5 kDa and 35 kDa, respectively.
Analysis of the deduced amino acid sequence of OsCPK11 indicates that this protein has the structural characteristics of the CPKs (Fig. 4C). This kinase contains a variable N-ter- minal region, a catalytic domain with the 11-signature sub- domains characteristic of the Ser/Thr protein kinase family, an autoinhibitory domain and a calcium-binding domain with four EF-hands. Regions corresponding to (ⅰ) an ATP-binding site and (ⅱ) a Ser/Thr protein kinase active site within the catalytic domain are well conserved.
Compared to several other CPKs, OsCPK11 shares 78%, 70%, 67% and 53% identity in deduced amino acids with wheat TaCPK9, Phalaenopsis PaCDPK1, OsCDPK14 and OsCDPK2, respectively (Fig. 4C). The phylogenetic tree based on the mRNA sequence of each CPK showed that OsCPK11 was different from other known rice CPKs and most closely re- lated to TaCPK9 (Fig. 4B).
Discussion
As an important kinase family, it is essential to identify each individual isoform of CPKs and to understand that iso- form’s function in plant development [56]. In this study, the biological function of rice OsCPK11 gene was characterized, and its cDNA was cloned. Expression of the OsCPK11 gene in rice tissues and hormone-treated aleurone layers was confirmed. The features of oscpk11 mutants were also inves- tigated, and some of their phenotypic features were found.
When Asano et al. [5] and Wan et al. [51] examined the expression patterns of rice CPK genes, they could not detect the transcript of OsCPK11 or several other OsCPKs in the rice tissues analyzed. In contrast, we found OsCPK11 ex- pression in young leaves, mature leaves and flowers (Fig.
1). The reason why our results differed from previous stud- ies may be due to the treatments, the tissues that were used and the sensitivity of the protocols [51]. Anyway, tissue-spe- cific expression data suggest that OsCPK11 might be related
to signal transduction in cell differentiation and metabolism in the leaves and flowers of rice.
OsCPK11 transcripts were also confirmed in GA-treated aleurone layers (Fig. 2B). Aleurone layer cells of cereal grains are used as a model system for dissecting the molecular and cellular machinery of GA and ABA responses [8, 40].
Functions of the aleurone layer cells include the synthesis and release of hydrolytic enzymes into the starchy endo- sperm during germination, which is enhanced by GA. ABA reverses this GA effect in the aleurone layers [47]. Therefore, to identify whether OsCPK11 is involved in GA and ABA responses, aleurone layers from rice were used. First of all, α-amylase activity of hormone-treated aleurone layers showed the expected kinetics of GA and/or ABA responses (Fig. 2A), consistent with that described by Day [16].
Aleurone layers treated with plant hormones were used to characterize the expression profile of the OsCPK11. GA-re- sponsiveness of the aleurone layers from various oscpk11 mutants was very similar to that of wild type (Fig. 2C). Thus, OsCPK11 takes part in GA signaling of the aleurone layer, but it does not seem to be necessary in this process.
One way to investigate the putative function of an un- known gene is to mutate the gene and compare the mutant phenotype with that of the wild type [32]. Here, the pheno- types of oscpk11 mutants were examined. Neither of the two mutant lines (ND0001, NF6012) showed serious abnormal phenotypes during growth. For instance, plant height of the mutants was indistinguishable from that of wild type plants.
However, defects in the OsCPK11 protein slightly impaired caryopsis development, considering the number of caryopsis in each panicle and caryopsis weight (Table 1). These results suggest that OsCPK11 is not a critical factor in these bio- logical events during growth; nevertheless, it may play some role in mediating seed development. In addition, many grains of the mutant lines had white belly materials in the endosperm (Fig. 3). To stabilize the metabolic perturbation caused by the absence of OsCPK11, it seems that the white belly endosperm phenotype is made by a compensatory pathway [26]. Further study is required to characterize their opacity. The phenotypes of opaque-grain mutants include dull, floury, glutinous, shrunken, white belly and white core endosperms [38, 45]. These aberrations can provide valuable information for elucidating the metabolic processes related to nutrition storage during grain filling [38].
Since the cDNA clone of CPK was first isolated from
Arabidopsis, various CPK genes have been cloned from plant
A B
C
Fig. 4. (A) Expression of OsCPK11-GST and GST protein from BL21 cells. OsCPK11-GST and GST protein were visualized with Coomassie blue staining after SDS-PAGE. Numbers in the left indicate the molecular weight of marker proteins. (B) Phylogenetic tree based on the mRNA sequences of OsCPK11 and other CPKs. Numbers on the branches are bootstrap proba- bilities determined from 1,000 samplings. Accession numbers are as follows: OsCDPK5 (AF194414); OsCDPK2 (X81394);
OsCDPK14 (AY144497); OsCPK11 (AC084296); TaCPK9 (EU181184); IiCPK2 (DQ458915); PaCDPK1 (EF555574); NtCDPK2 (AJ344154); NtCDPK3 (AJ344155); RiCDPK1 (AB0- 051808); SPK (D13436); OsCDPK13 (AB078634); OsCDPK7 (AB042550);
ScCPK (EU305742) and CaCDPK3 (AY295081). (C) Alignment of the predicted amino acid sequence of the OsCPK11 with other CPKs. Catalytic (I~XI), autoinhibitory and calcium-binding (EF1~EF4) domains are underlined above the amino acid sequences and signature residues of the kinase in catalytic domain (i and ii) are underlined below the amino acid sequences.
Accession numbers of the sequences were as follows: OsCPK11 (AAT75264); TaCPK9 (ABY59011); PaCDPK1 (ABU45516);
OsCDPK14 (AAN41657) and OsCDPK2 (CAA57157).
species such as carrot, maize, rice and tobacco [20, 55, 56].
In this study, the cDNA of OsCPK11 was cloned, and an OsCPK11-GST fusion protein of about 95.5kD was obtained (Fig. 4A). Since OsCPK11 has the typical structural features of the CPK family (Fig. 4C), purified protein could be used for the kinase assay. Moreover, the deduced amino acid sequence of the OsCPK11 showed strong homology (78%) to TaCPK9, a well-known CPK, and a moderate homology (52%) to SPK. Therefore, the study of OsCPK11 can provide a mechanism to decode calcium signals like all other CPKs.
There is a possibility that OsCPK11 could be localized in the cytoplasm, since OsCPK11 doesn't have a myristoylation or a palmitoylation motif in its N-terminal portion. Both myristoylation and palmitoylation appear to be important factors for the membrane binding of CPKs in plant species [17, 34, 36]. For instance, the OsCDPK2 that contains the myristoylation motif MGSCCS and the palmitoylation motif MGSCCS at its N-terminus is myristoylated and palmitoylated and is targeted to the membrane fraction [36].
In summary, the OsCPK11 gene was expressed in both the leaves and flowers of rice and in GA-treated aleurone layers, and it had an effect on caryopsis development.
OsCPK11, which contains the typical structural features of the CPK family, was cloned from rice. These results could be helpful in understanding the role of CPKs in Ca
2+-medi- ated signaling in plants.
Acknowledgement
This research was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education [NRF-2011- 0025592] to S.-H. Kim.
References
1. Abo-El-Saad, M. and Wu, R. 1995. A rice membrane cal- cium-dependent protein kinase is induced by gibberellin in rice leaf sheath. Plant Mol. Biol. 55, 541-552.
2. Anil, V. S. and Rao, S. K. 2000. Calcium-mediated signalling during sandalwood somatic embryogenesis. Role for exoge- nous calcium as second messenger. Plant Physiol. 123, 1301- 1311.
3. Anil, V. S., Harmon, A. C. and Rao, S. K. 2000. Spatio-tem- poral accumulation and activity of calcium-dependent pro- tein kinases during embryogenesis, seed development, and germination in sandalwood. Plant Physiol. 122, 1035-1043.
4. Asano, T., Kunieda, N., Omura, Y., Ibe, H., Kawasaki, T., Takano, M., Sato, M., Furuhashi, H., Mujin, T., Takaiwa, F.,
Wu, C., Tada, Y., Satozawa, T., Sakamoto, M. and Shimada, H. 2002. Rice SPK, a calmodulin-like domain protein kinase, is required for storage product accumulation during seed development: phosphorylation of sucrose synthase is a pos- sible factor. Plant Cell 14, 619-628.
5. Asano, T., Tanaka, N., Yang, G., Hayash, N. and Komatsu, S. 2005. Genome-wide identification of the rice cal- cium-dependent protein kinase and its closely related kinase gene families: Comprehensive analysis of the CDPKs gene family in rice. Plant Cell Physiol. 46, 356-366.
6. Asano, T., Hakata, M., Nakamura, H., Aoki, N., Komatsu, S., Ichikawa, H., Hirochika, H. and Ohsugi, R. 2011. Functio- nal characterisation of OsCPK21, a calcium-dependent pro- tein kinase that confers salt tolerance in rice. Plant Mol. Biol.
75, 179-191.
7. Berridge, M. J., Lipp, P. and Bootman, M. D. 2000. The ver- satility and universality of calcium signalling. Nat. Rev. Mol.
Cell Biol. 1, 11-21.
8. Bethke, P. C., Schuurink, R. and Jones, R. L. 1997. Hormonal signaling in cereal aleurone. J. Exp. Bot. 48, 1337-1356.
9. Bush, D. S. and Jones, R. L. 1988. Cytoplasmic calcium and α-amylase secretion from barley aleurone protoplasts. Eur.
J. Cell Biol. 46, 466-469.
10. Bush, D. S. 1995. Calcium regulation in plant cells and its role in signaling. Annu. Rev. Plant Physiol. Plant Mol. Biol.
46, 95-122.
11. Cheng, S. H., Willmann, M. R., Chen, H. C. and Sheen, J.
2002. Calcium signaling through protein kinases. The Arabidopsis calcium-dependent protein kinase gene family.
Plant Physiol. 129, 469-485.
12. Cheong, Y. H., Kim, K. N., Pandey, G. K., Gupta, R., Granta, J. J. and Luan, S. 2003. CBL1, a calcium sensor that differ- entially regulates salt, drought, and cold responses in Arabidopsis. Plant Cell 15, 1833-1845.
13. Chin, D. and Means, A. R. 2000. Calmodulin: a prototypical calcium sensor. Trends Cell Biol. 10, 322-328.
14. Christodoulou, J., Malmendal, A., Harper, J. F. and Chazin, W. J. 2004. Evidence for differing roles for each lobe of the calmodulin-like domain in a calcium-dependent protein kinase. J. Biol. Chem. 279, 29092-29100.
15. Cohen, S. N., Chang, A. C. Y. and Hsu, L. 1972. Nonchromo- somal antibiotic resistance in bacteria: Genetic transfor- mation of Escherichia coli by R-factor DNA. Proc. Nat. Acad.
Sci. USA 69, 2110-2114.
16. Day, P. M. 2008. Phosphatidylinositol 3-kinase is a positive regulator of gibberellin signaling. Ph.D. thesis, Penn State University, USA.
17. Ellard-lvey, M., Hopkins, P. B., White, T. J. and Lomax, T.
L. 1999. Cloning, expression and N-terminal myristoylation of CpCPK, a calcium-dependent protein kinase from zucchi- ni (Curbita pepo L). Plant Mol. Biol. 39, 199-208.
18. Evans, N. H., McAinsh, M. R. and Hetherington, A. M. 2001.
Calcium oscillations in higher plants. Curr. Opin. Plant Biol.
4, 415-420.
19. Harmon, A. C., Gribskov, M. and Harper, J. F. 2000.
CDPKs-a kinase for every Ca2+ signal? Trends Plant Sci. 5,
154-159.
20. Harper, J. F., Sussman, M. R., Schaller, G. E., Putnam-Evans, C., Charbonneau, H. and Harmon, A. 1991. A calcium-de- pendent protein kinase with a regulatory domain similar to calmodulin. Science 252, 951-954.
21. Harper, J. F. 2001. Dissecting calcium oscillators in plant cells. Trends Plant Sci. 6, 395-397.
22. Hirochika, H., Sugimoto, K., Otsuki, Y., Tsugawa, H. and Kanada, M. 1996. Retrotransposons of rice involved in muta- tions induced by tissue culture. Proc. Natl. Acad. Sci. USA 93, 7783-7788.
23. Hernandez, S., Hardin, S., Clouse, S., Kieber, J. and Huber, S. 2004. Identification of a new motif for CDPK phosphor- ylation in vitro that suggest ACC synthase may be a CDPK substrate. Arch. Biochem. Biophys. 428, 81-91.
24. Hrabak, E. M., Chan, C. W., Gribskov, M., Harper, J. F., Choi, J. H., Halford, N., Kudla, J., Luan, S., Nimmo, H. G., Sussman, M. R., Thomas, M., Walker-Simmons, K., Zhu, J.
K. and Harmon, A. C. 2003. The Arabidopsis CDPK-SnRK superfamily of protein kinases. Plant Physiol. 132, 666-680.
25. International Rice Genome Sequencing Project. 2005. The map-based sequence of the rice genome. Nature 436, 793-800.
26. Kang, H., Park, S., Matsuoka, M. and An, G. 2005. White- core endosperm floury endosperm-4 in rice is generated by knockout mutations in the C4-type pyruvate orthophosphate dikinase gene (OsPPKB). Plant J. 42, 901-911.
27. Knight, H. and Knight, M. R. 2001. Abiotic stress signalling pathways: specificity and cross-talk. Trends Plant Sci. 6, 262- 267.
28. Kolukisaoglu, U., Weinl, S., Blazevic, D., Batistic, O. and Kudla, J. 2004. Calcium sensors and their interacting protein kinases: genomics of the Arabidopsis and rice CBL-CIPK signalling networks. Plant Physiol. 134, 43-58.
29. Laemmli, U. K. 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 277, 680-685.
30. Lehninger, A. L., Nelson, D. L. and Cox, M. M. 1993.
Principles of biochemistry, pp. 772, 2nd ed., Worth Publishers, NY, USA.
31. Lee, S. H. 2009. Functional characterization of OsCPK11, a calcium-dependent protein kinase gene from rice and its cDNA cloning. Master’s thesis. Korea National University of Education, Chungbuk, Korea.
32. Lee, J. E. 2008. Functional analysis of OsCPK11, a calcium- dependent protein kinase gene from rice. Master’s thesis.
Korea National University of Education, Chungbuk, Korea.
33. Li, J., Lee, Y. R. and Assmann S. M. 1998. Guard cells pos- sess a calcium-dependent protein kinase that phosphor- ylates the KAT1 potassium channel. Plant Physiol. 116, 785- 795.
34. Lu, S. X. and Hrabak, E. M. 2002. An Arabidopsis cal- cium-dependent protein kinase is associated with the endo- plasmic reticulum. Plant Physiol. 128, 1008-1021.
35. Maria, K. and Grazyna, M. 2007. Structure and functions of plant calcium-dependent protein kinases. Acta Biochim.
Polonica 54, 219-233.
36. Martin, M. L. and Busconi, L. 2000. Membrane localization of a rice calcium-dependent protein kinase (CDPK) is medi- ated by myristoylation and palmitoylation. Plant J. 24, 429- 435.
37. Nelson, D. L. and Cox, M. M. 2013. Lehninger principles of biochemistry, 6th ed., Freeman and Company, NY, USA.
38. Nelson, L. E. and Pan, D. 1995. Starch synthesis in maize endosperms. Annu. Rev. Plant Physiol. Plant Mol. Biol. 46, 475-496.
39. Racies, M., Gargantini, P. R., Chinchilla, D., Crespi, M., Tellez-Inon, M. T. and Ulloa, R. M. 2003. Regulation of CDPK isoforms during tuber development. Plant Mol. Biol.
52, 1011-1024.
40. Ritchie, S. and Gilroy, S. 1998. Gibberellins: regulating genes and germination. New Phytol. 140, 363-383.
41. Saijo, Y., Hata, S., Kyozuka, J., Shimamoto, K. and Izuki, K. 2000. Over-expression of a single Ca2+-dependent protein kinase confers both cold and salt/drought tolerance on rice plants. Plant J. 23, 319-327.
42. Sanders, D., Brownlee, C. and Harper, J. F. 1999. Communi- cating with calcium. Plant Cell 11, 691-706.
43. Sanders, D., Pelloux, J., Brownlee, C. and Harper, J. F. 2002.
Calcium at the crossroads of signalling. Plant Cell 14 (Suppl), S401-S417.
44. Sasaki, T. and Burr, B. 2000. International Rice Genome Sequencing Project: the effort to completely sequence the rice genome. Curr. Opin. Plant Biol. 3, 138-141.
45. Satoh, H. and Omura, T. 1981. New endosperm mutations induced by chemical mutagen in rice, Oryza sativa L. Jap.
J. Breed. 31, 316-326.
46. Shen, W., Tao, G. Q., Li, D. C., Zhu, X. G., Bai, X. and Cai, B. 2008. Inhibition of pancreatic carcinoma cell growth in vitro by DPK4 gene transfection. World J. Gastroenterol. 14, 6254-6260.
47. Taiz, L., Zeiger, E., Moller, I. M. and Murphy, A. 2015. Plant physiology and development, 6th ed., Sinauer Associates, MA, USA.
48. Takada, A., Berezikov, E., Yamashita, Y., Laqos-Quintana, M., Kloosterman, W. P., Enomoto, M., Hatanaka, H., Fujiwara, S., Watanabe, H., Soda, M., Choi, Y. L., Plasterk, R. H., Cuppen, E. and Mano, H. 2006. Mouse microRNA profiles determined with a new and sensitive cloning method. Nucl. Acids Res. 34, e115.
49. Thompson, J. D., Higgins, D. G. and Gibson, T. J. 1994.
CLUSTAL W: improving the sensitivity of progressive mul- tiple sequence alignment through sequence weighting, posi- tions-specific gap penalties and weight matrix choice. Nucl.
Acids Res. 22, 4673-4680.
50. Trewavas, A. J. and Malho, R. 1998. Ca2+ signalling in plant cells: the big network! Curr. Opin. Plant Biol. 1, 428-433.
51. Wan, B., Lin, Y. and Mou, T. 2007 Expression of rice Ca2+-de- pendent protein kinases genes under different environ- mental stresses. FEBS Lett. 581, 1179-1189.
52. Yang, G., Shen, S., Yang, S. and Komatsu, S. 2003.
OsCDPK13, a calcium-dependent protein kinase gene from rice, is induced in response to cold and gibberellin. Plant
초록:벼의 칼슘-의존성 단백질 카이네즈 유전자인 OsCPK11 의 기능적 분석
이수희
1․이정은
2․필립 데이
3․사이몬 길로이
4․김성하
5*
(1이월중학교, 2거제중학교, 3미국 펜실베니아 주립대학교(알투나) 수학·자연과학과, 4미국 위스콘신 대학교(메디슨)
식물학과, 5한국교원대 생물교육과)
Calcium-dependent protein kinases (CDPKs)는 칼슘 이온을 매개로 한 신호전달 경로에서 중요한 역할을 한 다. 벼(Oryza sativa)에는 29개의 CDPKs가 확인되었지만 그들의 기능은 완벽히 밝혀지지 않았다. 이 연구는 OsCPK11 유전자에 초점을 맞춰 그것의 기능적인 특징을 조사하였다. 벼의 어린 잎, 성장한 잎, 꽃에서 OsCPK11 유전자의 조직-특이적 발현이 확인되었고, 지베렐린이 처리된 벼의 호분층에서도 이 유전자의 발현을 확인할 수 있었다. Tos-17이 삽입된 oscpk11의 표현형에서 돌연변이체 각각의 키는 야생형과 구분되지 않았지만, 영과의 수나 무게는 통계적으로 유의미한 차이가 있었다. 덧붙여 많은 돌연변이체 낟알의 배젖에서 white belly materials이 확인되었다. OsCPK11의 cDNA가 cloning되었고, 약 60.5 kD인 OsCPK11 단백질이 GST affinity chromatography 와 SDS-PAGE에 의해 얻어졌다. 아미노산 서열 분석을 통해 OsCPK11이 전형적인 CDPKs의 구조적 특징을 가짐 을 알 수 있었다. 이 결과는 OsCPK11유전자의 기능과 식물에서 칼슘 이온을 매개로 한 신호전달 경로에 CDPK 역할의 유용한 정보를 제공해줄 것이다.
Physiol. Biochem. 41, 369-374.
53. Ye, S., Wang, L., Xie, W., Wan, B., Li, X. and Lin, Y. 2009.
Expression profile of calcium-dependent protein kinase (CDPKs) genes during the whole lifespan and under phyto- hormone treatment conditions in rice (Oryza sativa L. ssp.
indica). Plant Mol. Biol. 70, 311-325 .
54. Zhang, X. and Chollet, R. 1997. Seryl-phosphorylation of soybean nodule sucrose synthase (nodulin-100) by a Ca2+-dependent protein kinase. FEBS Lett. 410, 126-130.
55. Zhang, M., Liang, S. and Lu, Y. T. 2005. Cloning and functional characterization of NtCPK4, a new tobacco cal-
cium-dependent protein kinase. Biochim. Biophys. Acta 1729, 174-185.
56. Zhang, T., Wang, Q., Chen, X., Tian, C., Wang, X., Xing, T., Li, Y. and Wang, Y. 2005. Cloning and biochemical prop- erties of CDPK gene OsCDPK14 from rice. J. Plant Physiol.
162, 1149-1159.
57. Zhijun, X., Shulan, Z. and Xhuo, Z. 2007 Expression and significance of the protein and mRNA of metastasis sup- pressor gene ME491/CD63 and integrin alpha5 in ovarian cancer tissues. Eur. J. Gynaecol. Oncol. 28, 179-183.