801
서 론동물의에너지수지에관한연구는에너지흐름을이해하고동 물생태계의기능을파악하는데있어유용한수단이된다
.
생태 계는복잡한먹이사슬또는먹이그물과같은영양적구조로집 단간의에너지흐름의양에밀접하게관련되어있으며,
이러한 복잡한먹이망속에서그물질의전환에따른에너지흐름을 파악하기위해서에너지수지를분석하는것이매우중요하다(Klekowski and Duncan, 1975).
어느한생물의개체에유입되 는모든에너지는열로써상실하거나,
다른영양단계로전이된 다.
생물학적물질계는물질전환에따른에너지흐름에의해서 체계화되어있다.
그리고먹이사슬로또는먹이그물과같은영 양적구조는집단의에너지흐름의양에밀접하게관련된다.
그 러므로에너지흐름의파악은자연집단의에너지수지를분석하 는것이필요하지만,
우선개체의생활사에따른에너지수지를 정확하게분석하는것이매우중요하다(Park, 1989).
갑각류의에너지수지에관해서는자연과실험실내집단을대 상으로많은연구가진행되어있으며
(Clutter and Theilacker, 1971; Levine and Sulkin, 1979; Emmerson, 1984; Harms, 1987; Kim and Chin, 1991),
성장,
산소소비,
질소배설및섭취 에대한에너지함량을파악하여에너지수지방식을통해서그 영양적체계를구성해볼수있다(Park, 1989).
동물의에너지 수지에관한연구는다른영양단계간의에너지전이유형을이 해하는총체적지표가되며,
그동물의생산성과생태적효율을 예측하는자료로서유용하다(Vernberg and Vernverg, 1981).
가시배새우
, Lebbeus groenlandicus
는십각목(Order Decap- oda)
꼬마새우과(Family Hippolytidae)
에속하는종으로꼬마 새우과중에서도거대종이며,
특히한국산꼬마새우과중에서 는가장크다.
몸이짧고굵으며,
두흉갑이짧고거친털로덮여 있다.
이종은울릉도근해에서통발로주로어획하는데,
수심200-400 m
에주로서식한다.
한해성종으로서아한대해역에분포하고있다
.
우리나라동해,
그린란드,
미국동북부,
캐나다Article history;
Received 8 May 2013; Revised 27 August 2013; Accepted 29 October 2013
*Corresponding author: Tel: +82. 33. 640. 2342 Fax: +82. 33. 640. 2340 E-mail address: [email protected]
Kor J Fish Aquat Sci 46(6) 801-806, December 2013 http://dx.doi.org/10.5657/KFAS.2013.0801 pISSN:0374-8111, eISSN:2287-8815
ⓒ The Korean Society of Fishereis and Aquatic Science. All rights reserved
가시배새우(Lebbeus groenlandicus)의 유생 단계 에너지수지
김원기·권오남
1·박기영
2*양양군청 해양수산과, 1강릉원주대학교 해양생물연구교육센터, *2강릉원주대학교 해양자원육성학과
Energy Budget of Spiny Lebbeid Shrimp Lebbeus groenlandicus Larvae
Won Gi Kim, O-Nam Kwon1 and Kie-Young Park2*
Marine and Fisheries Division, Yangyang country Office, Yangyang-gun 215-701, Korea
1
Marine Biology Center for Research and Education, Gangneung-Wonju National University, Gangneung 210-853, Korea
2
Department of Marine Bioscience, Gangneung-Wonju National University, Gangneung 210-702, Korea
Spiny lebbeid shrimp Lebbeus groenlandicus larvae were reared in the laboratory to estimate the energy budget from the zoeal to the post-larval stage. Energy expended by larvae on growth and respiration was determined from values for feeding, growth, molting, and metabolism. We calculated that 16.22 J were used for growth throughout all larval stages. Energy loss during molt- ing was estimated as 1.03 J, and energy used for respiration was estimated as 1.31 J. Energy taken in by feeding was estimated as 77.16 J, while the sum of energies expended in excretion and egestion was 58.61 J. Larvae were estimated to assimilate 24.6%
of ingested food as energy and to use ~85% of the assimilated energy for somatic growth. Gross growth efficiency (K
1) and net growth efficiency (K
2) were shown to be ~22% and 93%, respectively. Maintenance costs of respiration were estimated as ~9.7%
of assimilated energy.
Key words: Lebbeus groenlandicus, Metabolism, Energy budget, Spiny lebbeid shrimp, Assimilation efficiency, Growth efficiency
북극해연해
,
오호츠크해,
일본홋카이도등전세계적으로널리 분포하고있다(NFRDI, 2007).
따라서본연구는가시배새우유생을대상으로성장
,
탈피,
호 흡,
섭취및에너지함량을측정하여유생발달동안에너지수 지를분석하고섭취와성장에따른유생의에너지흐름의경향 을파악함으로써개체및개체군에대한생리적기초자료를얻 는데그목적이있다.
재료 및 방법
실험동물 및 유생사육
실험에 사용한 가시배새우 유생은 평균 체장
117.22 mm,
중량38.65 g
의모하로부터얻었다.
내장을제거한민들조개, Macridiscus melanaegis
를부화직전인암적색의알을달고있 는모하들에게먹이로공급하였다.
이들모하들을별도용기에 수용하여부화당일혹은부화후1
일령의부유성zoea I
기유생 을수집하여8±0.3℃
에서실험에사용하였다.
성장 및 에너지함량
초기유생의개체별성장을측정하기위하여수온
8±0.3℃,
250 mL
용기(
사육수, 200 mL)
에수용한후부화직후유생을1
마리씩수용하여Artemia nauplii (
이하Artemia)
를약10
개 체/mL
의농도로공급하면서매일아침탈피체를수거하여크 기를측정하였다.
성장측정은유생의손상을방지하기 위해 실험중인유생의탈피체를수거하여 측정하고,
각유생단계 의탈피체의에너지함량및건조중량은각 발달단계별로대형사육
(1 ton)
수조에서무작위추출하여측정하였다.
실험유생의체장은마지막복부체절의말단에서두흉갑의 앞쪽끝 까지의직선거리를광학현미경
(OLYMPUS IX70)
을이용하여0.01 mm
단위로측정하였다.
또한초기유생단계별유생및탈 피체에에너지함량은자동폭발열량계(Van calorimeter model PARR1315, USA)
로측정하여‘J'
로표시하였다.
산소소비
유생의산소소비량은
250 mL
산소병에가시배새우부화직후 유생을5
마리씩수용한후,
실험전후각용존산소를DO meter (YSI 556MPS, USA)
로측정하였고,
실험전후의용존산소차 로써정량하였다.
각실험은5
회반복한평균치로개체당단위 시간당산소소비량을μLO
2/ind/day
로표시하였다.
산소소비량 은대사열생산에의해소비된에너지로전환시키기위하여Gn-
aiger (1983)
가이용한산소분자의에너지전화계수를이용하였다
(1 mgO
2= 3.38 cal).
대사에쓰인에너지는4.71×10
-3cal/
μLO
2의변환계수를사용하여계산하였다.
개체의산소소비량 은환경수의산소분압에영향을받기때문에(Ikeda, 1974),
산 소소비측정이끝날때의실험용기내용존산소량이실험시작시산소량의
60%
이하로떨어지지않게조절하였다.
먹이 섭취
유생의섭취량측정시에는
250 mL
의용기에가시배새우유생을
1
마리씩수용한후일정량의계수된Artemia nauplius
를 투여하여24
시간동안의섭취량을측정하였다.
그리고1
개체당 일간Artemia nauplii
섭취량은no./ind./day
으로나타내었다.
섭취에너지는1
마리가1
일섭취한Artemia nauplii
의개체당 중량(Dry weight 1.97 μg/nauplius, 4,619 cal/mg) (Emmerson, 1980)
과Artemia nauplii 1
마리의에너지함량(37.9 mJ) (Park, 1989)
을기준으로계산하였다.
에너지 수지
각유생단계및전체유생발생에대한에너지수지는
Standard IBP
방정식(Klekowski and Duncan, 1975)
으로나타내었다.
C = Pg + Pe + R + E ………①
①
식에C
는섭취에너지, Pg
는유생단계별체성장에쓰인에 너지, Pe
는탈피에쓰인에너지, R
은호흡에쓰인대사에너지, E
는대사산물인뇨배설에너지그리고대사되지않고배출되는 분배출에너지,
섭취에너지(C)
와동화에너지(A = Pg + Pe + R)
간의차로서나타내었다(Dawirs, 1983).
생태효율
수산동물의동화효율측정은먹이섭취량과분배출량을측정 하여먹이섭취량에대한동화량을백분율로하여동화효율을 구하였다
.
가시배새우의유생에대한섭취,
성장,
탈피및호흡 을측정한자료를사용하여다음효율들을계산하였다.
동화효율
: (Pg + Pe + R) / C × 100 ………②
②
식은섭취량중동화된양의백분율을나타낸다.
총성장효율
(K1): (Pg + Pe) / C × 100 ………③
③
식은섭취량중성장으로전환된양의백분율을나타낸다.
순성장효율
(K2): (Pg + Pe) / (Pg + Pe + R) × 100 ………④
④
식은동화량중성장으로전환된양의백분율을나타낸다.
결 과
성장 및 에너지 함량
가시배새우의 부화후 경과시간에 따른 유생
(zoea I, II
및megalopa)
의체장,
건조중량및유생과탈피체의에너지함량은
Table 1
에나타내었다. Zoea I
기에서megalopa
기까지의체 장성장은지수적인증가를보였다(Fig. 1).
성장에사용된에너지즉
,
생체에너지함량은J/ind.
로산출 하였다. ZoeaⅠ
기에서megalopa
기까지성장하는동안mega-
lopa
기의생체에너지는16.21 J
의에너지를체성장에사용하 였다.
성장에수반되어탈피체로 손실된에너지는탈피체에너지함량을
J/molting body
로 계산하였는데, zoeaⅠ
기부터megalopa
기까지유생1
마리가사용한탈피에너지는1.03 J
이었다
(Table 1).
체장성장과건조중량은경과일수에대해서각각
y=6.088e
0.116x(R
2=0.973)
와y=0.165e
0.468x(R
2=0.893)
로나 타났다(Fig. 1
과2).
산소소비
가시배새우유생발달단계에따라산소소비량은
zoeaⅠ
기의1.449 μLO
2/ind./day
에서megalopa
기의4.731 μLO
2/ind,/day
로성장에따라 증가하였다(Table 2).
일간대사에너지(J/ind./
day)
를 사용하여 유생단계별경과시간에 따른 대사에너지를 계산하면zoeaⅠ
기기간동안에는0.268 J, zoeaⅡ
기는0.601 J, megalopa
기기간동안에는0.982 J
을소비하여유생기간동 안소비된총산소소비량은1.851 J
이었다.
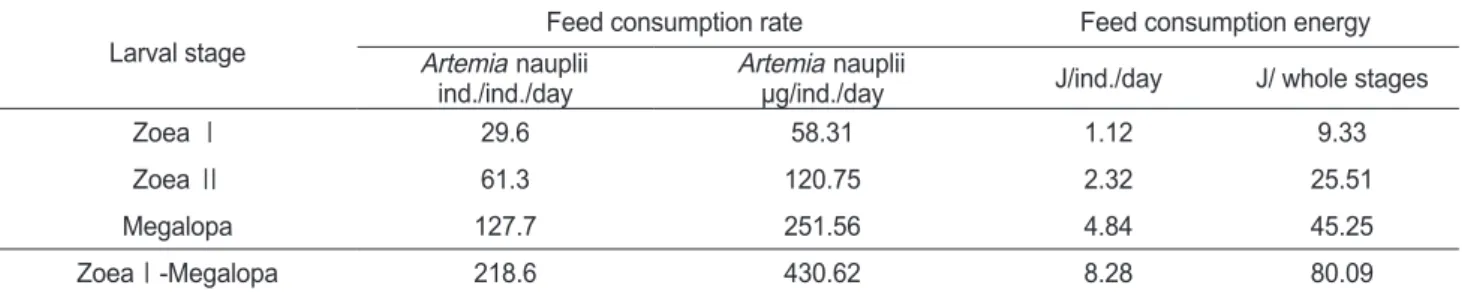
섭취
가시배새우의
zoea
유생들과megalopa
유생들에 의해 섭 취된일간섭취량과일간섭취에너지를Table 3
에나타내었다. ZoeaⅠ
기의29.6 Artemia/ind./day
에서megalopa
기의127.7
Artemia/ind./day
까지유생발달의진행과함께먹이섭취량은급격히증가하였다
. ZoeaⅠ
기는1.12 J/ind./day
의에너지를섭 취하는것으로조사되었으며,
유생이성장함에따라섭취에너 지는증가하여megalopa
기는4.84 J
의에너지를섭취하였다.
각유생단계별경과시간에따른각단계의섭취에너지를바탕 으로전유생기동안섭취한총섭취에너지는80.09 J
이었다. 에너지수지
성장에따른전유생기동안의유생의성장에너지함량
(Pg)
및 탈피체의에너지함량(Pe)
은Table 1
에서제시한결과를사용하 였으며,
호흡대사에너지(R)
는Table 2
의자료로부터계산하였 다.
그리고섭취에너지(C)
는Table 3
의일간섭취량과일간섭 취에너지로부터계산하였다.
뇨배설과분배출에너지(E)
는섭 취에너지(C)
와동화에너지(A)
의차이로서계산하였다.
전유 생기동안의누적에너지수지를Table 4
에나타내었다.
가시배 새우유생의섭취,
탈피체,
호흡및배설에너지는성장과함께 Fig. 1. Relationship between intermolt period and body length ofLebbeus groenlandicus for the different larval stages.
Fig. 2. Relationship between intermolt period and dry weight of Lebbeus groenlandicus for the different larval stages.
Table 1. The intermolt periods, body length, body weight and energy contents of Lebbeus groenlandicus for the different larval stages
Larval
stage Intermolt periods (day)
Body length
(mm)
weight Dry
(mg) J/ind. J/molt
ZoeaⅠ 0 6.92 0.29 16.21 1.03
ZoeaⅡ 8.32 7.52 0.35 16.21 1.03
Megalopa 10.98 8.74 0.74 16.21 1.03
Table 2. Individual oxygen consumption and respiratory energy expenditure of Lebbeus groenlandicus for the different larval stages
Larval stage O2 consumption Respiratory energy expenditure μLO2/ind./day J/ind./day J/whole
stages
Zoea Ⅰ 1.449 0.032 0.268
Zoea Ⅱ 2.467 0.055 0.601
Megalopa 4.731 0.105 0.982
ZoeaⅠ-
Megalopa 8.647 0.192 1.851
1 mgO2= 3.38 cal (Gnaiger, 1983) 1 cal = 4.71×10-3 J
y = 6.088e
0.116xR
2= 0.973 10
9 8 7 6
5
0 8.32 19.30
Body length (mm)
Intermolt periods (day)
Zoea I
Zoea II
Megalopa
y = 0.165e
0.468xR
2= 0.893 1.0
0.9 0.7 0.5 0.3 0.1 0.8 0.6 0.4 0.2
0.0
0 8.32 19.30
Body length (mm)
Intermolt periods (day)
Zoea I
Zoea II
Megalopa y = 6.088e
0.116xR
2= 0.973 10
9 8 7 6
5
0 8.32 19.30
Body length (mm)
Intermolt periods (day)
Zoea I
Zoea II
Megalopa
y = 0.165e
0.468xR
2= 0.893 1.0
0.9 0.7 0.5 0.3 0.1 0.8 0.6 0.4 0.2
0.0
0 8.32 19.30
Body length (mm)
Intermolt periods (day)
Zoea I
Zoea II
Megalopa
증가하였다
. ZoeaⅠ
기에서megalopa
기까지의총누적섭취에 너지는80.09 J
이었으며,
이것은성장에16.21 J,
탈피체손실 에1.03 J
그리고호흡에1.85 J
의에너지를사용하며뇨와분으로
61.00 J
의에너지가배설과배출이이루어지는것으로조사되었다
. 생태효율
가시배새우유생의섭취
,
성장,
탈피및호흡을측정한자료 로부터생태효율을Table 5
에나타내었다.
유생의동화효율은19.09%
이었으며,
총성장효율(K
1)
은21.53%,
순성장효율(K
2)
은
90.30%
로나타났다.
각에너지수지구성요소들의섭취에너지에대한백분율은성장이
20.24%,
탈피체가1.29%,
대사에 너지가2.31%
그리고뇨와분으로76.16%
를각각차지하였다.
ZoeaⅠ
기부터megalopa
기까지의총동화에너지는전체섭취에너지의
23.84%
인19.09 J
이었으며,
각에너지수지구성요 소들이동화에너지에대한백분율은성장이84.91%,
탈피체가5.40%
그리고대사에너지가9.70%
를각각차지하였다.
또한zoeaⅠ
기에서megalopa
기까지의 총생산에너지는총동화에Table 3. Consumption rate and consumption energy of Lebbeus groenlandicus for the different larval stages
Larval stage Feed consumption rate Feed consumption energy
Artemia nauplii
ind./ind./day Artemia nauplii
μg/ind./day J/ind./day J/ whole stages
Zoea Ⅰ 29.6 58.31 1.12 9.33
Zoea Ⅱ 61.3 120.75 2.32 25.51
Megalopa 127.7 251.56 4.84 45.25
ZoeaⅠ-Megalopa 218.6 430.62 8.28 80.09
37.9 mJ/Artemia nauplius (Park, 1989)
Table 4. Energy budgets of Lebbeus groenlandicus for the different larval stages
Larval stage C Pg Pe R E
Energy budget per hour (mJ)
Zoea Ⅰ 46.72 9.77 0.60 1.34 35.02
Zoea Ⅱ 96.80 18.52 1.18 2.28 74.83
Megalopa 201.65 41.80 2.67 4.38 152.80
Energy budget for the each larval and total development of zoeaⅠ to megalopa (J)
Zoea Ⅰ 9.33 1.95 0.12 0.27 6.99
Zoea Ⅱ 25.51 4.88 0.31 0.60 19.72
Megalopa 45.25 9.38 0.60 0.98 34.29
ZoeaⅠ~ Megalopa 80.09 16.21 1.03 1.85 61.00
Table 5. Accumulative energy budget of Lebbeus groenlandicus larvae
Symbol Total energy (J) %P %A %C
Molting Pe 1.03 5.97 5.40 1.29
Growth Pg 16.21 94.03 84.91 20.24
Total production P 17.24 100.00 90.30 21.53
Respiration R 1.31 9.70 2.31
Total assimilation A 19.09 100.00 23.84
Excretion & Egestion E 61.00 76.16
Consumption C 80.09 100.00
%P : Total production percentage, %A : Total assimilation percentage, %C : Consumption percentage
너지의
90.30%,
총섭취에너지의21.53%
인17.24 J
이었으며,
이것은성장이94.03%,
탈피체가5.97%
를각각차지하였다.
고 찰
해산동물의에너지흐름을파악하기위해서는사전에그생물 의생활사를정확하게조사하는것이필요하다
.
갑각류의성장 은환경에따라다양한반응을보이지만(Hartnoll and Dalley,
1981),
특히섭취,
성장및대사율은생육단계별로도많은변동을보이는것으로알려져있다
(Reeve, 1969; Hiroko et al., 1990).
십각목갑각류유생의성장은일반적으로지수적성장을나 타낸다
(Mootz and Epifanio, 1974; Logan and Epifanio, 1978;
Johns and Pechenik, 1980; Stephenson and Knight, 1980;
Dawirs, 1983).
본연구에서실내사육한가시배새우의유생성 장은지수적성장을보였으며, Park (1989)
의실험실내에서사 육한보리새우, Penaeus japonicus
의초기유생의성장에서지 수적성장을보였다.
그리고Her (2002)
의물렁가시붉은새우, Panadalopsis japonica
의초기유생에서의성장에서도 지수적 인성장을보임으로써,
다른갑각류의유생성장양상과도유사 하였다(Dawirs, 1983; Jacobi and Anger, 1985; Harms, 1987;
Kim, 2000).
가시배새우 유생의 산소소비량은 체중이 유생발육이 진행 될수록증가하였다
.
이는Carcinus maenas (Dawirs, 1983)
와Hyas coarctatus (Jacobi and Anger, 1985)
의연구된결과에서 도다른갑각류의유생과유사한결과로이러한체중증가에따 른산소소비량의증가는갑각류의일반적인경향을나타내었 다(Beamish, 1964; Fry, 1971; Dabrowski, 1986; Degani et al., 1989; Cai and Summerfelt, 1992).
수서무척추동물은동화에너지의거의
2/3
를호흡에사용하는 것으로알려져있고(Clutter and Theilacker, 1971),
특히생물 에있어서호흡을통한대사에너지는에너지수지구성요소별 사용에너지의백분율에많은변동을야기할수있다. Chin et
al. (1992)
은호흡열상실에따른에너지계산은동물이서식지에서경험하는수온범위내의여러수온구에서산소소비량 을측정하고
,
이결과로부터얻어진수온및체중과상관관계 를갖는산소소비량회귀식을적용하는것이생물의호흡을통 한대사에너지에대한이해도를높일수있는방법이라고제안 하였다.
가시배새우유생의섭취량은
Park (1989)
이보고한Penaeus japonicus
의zoea
유생보다는훨씬높았으며, mysis
유생과비 슷한섭취량으로나타났다.
그러나Emmerson (1984)
이보고한Penaeus indicus
의protozoea
유생보다는훨씬낮게나타났다.
본연구결과에서
23.84%
의동화효율은, Penaeus japonicus
의17.62% (Park, 1989)
및Carcinus maenas
의22.2% (Dawirs, 1983)
와는 비슷한 수준을 보였다.
하지만Rhithropanopeus harrisii
의45.0% (Levine and Sulkin, 1979), Menippe mer-
cenaria
의63.2% (Mootz and Epifanio, 1974)
및Cancer ir- roratus
의77.0% (Johns, 1982), Neomysis intermedia
의암컷85.65%,
수컷81.99% (Choi et al., 1997)
에비해서는훨씬낮 은값을보였다.
이와같은낮은동화효율은종에따른차이도 있을수있지만,
환경수의온도와비례하는소화기능과밀접한 관련을가지고있기때문에,
본연구에서처럼한해성새우의경 우동화효율이낮은반면분으로배출되는에너지비율이높았 던것으로판단된다.
가시배새우의 총 성장효율
(K
1)
은21.53%
로Penaeus ja- ponicus (Park, 1989)
및Rhithopanopeus harrisii (Levine and Sulkin, 1979), Menippe mercenaria (Mootz and Epifanio, 1974)
와Cancer irroratus (Johns, 1982)
의23-39%
와유사한 결과를보였지만, Carcinus maenas
의3.3% (Dawirs, 1983), Neomysis intermedia
의암컷8.71%,
수컷9.02% (Choi et al., 1997), Pandalus hypsinotus
의12.91% (Kim, 1994)
보다높 았다.
순 성장효율(K
2)
은90.30%
로Pandalopsis japonica
의91.39% (Kim, 2000)
와비슷하게나타났으며동화량에대하여 성장으로전환된에너지가상당히높은것으로나타났다.
이러 한효율적차이는각각의종에대한유생단계와공급된먹이에 서많은영향을받는것으로판단된다.
대사활동과정에서에너지의소비는에너지수지로구체적으 로설명된다
(Adams and Breck, 1990).
다양한생물학적,
물리 적,
화학적조건하에서오는에너지수지에관한생체역학모델 구축(Bartell et al., 1986)
을위한일환에서본연구의가시배새 우유생에너지수지에대한구체적인정보는매우유용하게활 용될것으로판단된다.
사 사
본 연구는농림축산식품부수산실용화기술개발사업에의해 이루어진것임
.
References
Adams SM and Breck JE. 1990. Bioenergetics. In: Methods for Fish Biology. Schreck CB and Moyle PB, eds. American Fisheries Society, Bethesda, MA, U.S.A., 389-415.
Bartell SM, Breck JE, Gardner RH and Brenkey AL. 1986. Indi- vidual parameter perturbation and error analysis of fish bio- energetics models. Can J Fish Aquat Sci 43, 160-168.
Beamish FWH. 1964. Respiration of fishes with special em- phasis on standard oxygen consumption. Ⅱ. Influence of weight and temperature on respiration of several species.
Can J Zool 42, 177-188.
Cai Y and Summerfelt RC. 1992. Effects of temperature and size on oxygen consumption and ammonia excretion by walleye. Aquaculture 104, 127-138.
Chin P, Kim HY and Sin YK. 1992. Energy budgets for the de- velopment stages of Palaemon macrodactylus. Bull Korean
Fish Soc 25, 341-358.
Choi YG, No S, Ju SD and Park KY. 1997. Energy budget of the Mysis shrimp, Neomysis intermedia reared in the Labora- tory. Journal of Aquaculture 10, 289-300.
Clutter RI and Theilacker GH. 1971. Ecological efficiency of a pelagic mysid shrimp: Estimates from growth, energy bud- get and mortality studies. Fish Bull US 69, 93-115.
Dabrowski KR. 1986. Active metabolism in larval and juvenile fish: ontogenetic changes, effect of water temperature and fasting. Fish Physiol. Biochem 1, 125-144.
Dawirs RR. 1983. Respiration, energy balance and developmen- tal pattern in growing and starving larvae of Carcinus means L. (Decapoda, Portunidae). J Exp Mar Ecol 69, 105-128.
Degani G, Gallagher ML and Meltzer A. 1989. The influence of body size and temperature on oxygen consumption of the European eel, Anguilla anguilla. J Fish Biol 34, 19-24.
Emmerson WD. 1980. Ingestion, growth and development of
Penaeus indicus larvae as a function of Thalassiosira weiss- flogii cell concentration. Mar Bio 58, 65-73.
Emmerson WD. 1984. Predation and energetica of Penaeus in-
dicus (Decapoda, Penaeidae) larvae feeding on Brachionus plicatilis and Artemia nauplii. Aquachlture 38, 201-210.
Fry FEJ. 1971. The effect of environmental factors on the physi- ology of fish. In: Fish Physiology. Hoar WS and Randall DJ, eds. Academic Press, New York, U.S.A., 1-98.
Gnaiger E. 1983. Calcultion on energetic and biochemical equivalents of respiratory oxygen consumption. In: Plaro- graphic oxygen sensors. Gnaiger E. and Forstner H, eds.
Springer, Berlin, Germany, 337-345.
Harms J. 1987. Energy budget for the larval development of
Elminius midestus(Crustacea, Cirripedia). Helgolander wiss
Meeresunters 41, 45-67.Hartnoll RG and Dalley R. 1981. The control of size variation within instars of a crustacean. J Exp Mar Biol Ecol 53, 235- Her JY. 2002. Molting and growth of Pandalopsis japonica lar-239.
vae reared at different food and temperature. M.S. Thesis, Gangneung-Wonju National University, Gangneung, Korea.
Hiroko Y, Ceccaldi HJ and Gaudy R. 1990. Combined influ- ence of temperature and salinity on oxygen consumption of the larvae of the pink shrimp, Palaemon serratus(Pennant) (Crustacea, Decapoda, Palaemonidae). Aquaculture 86, 77- Ikeda T. 1974. Nutritional ecology of marine zooplankton. Mem 92.
Soc Fish Hokkaido Univ 23, 1-97.
Jacobi CC and Anger K. 1985. Growth and respiration during the larval development of Hyas coarctatus(Decapoda, Maji- dae). Mar Biol 87, 173-180.
Johns DM and Pechenik JA. 1980. Influence of the water-ac- commodated fraction of No. 2 fuel oil on energetics of Can-
cer irroratus larvae. Mar Biol 55, 247-254.
Johns DW. 1982. Physiological studies on Cancer irroratus lar-
vae. Ⅲ. Effects of temperature and salinity on the partition- ing of energy resources during development. Mar Ecol Prog Ser 8, 112-121.
Kim DH. 1995. Growth, Reproduction and Zoea larval Energy Budget of Pandalus hypsinotus. M.S. Thesis, Cheju Nation- al University, Jeju, Korea.
Kim HY and Chin P. 1991. Growth and energy budget of opos- sum shrimp, Neomysis awatchensis. Kor J Zool 34, 594-609.
Kim JK. 2000. Growth and energy budget of Pandalopsis ja-
ponica larvae. M.S. Thesis, Gangneung-Wonju National
University, Gangneung, Korea.Klekowski RZ and Duncan A. 1975. Physiological approach to ecological energetics. In: Methods for ecological bioener- getics. Grodzinski W, Klekowski RZ and Duncan A, eds.
Blackwell Scientific, London, U.K., 15-64.
Levine DM and Sulkin SD. 1979. Partitioning and utilization of energy during developments of the xanthid crab, Rhithro-
panopeus harrisii (Gould). J Exp Mar Biol Ecol 40, 247-
Logan DT and Epifanio C. 1978. A laboratory energy balance 257.for the larvae and juveniles of the american lobster, Homa-
rus americanus. Mar Biol 47, 381-389.
NFRDI. 2007. A Guide to identification of similar species.
NFRDI 42, 71.
Mootz CA. and Epifanio CE. 1974. An energy budget for
Menippe mercenaria larvae fed Artemia naupii. Biol Bull
Woods Hole Mass 146, 44-55.Park KY. 1989. Energy flow of the oriental brown shrimp,
Penaeus japonicus reared in the laboratory. Ph. D. Thesis,
Pusan National Fisheries University, Busan, Korea.Reeve MR. 1969. Growth, metamorphosis and energy conver- sion in the larvae of the prawn, Palaemon serratus. J Mar Biol Ass, 49, 77-96.
Stephenson MJ and Knight AW. 1980. Growth, respiration and caloric content of larvae of the prawn, Macrobrachium
rosenbergii. Comp Biochem Physiol 66A, 386-391.
Vernberg FJ and Vernverg WB. 1981. Functional adaption of marine organisms. Academic Press, New York, U.S.A., 1-347.