Received: April 4, 2017 Revised: June 27, 2017 Accepted:July 5, 2017
OPEN ACCESS
HORTICULTURAL SCIENCE and TECHNOLOGY 36(2):292-301, 2018
URL: http://www.kjhst.org pISSN : 1226-8763 eISSN : 2465-8588
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Copyrightⓒ2018 Korean Society for Horticultural Science.
본 연구는 한국연구재단 중점연구소지원사업(NRF- 2015R1A6A1A03031413), 경상대 신임교원연 구기반조성 사업(2014-04-014)의 지원에 의해 수 행되었음. 이준성은 교육부 BK21Plus 사업의 장학 금을 수혜 받았음.
유전자 침묵현상을 이용한 고추 리포옥시게네이즈 유사 유전자의 생육과 발달 관련 기능 분석
강원희1† · 이준성2† · 남재영1 · 염선인1,2*
1경상대학교 농업생명과학연구원, 2경상대학교 응용생명과학부(BK21 Plus) 농업식물과학과
Identification of CaLOP Regulating Development and Growth Through Virus-Induced Gene Silencing in Pepper
Won-Hee Kang 1† , Junesung Lee 2† , Jae-Young Nam 1 , and Seon-In Yeom 1,2*
1Institute of Agriculture & Life Science, Gyeongsang National University, Jinju 52828, Korea
2Department of Agricultural Plant Science, Division of Applied Life Science (BK21 Plus program), Gyeongsang National University, Jinju 52828, Korea
*Corresponding author: [email protected]
†
These authors contributed equally to this work.
Abstract
The proteins secreted into the cell wall and extracellular space have multiple roles in plant development, metabolism, and stress responses. In a previous study, examples of such proteins secreted during pepper–pathogen interactions were isolated to elucidate their roles. Among these proteins, we further characterized a lipoxygenase homology gene CaLOP (Capsicum annuum lipoxygenase homology protein), which has a 730 bp cDNA sequence and encodes 184 amino acids. CaLOP contains a lipoxygenase homology (PLAT/LH2) domain. A database search revealed that CaLOP shares significant sequence identity with putative orthologs in other plants. CaLOP was constitutively expressed in root tissue, and was upregulated in response to TMV-P0, TMV-P2, and PepMoV viruses, as well as Phytophthora capsici. To examine the biological function of this gene in plant development, we performed in planta knockdown assays using the Tobacco rattle virus-based gene-silencing system. Silencing CaLOP in pepper resulted in abnormal growth and severe developmental disorders, including lethal phenotypes, compared with controls. Consequently, our results suggest that CaLOP may be important in plant growth and development, and pathogen defense.
Additional key words:
Capsicum, lipoxygenase-homology protein, secreted protein, VIGS
서 언
식물은 끊임없이 외부에서 비생물학적
,생물학적 스트레스의 영향을 받는다
(Lee and Yeom, 2015).식물이 생물학적 스트레 스를 극복하기 위한 방법 중의 하나는 세포벽을 이용하여 세포 밖의 다양한 환경에 대항하는 것이다
(Hugot et al., 2004).특히
,식물은 병원균 침입 초기에 병원균의 분비 유도체를 인지하여 신호전달경로를 활성화하고
,세포벽 및 세포간극으로 분비단백 질
(secretome)을 내보내어 방어기제를 시작한다
(Lee et al., 2004).식물의 다양한 방어기제에 중요한 역할을 하는 것으로 알려 진 분비단백질은 식물이 병원균에 공격을 받았을 때 세포 편광과 유두 형성을 통한 세포벽 비대에 관여하는 것으로 보고되었다
(Schulze-Lefert, 2004).또한
, PR단백질
(pathogenesis-related protein)과 같은 분비단백질은
systemic acquired resistance (SAR)을 통한 식물의 저항성에 기여하는 것으로 보고되었고
(Schmelzer, 2002; Yeom et al., 2012),식물의 형태 변화
,조직 분 화 및 생장과 같은 생육과 발달에도 중요한 역할을 한다고 알려져 있다
(Yamane et al., 2005; Lee et al., 2011; Yeom et al., 2011; Seo et al., 2012).선행 연구에서 고추
(Capsicum annuum L.)와 고추 역병균
(Phytophthora capsici)과의 상호작용을 통해 분비되는 유전자들 을 효모 기반 분비단백질 분리 기법
(yeast-based secretion trap screen)을 통해 고추 분비단백질
(C. annuum secretome, CaS)유 전자를 대량 분리하였다
(Yeom et al., 2011).고추 역병균에 대해 저항성 고추와 감수성 고추에서 특이적으로 발현되는 분비단 백질을 코딩하는 유전자들에 대한 발현 분석을 통해 후보 유전자들을 선별하고
,이들에 대해 바이러스 기반 유전자 침묵 기법
(virus-induced gene silencing, VIGS)을 통해 저항성 기능 분석을 수행하였다
.또한
,병원균과 상호작용하는
CaS유전자가 식 물의 생육과 발달에도 중요한 역할을 할 것이라는 가능성도 제시하였다
(Seo et al., 2012).하지만
,그 유전자들의 다양한 환경 스트레스에 대한 상호작용과 식물의 생육 및 발달에 관한 생물학적 기능에 대해서는 제한적으로 연구가 진행되었다
.본 연구는 선행연구에서 분리된
CaS유전자들 중에서 고추
lipoxygenase유사 유전자인
CaS17(CaLOP)에 대한 식물 생육 및 발달에 관한 기능을 밝히고자 하였다
.이를 위해
RNA-seq정보를 활용하여 다양한 조직에서의 발현 양상 분석 및 세균
,바이 러스
,역병 등 다양한 병원균 침입 시의 발현 양상 분석 등을 통해 다양한 병원균과의 상호작용에 관한 식물 방어 기제 연관성을 추가 분석하였고
,바이러스 기반 유전자 침묵 기법을 이용한 유전자의 발현 억제를 유도하여
CaS17(CaLOP)의 생물학적 기능 을 검정하였다
.또한
, CaS17(CaLOP)과 동종 상동성 유전자인
CaS4와의 비교 분석을 통해 두 유전자 간의 생육과 발달에 관련 된 기능적 차이를 확인하였다
.재료 및 방법
식물재료 및 VIGS, 표현형 분석
광
16시간
,암
8시간의 순환의
23°C생장상
(Kim et al., 2008)에서 키운 고추를 바이러스 기반 유전자 침묵 기법
(VIGS)을 수
행에 사용하였다
. VIGS는
Yeom et al.(2012)에서 보고된
TRV-based VIGS방법을 참조하여 다음과 같이 수행하였다
. DNA염기서열 특이적인
CaS4와
CaS17유전자가 클로닝된
pTRV벡터를
Agrobacterium tumefaciens sp. strain GV2260에 형질전
환 후
, Agrobacterium을
rifampicin 50mg·mL-1과
kanamycin 50mg·mL-1를 포함한
YEP배지
(yeast extract peptone media)에
서
2일 동안
30°C에서 진탕배양하였다
.증식한
Agrobacterium을 원심분리기로 세포를 침전시켜 접종 버퍼
(10mM MES, 10mM MgCl2, pH 5.5)에 풀어준 후
,배양액의 흡광도
(optical density, O.D.)를
0.5로 정량하여 사용하였다
.이 후 고추의 떡잎
에 배양액을
1mL주사기로 접종한 후
24시간 동안
16°C에서 저온배양 후에 광
16시간
,암
8시간 순환의
21°C생장상
(JSPC- 420C, JS Research Inc., Gongju, Korea)에서 키웠다
.표현형은 접종 후
5주차의 고추에서 관찰하였으며 각 개체의 초장과 엽면
적을 측정하였다
.엽면적은 각각의 식물체의
3번째 본엽을 채취하여 측정하였다
.통계분석은
SPSS프로그램
(PASW statistics 18, Chicago, IL, USA)을 사용하여 분산분석
(ANOVA)을 하였다
.분산분석 간에 평균의 비교는
Tukey의 다중비교법을 이용
하였다
.유전자 서열, 구조 및 진화 분석
CaS4
와
CaS17의 전체 염기서열은 고추 유전체 사이트
(http://genome.pepper.snu.ac.kr/)에서 확보하였다
.단백질의 도메인 구조 분석은
SMART(http://smart.embl-heidelberg.de/)를 이용하였다
.고추 외의 토마토
,감자
,담배
,벼
,애기장대에서의 이 종 상동성 유전자 서열은
Sol Genomic Network(https://solgenomics.net/,토마토
,감자
,담배 서열
), Rice Genome Annotation Project(http://rice.plantbiology.msu.edu/), Arabidopsis TAIR(https://www.arabidopsis.org)에서 각각 확보하였다
.유전자 서 열분석
(multiple sequences alignment, MSA)은
EMBL-EBI(http://www.ebi.ac.uk/)의
MUSCLE프로그램을 사용하였다
.진 화 계통수 분석은
Seo et al.(2016)에서 보고된 방법을 참조하여 다음과 같이 수행하였다
. MEGA7(http://www.megasoftware.net/)프 로그램을 기반으로 계통분석 시 방법은
neighbor-joining을 사용하였고
, bootstrap method로
100번 반복하였다
.치환모델은
Jones-Taylor-Thornton(JTT) model을 사용하고
,부분 간의 비율은
uniform rates를 사용하였다
.공백과 잘못된 데이터의 처리 는
partial deletion을 사용하여
95%이하로 겹치는 것은 제거하였다
.RT-PCR
RNA
의 추출은
VIGS수행 후
5주차의 식물체를 대상으로 하여
3-4번째 본엽에서 추출하였으며
Trizol(Invitrogen, Carlsbad, CA, USA)을 이용하여 제조회사의 지침에 따라 진행하였다
. cDNA는
M-MLV Reverse Transcriptase(Promega, Madison, WI, USA)를 사용하여 합성하였다
. RT-PCR에 사용된 프라이머는
CaTin1_C_F(5'-GGCAGTGGTTACGACTACTT-3'), CaTin1_C_R(5'-CATGATAGGACTGAGGATCG-3'), CaLOP_N_F(5'-ACTGCCATGGGAGTAGCTCAGGT-3'), CaLOP_F_C_R(5'-GCCTACACAAATGATGAGATAACGT-3')
을 사용하였다
.조직 특이적 및 병원균 상호작용 발현패턴 분석
고추 유전자의 조직 별
RNA-seq정보는 선행연구 결과
(Kim et al., 2014)를 기반으로
5개의 조직
(잎
,줄기
,뿌리
,과피
,태좌
)에서
7개의 과실 숙도에 따른 발달 단계인 수분 후
6일
(6DPA, days post-anthesis), 16일
(16PDA), 25일
(25PDA),과실의 녹색 기
(mature green, MG),변색기
(breaker, B),변색기 후
5일
(B5), 10일
(B10)에서의 유전자 발현 패턴을 분석하였다
. TMV-P0, TMV-P2, PepMoV,고추역병과의 고추와의 상호작용
RNA-seq정보 및 발현 분석은
Kang et al.(2016)에서 보고된 방법을 기 반으로 병원균을 접종하지 않은 음성 대조군 값으로 정규화
(normalization)한 후에 분석하였다
.각 유전자별 발현패턴은
RPKM(read per kilobase per million mapped reads)값에
log2값을 취하여 수행하였으며
, heatmap은 정규화된
RNA-seq정보 를
heat-map R package(http://bioconductor.org/)를 이용하여 분석하였다
.결과 및 고찰
CaS17 유전자 동정
선행연구를 통해 분리한 분비단백질 유전자
CaS17와
CaS4의 특성 분석을 위해 두 유전자들의 클론을 시퀀싱하여 유전자의
서열을 확보하고
,이 서열들을 고추 유전체 데이타베이스에서
BLAST하여 전체 유전자를 검정하였다
.각 유전자의 확보한 서
열 크기는
CaS4와
Cas17유전자가 각각
UTR을 포함하여
804bp(179aa)와
730bp(184aa)였다
.이 후
,두 유전자들의 기능 및
특성을 예측하기 위하여
NCBI에
BLAST한 결과
, CaS4유전자는 단백질 수준에서 기존에 보고된
CaTin1유전자
(Shin et al., 2003)와
100%로 동일한 유전자였으며
, CaS17유전자는 기존에 보고된
CaTin1-2유전자
(Shin et al., 2003)와
84% coverage, 99% similarity를 가지고 있는 것을 확인하였다
.또한
CaS4와
CaS17유전자를 비교했을 때는 서로
62% similarity를 가지는 것
을 알 수 있었다
.이를 통해
CaS4와
CaS17은 서로 상동성이 높지만
,서로 다른 유전자인 것을 확인하였으며 유사한 기능을 가
질 것으로 유추할 수 있었다
.기존에 보고된 선행 연구 결과를 살펴보면
, CaTin1유전자는
CaTin1-2유전자와 동일한 프로모터 를 사용하며
,두 유전자가 함께
Tobacco mosaic virus(TMV)저항성과 에틸렌의 산화
,환원 반응에 관여한다고 알려져 있다
(Shin et al., 2003; Shin et al., 2004).그러나 다른 병원균에 대한 유전자의 발현 양상 등의 반응성과 작물의 생육과 발달에 대한 기능에 대해서는 연구 결과가 아직 보고되지 않았다
.그러므로 아직 그 기능이 보고 되지 않은
CaS17유전자로 후속 연구를 진 행하였다
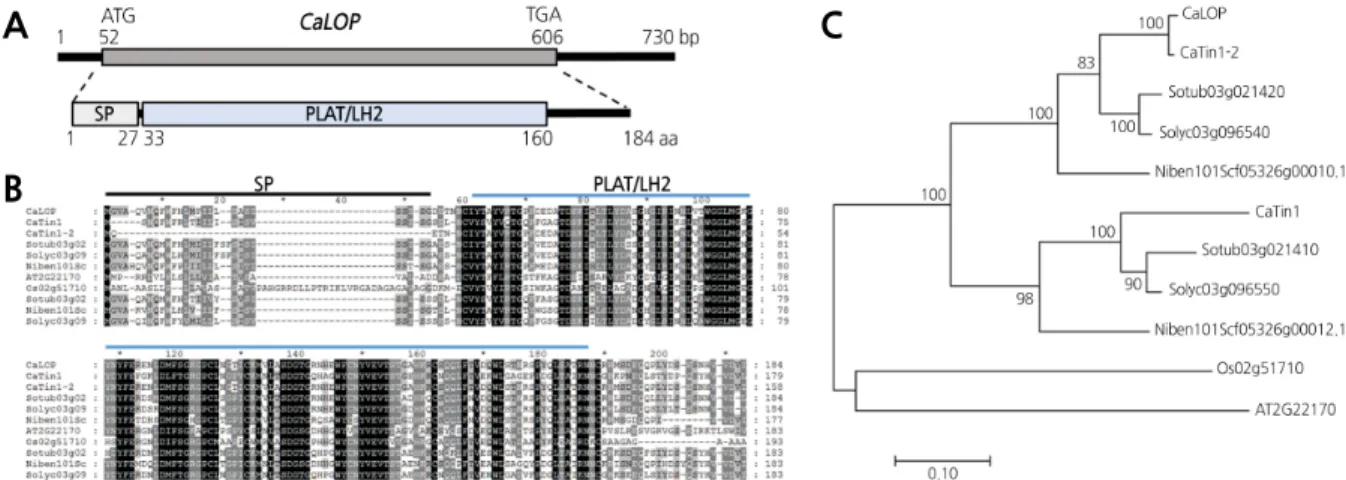
.CaLOP 유전자의 구조 및 계통 분석
CaS17
유전자의 구조적 특징을 확인하기 위하여
, SMART프로그램으로 구조 분석을 수행하였다
.유전자의
5'말단
(1-26aa)에
signal peptide(SP),중간 부분부터
3'말단
(33-160aa)에는
PLAT/LH2구조를 가지고 있는 것을 확인하였다 (Fig. 1A). 예측 한 구조 중에서
PLAT(ploycystin-1, lipoxygenase, alpha-toxin)도메인과
LH2(lipoxygenase homology)도메인은
lipoxygenase와 관련된 것으로
CaS17도
lipoxygenase의 역할을 할 것으로 예측할 수 있었다
.이에 따라
CaS17유전자를
C. annuum lipoxygenase homology protein(CaLOP)로 명명하였다
.CaLOP
유전자의 상동유전자 간의 진화 및 특징 차이를 분석하기 위하여
CaLOP유전자와 동종 상동성 유전자인
CaTin1-2, CaTin1(CaS4)와 다른 작물의 이종 상동성 유전자들 간의 단백질 서열 비교 및 계통 분석을 하였다
. CaLOP와
CaTin1의 서열 을 바탕으로 감자
(Sotub03g021410, Sotub03g021420),담배
(Niben101Scf05326g00010.1, Niben101Scf05326g00012.1),토 마토
(Solyc03g096540, Solyc03g096550),벼
(Os02g51710),애기장대
(AT2G22170)에서 상동유전자를 확보하였다
.벼와 애 기장대의 경우 두 유전자와 상동성이 높은 유전자가 각각 하나씩 확인되었다
.이들 유전자들은
CaLOP유전자와 동일하게 모 두
SP와
PLAT/LH2도메인 구조를 가지고 있었고
,단백질 서열 간의 상동성이 높은 것을 확인하였다 (Fig. 1B). 계통도 분석
(phylogenetic analysis)을 통한 진화분석을 수행한 결과 총
3개의 그룹으로 분류할 수 있었으며
,가지과 작물끼리 그룹으로 나
A
B
C
Fig. 1. Structural and phylogenetic analysis of CaLOP and its homologs. A Structure of CaLOP. The CaLOP gene comprises two exons and one intron. The CaLOP protein contains a signal peptide (SP) and lipoxygenase homology 2 (PLAT/LH2) domain. Numbers shown underneath the CaLOP structure represent the location of each domain on the gene. B Comparative analysis of predicted amino acid sequences of CaLOP and its homologs. Bars with SP and PLAT/LH2 above the sequence alignment indicate the signal peptide and lipoxygenase homology 2 domains, respectively. C Phylogenetic tree of CaLOP, and homologs of CaLOP. CaLOP was clustered with CaTin1-2 (Capsicum annuum), Sotub03g021420 (Solanum tuberosum), Solyc03g096540 (Solanum lycopersicum), and Niben101Scf05326g00010.1 (Nicotiana sylbestris).
CaTin1 was clustered with three genes (Sotub03g021410 from Solanum tuberosum, Solyc03g096550 from Solanum
lycopersicum, and Niben101Scf05326g00012.1 from Nicotiana sylbestris). Rice (Os02g51710) and Arabidopsis
(AT2G221700) genes were clustered at the same clade. The tree is drawn to scale, with branch lengths measured in the
number of substitutions per site.
뉜 것이
2개
,벼와 애기장대가 함께
1개의 그룹으로 나뉘었다
.가지과 내에서는
CaLOP유전자와
CaTin1유전자를 중심으로
2개의 그룹으로 나뉘었다
. CaLOP유전자와
CaTin1-2, Sotub03g021420, Solyc03g096540, Niben101Scf05326g00010.1유 전자가 하나의 그룹으로 분류되었고
, CaTin1유전자와
Sotub03g021410, Solyc03g096550, Niben101Scf05326g00012.1유 전자가 다른 한 그룹으로 분류되었다 (Fig. 1C). 이 결과를 통하여
, CaLOP와
CaTin1은 가지과 작물이 종 분화되기 이전에 원시 기원 유전자에서
CaLOP와
CaTin1이 각각 생성되어 진화한 것으로 예측할 수 있었다
.또한
, CaTin1-2유전자와
CaLOP유전 자는 가지과 종분화 이후에 복제 기작에 의해 동종 상동성 유전자가 되었다는 것을 시사한다
. CaLOP유전자는 감자
,토마토
,담배에서 각각 이종 상동성의 유전자들의 유전자 구조가 잘 보존되어 있는 것을 알 수 있었다
.또한
,벼와 애기장대에서도 유사 성이 높은 유전자가 있는 것으로 보아 식물체 내에서 보존된 기능을 수행하고 있을 것으로 보인다
.A
B C
D E
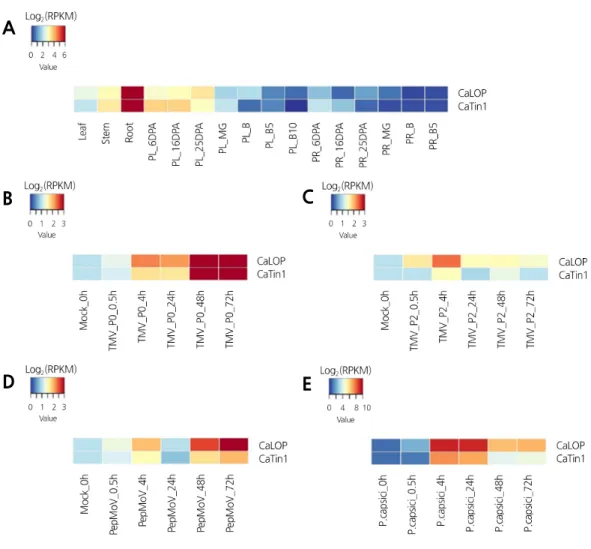
Fig. 2. Heat maps showing the expression patterns of CaLOP and CaTin1 in various tissues, and responses to pathogen
infection in pepper. Tissue-specific expression profiles (A) were analyzed in various tissues and developmental stages in
pepper. PR, pericarp; PL, placenta; MG, mature green; B, breaker stage; DPA, days post-anthesis; B5 and 10, 5 and 10
days post-breaker stage. The expression values were normalized by log2 (RPKM, Reads Per Kilobase of transcript per
Million mapped reads). The expression patterns of CaLOP and CaTin1 were examined in response to TMV-P0 (B),
TMV-P2 (C), PepMoV (D), and P. capsici (E). The RPKM expression values of responses to pathogen infection were
normalized relative to expression values and transformed log
2values. Blue represents low expression; red represents high
expression.
발현 패턴 분석
C. annuum cv. CM334
의
RNA-seq결과를 이용한 발현양상 분석을 통하여
CaLOP의 기능을 예측하고자 하였다
. CaLOP유전자의 발현 양상을 비교 분석하기 위해
CaTin1유전자도 함께 분석을 수행하였다
. CaLOP유전자와
CaTin1유전자의 고추 조직 별 및 과실의 발달 단계별 발현패턴을 분석한 결과
,두 유전자는 테스트한 조직 모두에서 비슷한 발현 패턴을 보였다 (Fig.
2A). 두 유전자 모두 잎보다는 줄기
,줄기보다는 뿌리에서의 발현 양이 많았으며
,과실의 발달 단계에서는 과피
(PR)보다는 태 좌
(PL)에서 발현이 더 많이 되는 것을 알 수 있었다
.또한 태좌에서 과실의 발달단계에 따른 발현양상을 살펴보면
,초기 발달 단 계인 개화 후
6일
(6DPA)부터
25DPA에서 발현이 많이 되고
,그 이후 후기 단계인 녹색기와
(MG)변색기
(B)에서는 발현이 감 소하는 것을 확인하였다 (Fig. 2A). 이를 통해
CaLOP및
CaTin1은 생식 생장에 관련된 조직보다는 영양 생장에 관련된 조직에 서 발현이 증가하여
,영양 생장 조직의 형성 및 발달에 영향을 미칠 것으로 예측할 수 있었다
.CaLOP
와 원시유전자 및 동일 프로모터를 공유한
CaTin1과 상동 유전자
CaTin1-2는
TMV-P0에 대해 바이러스 접종 후 각 각
36시간과
48시간부터 발현이 증가한다는 결과를 통해
CaTin1과
CaTin1-2는
TMV저항성과 관련이 있는 것으로 보고되었 다
(Shin et al., 2003).그러므로
, CaLOP도
TMV를 포함한 다른 병원균에 대해서 저항성 반응과 관련이 있는지를 확인하기 위 해
, 3가지 바이러스
[Tobacco mosaic virus(TMV)–
Pathotype0(P0), TMV-P2, Pepper mottle virus(PepMoV)]와 고추 역병
(Phytophthora capsici)을 접종한
RNA-seq정보를 이용하여 발현양상을 분석하였다
.조직 발현 분석과 마찬가지로
CaTin1도 함께 분석하여 발현 양상을 비교하였다
.본 연구에서 사용한
RNA-seq재료인
C. annuum cv. CM334는
TMV의
pathotype에 따라서 각기 다른 저항성 양상을 보이며
, TMV-P0에는 대해서는 저항성으로
, TMV-P2에 대해서는 감수성으로 보고되었다
(Tomita et al., 2011; Yang et al., 2012).CaLOP
와
CaTin1은
TMV-P0를 접종하였을 때
,접종 후
30분
(0.5h)부터 발현이 증가하기 시작하여
,시간이 지날수록 유전 자의 발현이 점점 증가하는 양상을 보였다
.테스트한 시간 중
48hpi와
72hpi에서 유전자의 발현이 다른 시간대에 비해 매우 강 하게 나타나는 것을 확인할 수 있었다 (Fig. 2B). 이 결과를 통해서
CaTin1은 선행연구에서 보고된 것과 마찬가지로
48시간 이 후부터 발현이 매우 높게 증가한다는 것을 확인하였으며
, CaLOP는
CaTin1과 비슷한 양상을 보이면서
, 48hpi부터 강하게 발 현되는 것을 알 수 있었다
.그리고
TMV-P2를 접종한 샘플에서는
TMV-P0를 접종한 샘플처럼
CaLOP가
CaTin1보다 많은 발 현량이 보였지만
,전체적인 발현 양상을 보면
, TMV-P0를 접종했을 때보다
TMV-P2를 접종한 샘플에서의 두 유전자 발현이 상대적으로 모두 약하게 발현하는 것을 확인하였다 (Fig. 2C).
CaLOP
유전자는
CM334가 감수성을 보인
TMV-P2보다 저항성을 보인
TMV-P0에서 강하게 발현하는 양상을 보였고
,이 를 통하여
CaLOP유전자도
TMV에 대한 저항성에 관여한다는 것을 유추할 수 있었다
.또한
PepMoV를 접종한 샘플에서도
CaLOP유전자는
PepMoV접종 전
(0h)보다 접종 후에 발현이 증가하여
72hpi에서 가장 발현이 높았다 (Fig. 2D). 고추 역병을 접종한 샘플에서는 접종 후 초기 단계인
6hpi와
24hpi에서 발현이 증가하였지만
, 48hpi와
72hpi에서는 이전 시기에 비해 상대 적으로 발현이 감소하였다
. CaLOP유전자뿐만 아니라
CaTin1유전자에서도 비슷한 양상을 확인하였고
,고추 역병과 관련하 여 접종 후 초기 단계에 저항성과 관련이 있는 것으로 생각되었다 (Fig. 2E). 지금까지의 결과를 토대로
,생물학적 스트레스에 대한
CaLOP유전자는 기존에 상동 유전자에서 보고된
TMV이외에도 고추 역병과
PepMoV와 같은 다양한 생물학적 스트레 스와의 상호작용에 관여한다는 것을 시사한다
.CaLOP
유전자의 식물의 발달 및 생육 관련 기능 분석
CaLOP
유전자의 식물의 발달 및 생육에 관련된 기능을 분석하기 위해 고추에서
VIGS를 이용하여 유전자의 발현을 억제하
였다
. CaLOP유전자의 비교 분석을 위하여
CaTin1유전자도
TRV유래 벡터를 사용하여
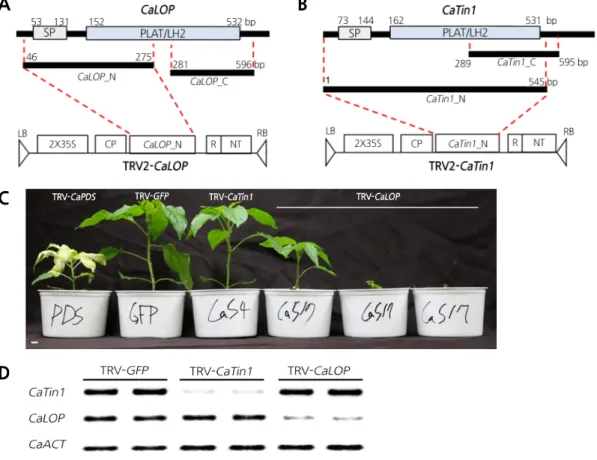
VIGS를 수행하였다 (Fig. 3A and
3B). 대조군으로는
green fluorescent protein(GFP)와
endogenous phytoene desaturase(PDS)유전자에 대한
VIGS실험을 수
행하였다
.이 후
, VIGS수행
5주 후에 식물체의 표현형 변화를 관찰하였다
. GFP를 이용하여
VIGS를 수행한 대조군
(TRV-GFP)
은
CaTin1의 발현을 억제한 식물
(TRV-CaTin1)과 비교했을 때
,생육에서 차이를 보이지 않았다 (Fig. 3C). 그러나
CaLOP유전자의 발현을 억제한 식물
(TRV-CaLOP)에서는 초기 생육단계에서 고사 또는 잎의 변형과 함께 초장의 단축을 보 이거나 생육 저해로 인한 초장의 단축 현상만 보이는
3가지 형태의 표현형을 보였다 (Fig. 3C). 동일한
CaLOP유전자의 발현을 억제하였을 때
,생육이 억제되는 동일한 현상이 나타나지만
,그 안에서의
3종류의 다른 표현형이 나타나는 이유는
VIGS의 효 율에 따라서 각기 다른 표현형을 보이는 것으로 예상할 수 있었다
. VIGS를 수행한지
5주째 되는 식물에서 샘플링을 수행하여 각 유전자들의 발현 억제 유무를 확인하고자
RT-PCR을 수행하였다
.그 결과
,비교 유전자
CaTin1와
CaLOP의 발현을 억제한 식물체에서 각 해당 유전자만 특이적으로 발현이 억제된 것을 확인하였고
,이는 앞서 관찰한 표현형이 유전자 침묵현상으로 인 해 관찰된 것임을 확인할 수 있었다 (Fig. 3D).
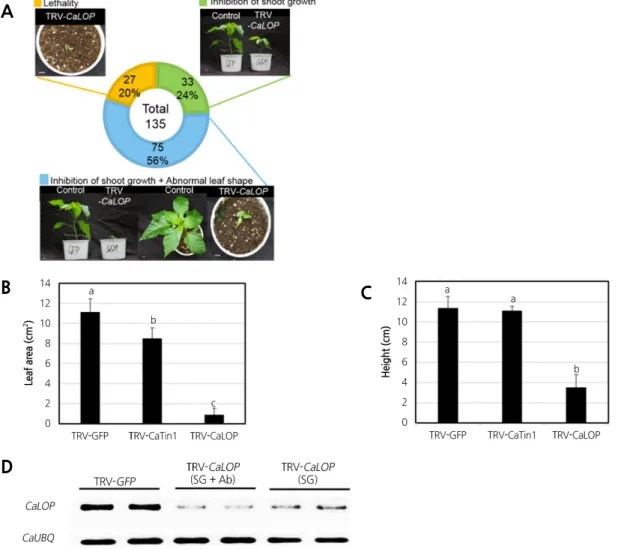
반복 실험을 통해
CaLOP유전자의 발현 억제를 통한 식물체 생육과 발달의 변화 양상을 조사하였다
.여러 번의 반복실험을 통하여 총
135개체의 표현형을 관찰하였고
, 135개체의 식물체 중
56%에 해당하는
75개체의 식물에서 초장이 매우 작고
,잎의 모양이 심하게 변형되는 표현형을 보였고
, 27개체
(20%)의 식물체는
CaLOP유전자가 강하게 발현이 억제되면서 초기 생육 단 계에서 고사하였다 (Fig. 4A). 그리고 나머지
33개체
(24%)의 식물체는 대조군과 비교하였을 때 초장의 길이 및 잎의 크기가 작
A B
C
D
Fig. 3. Silencing effects of CaLOP and CaTin1 on pepper growth and development. A Diagram of the silencing construct
TRV-CaLOP. CaLOP_N, targeting the N terminus of CaLOP, was designated TRV-CaLOP, which contains signal peptide
(SP) and lipoxygenase homology 2 PLAT/LH2 domains. N, N-terminal region used for the Tobacco rattle virus
(TRV)-based virus-induced gene silencing (VIGS) system. B Diagram of the silencing construct TRV-CaTin1. CaTin1_N
was used as TRV-CaTin1, including the N-terminus of CaTin1. C Phenotype of CaLOP-silenced and CaTin1-silenced
pepper. The picture was taken 5 weeks after silencing. TRV-CaPDS and TRV-GFP were used as controls. D Semi-quantitative
reverse-transcription polymerase chain reaction expression analysis of CaLOP and CaTin1 in TRV-GFP, TRV-CaLOP
(N-terminal) and TRV-CaTin1 (N-terminal)-infiltrated pepper. The CaACT gene was used as a control.
아진 것으로 확인되었다 (Fig. 4A). 식물체의 엽면적과 초장을 측정하여 대조군과 비교하였다
.그 결과
VIGS 5주차 식물체의 엽면적은
TRV-GFP는 평균
11.1cm2, TRV-CaTin1은
8.5cm2, TRV-CaLOP는
0.9cm2로 나타났으며
, TRV-GFP와 비교하였 을 때
, TRV-GFP보다
TRV-CaTin1은
2.6cm2, TRV-CaLOP 10.2cm2가 작아진 것을 확인할 수 있었다 (Fig. 4B). 초장에서는
TRV-GFP는 평균
11.3cm로 확인 되었고
, TRV-CaTin1의 초장은
11.1cm로 확인되어
TRV-GFP와 비교하였을 때 유의한 차 이를 보이지 않았다
.하지만
TRV-CaLOP식물체에서는 평균
3.5cm로
TRV-GFP와 비교하였을 때 초장이 평균
7.8㎝ 더 작아 진 것을 확인할 수 있었다 (Fig. 4C). VIGS 수행 후
5주째 되는 식물을 샘플링을 하여
CaLOP발현억제 시
3가지 표현형에 따른 유전자 발현을 확인하였다
.식물이 고사하는 표현형을 가진 식물체에서는 식물체의 고사로 인해 샘플링을 수행할 수 없었다
.A
B C
D
Fig. 4. Phenotype analysis of CaLOP-silenced pepper. A Phenotypic classification of CaLOP-silenced plants. CaLOP-silenced
plants are categorized into three classes, each with a specifically colored square box on the left side of the class
description. The circular diagram indicates the percentage of each class. Representative CaLOP described in each class is
shown. The picture was taken 5 weeks after silencing. A total of 135 plants were silenced, with three independent
experiments. A representative result is shown. Control (TRV-GFP) in this figure indicates GFP-silenced pepper. B, C
Measurement of height and leaf area in silenced plants. The plant leaf area and height was measured 5 weeks after
silencing each gene in pepper. Error bars represent standard deviation. Different letters above bars denote statistically
different values according to Tukey’s honest significant difference test, p ‹ 0.05]. D Semi-quantitative reverse-transcription
polymerase chain reaction expression analysis of CaLOP in TRV-GFP, TRV-CaLOP (N-terminal) with inhibition of shoot
growth phenotype and abnormal leaf shape (SG + Ab), and TRV-CaLOP (N-terminal) with inhibition of shoot growth
phenotype (SG)-infiltrated pepper. The CaUBQ gene was used as a control. SG, inhibition of shoot growth; SG + Ab,
inhibition of shoot growth and abnormal leaf shape.
RT-PCR
결과
,초장이 단축과 잎이 작아지는 표현형
(SG)을 가진 식물체보다 잎 모양이 심하게 변형되고 초장이 매우 작아진 식물체
(SG + Ab)에서
CaLOP의 발현이 더 억제되는 양상을 보였다 (Fig. 4D). 이 결과를 통하여
,동일한
CaLOP유전자의 발 현을 억제하였을 때
,각기 다른 표현형이 나타나는 이유는
VIGS의 효율에 의한 것임을 알 수 있었다
.이러한 결과들을 통하여
, CaLOP은
CaTin1유전자와 달리 식물의 생육과 발달에 핵심적인 기능을 수행한다는 것을 알 수 있 었다
. Fig. 4A의 표현형 조사결과에서
56%의 개체가 해당되는 많은 수의 식물체가 키가 매우 작아지고 잎의 모양이 정상적으 로 발달하지 않는 표현형을 나타내는 것으로 보아
, CaLOP유전자는 식물의 생육에 깊은 관련이 있고
,동시에 발달에도 관여하 여 초장 및 잎의 형태를 유지하는 데에 중요한 역할을 할 것으로 생각된다
.초 록
식물 세포벽 및 세포 사이에 존재하는 분비단백질은 식물 생육과 발달뿐만 아니라 다양한 환경 스트레스 조절 기작에 관여한 다
.선행 연구에서 고추와 병원균의 상호작용을 통해 분비된 고추 유래 분비단백질의 대량 동정을 통해 방어 기제 연구가 진행 되었다
.이 중 병원균 상호작용 분비 단백질의 작물 발달 및 생육 관련 연구를 위해
CaLOP유전자를 선발하였다
. CaLOP유전 자는
PLAT/LH2도메인을 가지고 있었으며
,같은 가지과 식물인 토마토
,감자
,담배 이외에도 애기장대와 벼에서 이종 상동성 유전자들을 확인할 수 있었다
.발현 양상을 살펴보면
, CaLOP유전자는 뿌리 조직에서 강하게 발현되었으며
,고추 역병뿐만 아 니라
TMV-P0, TMV-P1,2및
PepMoV바이러스와의 상호작용 과정의 특정 시기에 따라 발현이 증가하였다
.식물 생육 및 발 달에서의 기능을 확인하기 위해
,유전자의 침묵 현상을 유도하여
CaLOP유전자의 발현을 억제한 결과 대조군에 비해 엽면적 이 작아지고
,초장이 감소하는 등의 비정상적 생육과 생장점의 고사 등의 심각한 발달장애가 나타났다
.이를 통해
CaLOP유전 자는 병원균에 대항하는 방어기제 이외에도 식물의 생육과 발달에 중요한 기능을 하는 유전자라는 것을 확인할 수 있었다
.추가 주요어:
고추
,리포옥시게네이즈 유사 유전자
,분비단백질
,유전자 침묵 현상
Literature Cited