325
[Note]
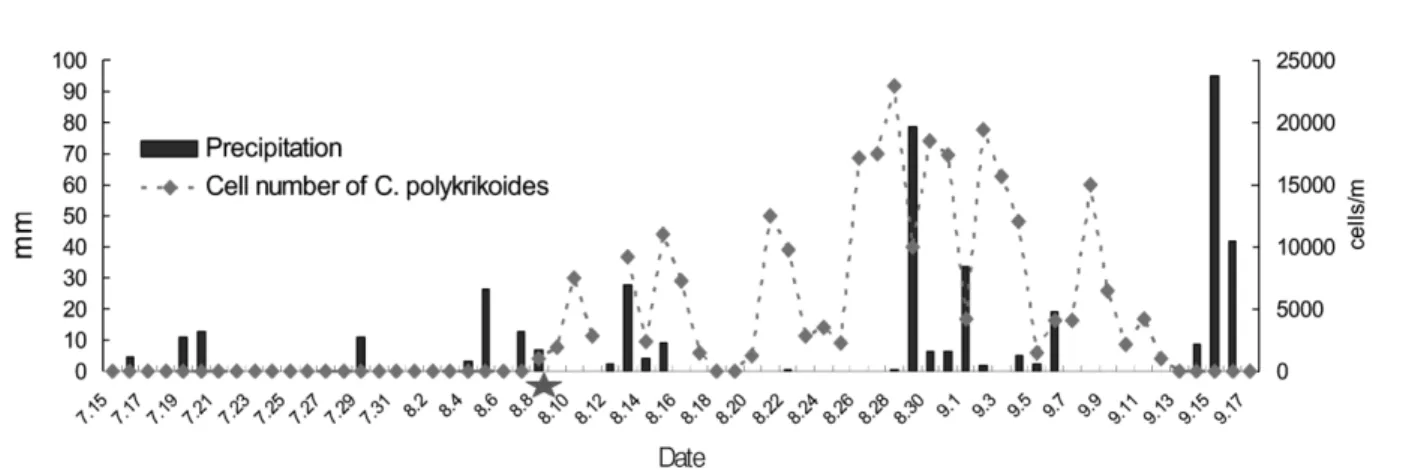
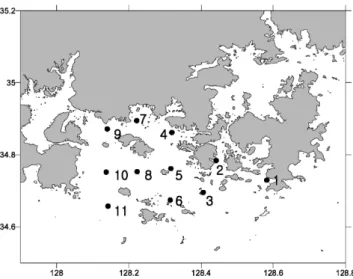
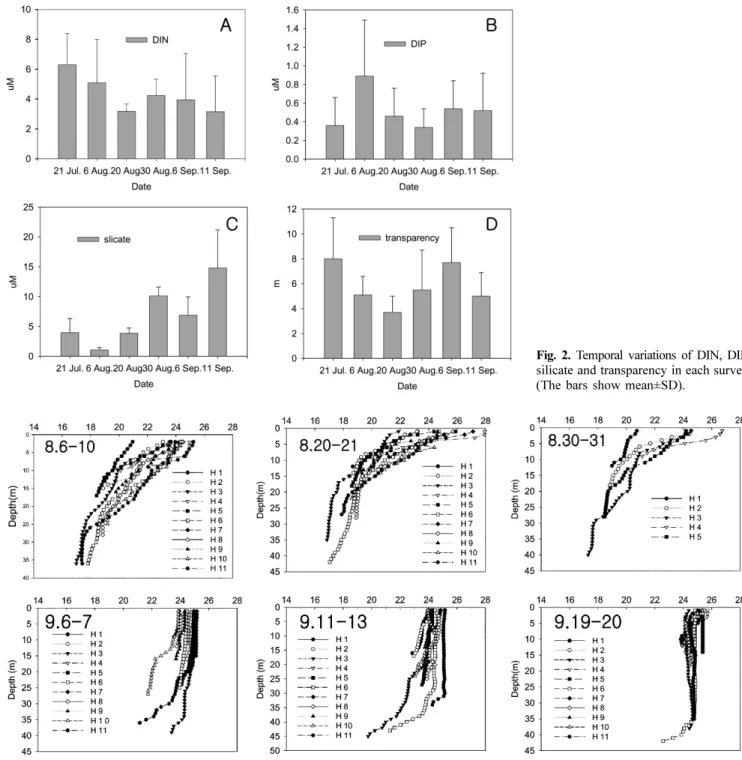
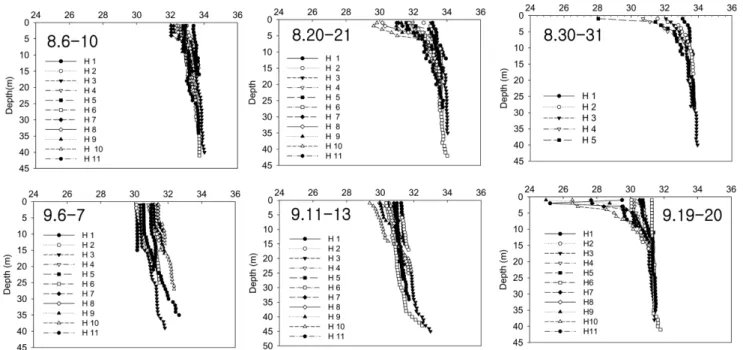
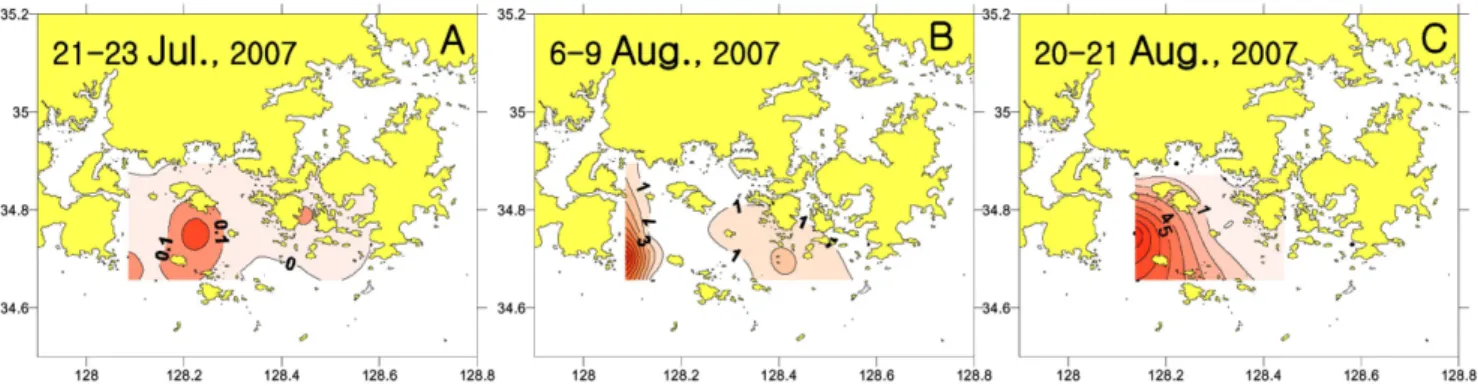
2007년 남해동부해역의 Cochlodinium polykrikoides 적조 발생과 소멸에 미치는 환경 특성
임월애*·이영식·이삼근
전체 글
임월애*·이영식·이삼근
수치
관련 문서
Monthly variation of species richness, evenness and di- versity of fishes collected by set net fishery in coastal waters of Sirang-ri, Gijang-gun from 2007 to 2009... 동해