Kor J Fish Aquat Sci 47(3),241-246,2014
한수지 47(3), 241-246, 2014Original Article
241
Copyright © 2014 The Korean Society of Fisheries and Aquatic Science pISSN:0374-8111, eISSN:2287-8815
서 론
참돔이리도바이러스병
(RSIVD, red seabream iridovirus dis- ease)
은1990
년일본시코쿠지역의참돔양식장에서처음발 병된후,
매년발병지역의확산되고있다(Inouye et al., 1992).
원인체가 되는
Megalocytiviruses
는major capsid protein (MCP) gene, adenosine triphosphatase (ATPase) gene
등을사 용한계통적분석에의해4
개의subgroup
으로분류되고있다(Imajoh et al., 2007). Subgroup 1
은RSIV
를포함하고있으며, subgroup 2
는우리나라의주요병원체로서돌돔(Oplegnathus fasciatus)
등에매년대량폐사를유도하는rock bream irido- virus (RBIV)
를포함한다. Subgroup 3
은중국의담수식용어인mandarin fish (Siniperca chuatsi)
에서분리된infectious spleen and kidney necrosis virus (ISKNV)
가대표적이며, subgroup
4
는최근넙치(Paralichthys olivaceus)
와터봇(Scophthalmus maximus)
에서 분리된flounder iridovirus (FLIV)
와turbot reddish body iridovirus (TRBIV)
을 포함하고 있다.
이러한subgroup
중최근까지국내의어류양식장에서는subgroup 2
와4
에의한질병의발생만이보고되어있다(Do et al., 2005;
Oh et al., 2006; Jeong et al., 2003).
패류는여과섭식
(filter-feeding)
을 통해먹이섭취를하는과 정중norovirus
등의human enteric virus
뿐만아니라marine birnavirus (MABV), white spot syndrome virus (WSSV)
와같 은수생동물에영향을줄수있는바이러스들이중장선(diges- tive gland)
에축적되어존재하는것이보고되고있다(Atmar et al., 1995; Gerba et al., 1978; Goyal et al., 1979; Suzuki and Nojima, 1999; Vazquez-Boucard et al., 2010).
특히Megalo-
cytivirus
는국내에서매년주기적으로발병하고있으나최근HRM 분석법을 이용한 패류 내 Megalocytiviruses의 검출과 유전적 분석
김광일·진지웅
1·김영철
1·정현도
1*
국립수산과학원 수산생물방역과, 1부경대학교 수산생명의학과
Detection and Genetic Differentiation of Megalocytiviruses in Shellfish, via High-Resolution Melting (HRM) Analysis
Kwang Il Kim, Ji Woong Jin
1
, Young Chul Kim1
and Hyun Do Jeong1
*Aquatic Life Disease Control Division, National Fisheries Research & Development Institute, Busan 619-902, Korea
1
Department of Aquatic Life Medicine, Pukyong National University, Busan 608-737, Korea
Viruses in the genus Megalocytivirus have been subdivided into four subgroups. Among these subgroups 2 and 4, represented by the red sea bream iridovirus (RBIV) and the olive flounder iridovirus (FLIV), respectively, are non- exotic. subgroups 1 and 3, represented by the red sea bream iridovirus (RSIV) and the infectious spleen and kidney necrosis virus (ISKNV), respectively, have not been detected in Korea and are known as exotic. Shellfish are filter- feeders, and can thus filter and accumulate Megalocytivirus in their digestive glands, allowing us to track viral con- tamination in surrounding aquatic environment. In this study, we developed a high-resolution melting (HRM) analy- sis to differentiate among subgroups of Megalocytivirus accumulated in shellfish, and confirmed the convenience and efficiency of this method. More than two subgroups of Megalocytivirus were found in the digestive gland of a single shellfish. We classified all Megalocytivirus viruses from shellfish in Korea into subgroups 2 and 4, although proportions of subgroups were different among regions. Compared to nucleotide sequencing analysis, HRM analysis is a simple and rapid method for differentiating of Megalocytivirus subgroups.
Key words: Megalocytivirus, Differentiation , 2-step PCR, High-resolution melting, Shellfish
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial Licens (http://creativecommons.org/licenses/by-nc/3.0/)which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
http://dx.doi.org/10.5657/KFAS.2014.0241 Kor J Fish Aquat Sci 47(3) 241-246, June 2014
Received 21 April 2014; Revised 27 May 2014; Accepted 16 June 2014
*Corresponding author: Tel: +82.51.629.5941 Fax: +82. 51. 629. 5938
E-mail address: [email protected]
에야국내에서식하는패류내로의축적및분석방법에대한접 근이겨우이루어지고있는실정이다
(Kim et al., 2011).
더구나 현재까지패류내에축적된Megalocytivirus
의유전적분류에 대한신속한분석방법이마련되지않고있어보다정밀한패류 내바이러스의위험성에대한유전적정보가충분히이루어지 지않고있다.
현재
PCR
방법과함께자주사용되고 있는high-resolution melting (HRM)
기법은PCR
수행시intercalating fluorescent dye
를첨가하여PCR
과정에서생성되는amplicon
에결합시키 고,
이후온도상승을줌으로써target
부위의염기서열에따라 서각기다른형태로나타나는fluorescence signal
의변화특성 을분석하는방법이다(Kristensen and Dobrovic, 2008; Tajiri- Utagawa et al., 2008; Wittwer et al., 2003).
따라서HRM
기법 의적용은PCR target
부위에서의variation
을보다빠르고손 쉽게구별함으로써패류내에축적된Megalocytivirus
의진단과함께
subgroup
까지의유전적특성을분별확인을가능하게하여어류질병발생과패류내축적원인체간의상관관계를 확립할수있게한다
.
본연구에서는
Megalocytivirus
의패류중장선에의축적을확 인하고,
이들바이러스의유전적분류를위하여, MCP
유전자부위에서나타나는유전적변이특성에대해
HRM
기법을적용하여
subgroup
을확인하는기법을확립하고국내발생RSIV
질병원인체에대한유전적의미를보고하고자한다.
재료 및 방법
실험재료
본실험에사용된굴은
2010
년3
월진하(
동해지역)
및2011
년3
월고성(
남해지역)
에서채취한굴(Crassostrea gigas)
그리고2011
년9
월서산(
서해지역)
에서바지락(Tapes philippinarum)
을채취하였다.
채취한각각의패류로부터중장선을각각분리 하여-70℃
에보관하면서사용하였다.
바이러스
2000
년9
월에 남해안 양식장에서Megalocytivirus
에 감염 된 돌돔비장에서 분리한IVS strain (subgroup 2, Jeong et al., 2003, GenBank accession numbers for ATPase gene : AF487899)
과2004
년국내로수입되는관상어중Megalocyti- virus
에감염된pearl gourami (Trichogaster leeri)
의비장에서분리한
PGIV-K1 strain (subgroup 3, Jeong et al., 2008)
을숙 주세포인GF
세포(ATCC CCL-58)
에접종하여배양하였다.
배 양방법으로70-80%
배양된단일층의GF
세포에감염비장조 직마쇄액200 μL (10 mg/mL)
를접종하여25℃
에서배양하는 방법을사용하였다. 2-3
일배양한후cytopathic effect (CPE)
가일어나부착된세포가떨어지면15 mL tube
에회수하여냉 동과해동의과정을3
번씩수행한후1,500 × g
에서10
분간원 심분리하여상징액1 mL
씩분주하여서-70℃
에보관하여사 용하였다.
그리고2010
년11
월포항넙치양식장에서Mega- locytivirus
에감염된넙치의비장으로부터분리된FLIV strain (subgroup 4)
을-70℃
에보관하여사용하였다.
DNA의 추출
패류의중장선
50 mg
또는배양된바이러스상등액200 μL
로 부터AccuPrep
®Genomic DNA Extraction Kit (Bioneer, Ko- rea)
을사용하여제조사의protocol
에따라서DNA
를분리하여50 μL
의TE buffer
에현탁하였다.
분리된DNA
는분광광도 계(Eppendorf
®BioPhotometer)
를 사용하여 흡광도측정으로A260/A280 nm
값을구하여DNA
의양을측정하였으며실험 전까지-20℃
에서보관하였다.
2-step PCR
추출한
DNA
를template
로사용하여1-step PCR (Applied Biosystems
®2720 Thermal Cycler)
를아래와같은방법으로 실시하였다. 10× PCR buffer 2 μL, 200 μM
의각각의dNTP, 1 μM
의sense primer
와1 μM
의antisense primer (Table 1), Taq DNA polymerase (G-Taq DNA polymerase, Cosmogentech, Korea)
및template 1 μL
를첨가한후증류수로최종액의vol- ume
이20 μL
가되도록했다. PCR
혼합물은94℃
에서3
분간pre-denaturation
시킨후, 94℃
에서30
초denaturation, 55℃
에서
30
초annealing, 72℃
에서30
초extension
의반응을35 cycle
수행한후72℃
에서7
분간post-extension
시켰다. 2-step PCR amplification
은1-step PCR product 1 μL
를template
로 사용하여위와동일한방법으로실시하였다.
PCR
후증폭산물은0.5 μg/μL EtBr (Ethidium Bromide)
이 첨가된2% agarose gel
을이용하고, 1×TAE buffer (40 mM Tris-acetate, 1 mM EDTA)
를전기영동을위한완충액으로하 여전기영동을실시하였다. UV
검출기에서나타나는band
를 관찰하여그증폭여부를확인하였다.
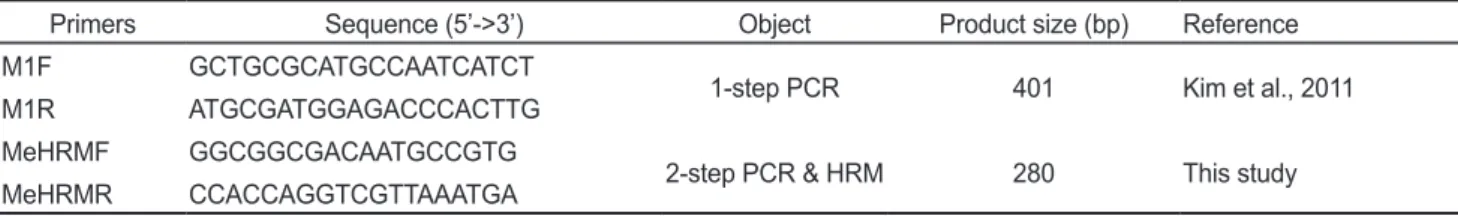
Table 1. Primers information used in this study
Primers Sequence (5’->3’) Object Product size (bp) Reference
M1F GCTGCGCATGCCAATCATCT
1-step PCR 401 Kim et al., 2011
M1R ATGCGATGGAGACCCACTTG
MeHRMF GGCGGCGACAATGCCGTG
2-step PCR & HRM 280 This study
MeHRMR CCACCAGGTCGTTAAATGA
HRM분석법을 이용한 Megalocytivirus의 유전적 분석
243
Cloning 및 Plasmid 분리
각지역에서가장뚜렷한
2-step PCR
양성을보이는시료를 하나씩택하여amplicon (280 bp)
을GeneAll
®Expin Gel SV kit (GeneAll Biotechnology, Korea)
을 사용하여정제하였으 며,
정제된DNA
를pGEM-T Easy vector (Promega, USA)
에ligation
후E. coli DH5α
균주에transformation
시켰다. Am- picillin (50 μg/mL)
과X-gal (5-bromo-4-chloro-3-indolyl-β- D-galactopyranoside, Sigma, USA; 40 μg/mL)
이첨가된LB (Luria-Bertani, Difco, USA)
평판배지에서 증식된colonies (clones)
를무작위로선택하여LB broth
에접종후37℃
에서24
시간 배양하였다.
배양액으로부터GeneAll
®Plasmid SV Mini kit (GeneAll Biotechnology, Korea)
를이용하여plasmid DNA
를분리하였으며이를HRM
분석에사용하였다. HRM 분석기법을 이용한 Megalocytivirus의 sub- group 분석
Megalocytivirus
의sub-typing
에사용된표준검정시료는배 양된 바이러스(subgroup 2, IVS; subgroup 3, PGIV-K1)
과FLIV (subgroup 4)
감염넙치의비장으로부터추출된DNA
를 사용하였다.
그리고Megalocytivirus
양성패류의중장선으로 부터직접추출한DNA
또는2-step PCR
생성물로부터cloning
하여분리된plasmid DNA
를HRM template
로사용하였다.
HRM
을시행하기위한반응액에는10×buffer 2 μL, 200 μM
의각각의dNTP, 1 μM
의forward
와reverse primer, EvaGreen (Bioyium, Korea) 1 μL, Hot start Taq (HS prime Taq DNA polymerase, Genet Bio, Korea)
및template 1 μL
를첨가후증 류수로최종액의volume
이20 μL
가되도록했다. HRM
분석 은Rotor-Gene
TM6000 (Qiagen, Germany)
을사용하였고아래 와같은방법으로실시하였다.
먼저
95℃
에서5
분간pre-denaturation
시킨 후, 95℃
에서10
초denaturation, 56℃
에서10
초annealing, 72℃
에서15
초extension
의반응을45 cycle
수행하였다. 45 cycle
이끝난후80-90℃
에서0.1℃/s
의속도로온도를증가시키며Tm (melt- ing temperature)
값을측정하였다.
결 과
패류 중장선 내 Megalocytivirus의 검출
채집된패류각개체별로
(18
개체)
분리된DNA
를template
로하여Megalocytivirus
에대해PCR
을수행하였을때1-step PCR (35 cycles)
에서 모두 음성의 결과를 보였으나, 2-step PCR (35 cycles)
수행시진하굴시료에서50.0%,
고성굴시 료에서38.9%,
서산바지락시료에서39.0%
의시료에서280 bp
의amplicon
을생성하는양성률을보여지역간의뚜렷한차 이는나타나지않았다.
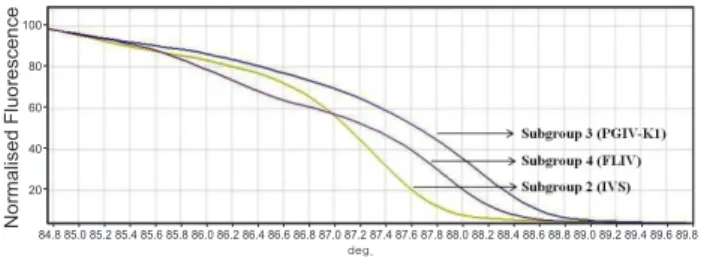
HRM 분석 기법의 최적화
HRM
분석기법의최적화를실시하기위해서Megalocytivi- rus
의subgroup 2 (IVS), subgroup 3 (PGIV-K1)
그리고sub-
group 4 (FLIV)
의각각에해당하는표준검정시료를대상으로HRM
을실시하였다.
현재subgroup 1
은우리나라,
일본중국 에서도발견되지않으며또한새롭게제시되고있는현재의분 류체계에서는subgroup 2
에포함하는경우가많으므로여기서 는따로구별하여분석을실시하지않았다HRM
을 수행한결과, Megalocytivirus
의 각subgroup
별로Tm
값이IVS
의경우87.35℃, PGIV-K1
의경우88.13℃, FLIV
의경우87.90℃
차이가나타났으며,
증폭된산물의염기서열 차이에따라서감소하는형광값이차이가났다.
그리고감소 하는형광값을normalized fluorescence
로변환하였을때sub- group
별로구별이가능하였다(Fig. 1).
HRM을 활용한 패류 내 존재하는 Megalocytivirus의 subgroup분석
Megalocytivirus
양성개체로부터분리된DNA
를직접tem- plate
으로하여HRM
을수행할경우PCR
증폭이이루어지지 않아melting-curve
의분석을할수없었다.
하지만2-step PCR
생성물로부터cloning
하여분리된plasmid DNA
를template
로 하여
HRM
을 실시하였을때 표준양성시료와동일한Tm
값을확인할수있었으며
subgroup
분석이가능하였다. HRM
을수행한모든양성시료에서Megalocytivirus subgroup 2
와subgroup 4
가존재하고있음을확인할수있었으며,
나타난subgroup
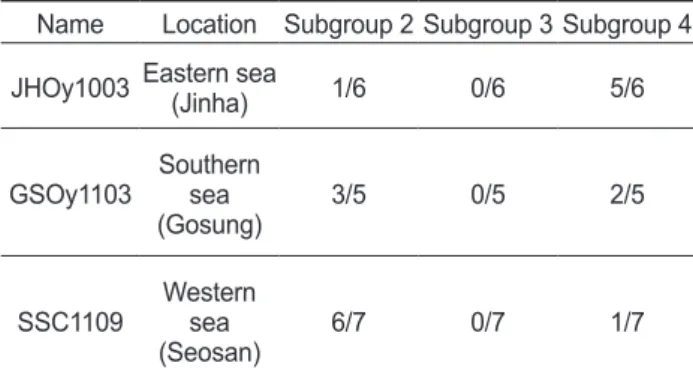
들의비율은시료별로차이가나고있었다. 2010
년3
월진하굴내의Megalocytiviruses (JHOy1003)
의경우분석 된6
개의clones
중5
개가subgroup 4, 1
개가subgroup 2
에속 하는것으로나타났으며, 2011
년3
월고성굴내의Megalocy- tiviruses (GSOy1103)
의경우분석된5
개의clones
중2
개가subgroup 4
에, 3
개가subgroup 2
로나타났다.
그리고2011
년9
월서산바지락내의Megalocytiviruses (SSC1109)
의경우분 석된7
개의clones
중1
개가subgroup 4, 6
개가subgroup 2
에 속하는것으로나타났다(Fig. 2, Table 2).
또한, HRM
분석에 사용된plasmid DNA
를염기서열분석을통해subgroup
을확100 90 80 70 60 50 40 30 20 10
Normalised Fluorescence
84.8 85.0 85.2 85.4 85.6 85.8 86.0 86.2 86.4 86.6 86.8 87.0 87.2 87.4 87.6 87.8 88.0 88.2 88.4 88.6 88.8 89.0 89.2 89.4 deg.
84.6
100 90 80 70 60 50 40 30 20 10
Normalised Fluorescence
84.8 85.0 85.2 85.4 85.6 85.8 86.0 86.2 86.4 86.6 86.8 87.0 87.2 87.4 87.6 87.8 88.0 88.2 88.4 88.6 88.8 89.0 89.2 89.4 89.6 deg.
100 90 80 70 60 50 40 30 20 10
Normalised Fluorescence
84.8 85.0 85.2 85.4 85.6 85.8 86.0 86.2 86.4 86.6 86.8 87.0 87.2 87.4 87.6 87.8 88.0 88.2 88.4 88.6 88.8 89.0 89.2 89.4 89.6 deg.
100 80 60 40
20
Normalised Fluorescence 84.8 85.0 85.2 85.4 85.6 85.8 86.0 86.2 86.4 86.6 86.8 87.0 87.2 87.4 87.6 87.8 88.0 88.2 88.4 88.6 88.8 89.0 89.2 89.4 89.6 89.8
deg.
Fig. 1. Genetic differentiation of the reference Megalocytiviruses using high-resolution melting (HRM) analysis.
김광일
ㆍ
진지웅ㆍ
김영철ㆍ
정현도244
인한결과
HRM
결과와일치하고있음을알수있었으며,
국내
Megalocytiviruses
에서는동일subgroup
내에서의유전적variation
이매우제한적으로일어나고있음을확인할수있었 다(Fig. 3).
고 찰
패류의여과섭식을통한먹이섭취과정중축적된
norovirus
등의human enteric virus
에대한분석방법은많은연구에의 해보고되고있으며,
수생동물에영향을줄수있는바이러스인
MABV, WSSV
의패류조직내에서의축적도보고되고있다
(Atmar et al., 1995; Gerba et al., 1978; Goyal et al., 1979;
Suzuki and Nojima, 1999; Vazquez-Boucard et al., 2010).
또 한Megalocytivirus
의패류내축적은추출방법론적인부분만 이루어져있고정확한감염의생태적상황,
그리고병원체에대 한빠르고신속한새로운유전적비교방법에대한결과는아직 미진한상태에있다(Kim et al., 2011).
현재까지감염어류를대상으로하여국내에서보고된
Mega-
locytivirus
의subgroup
은돌돔등의돔류에서보고된subgroup 2
와 넙치,
터봇에서 보고된subgroup 4
의 두 종류이며 아직subgroup 1
과subgroup 3
은보고되지않고있다.
따라서
feeding-activity
를가지는패류에오염된Megalocy-
tivirus
의유전적특성규명은주변수권에서의질병발생과의연관관계추적및어류양식산업에있어서본바이러스의위 험성재고측면에서매우의미있는정보라고할수있을것이 다
.
다만여기서명확히하여야할것은현재는subgroup 2
는subgroup 1
을포함한다는분류가일반적으로받아들여지고있으며
(Kurita and Nakajima, 2012),
또한현재까지한,
중,
일의 최근10
년간의보고에서subgroup 1
에대한질병보고가없 었으며(Imajoh et al., 2007) subgroup 1
과subgroup 2
의구별 은큰의미를부여하기어려우므로본연구에서subgroup 1
과subgroup 3
로나누어논하고있는Megalocytivirus
는실질적 인측면에서는새로운개념으로받아들여지고있는subgroup 3
라는하나의의미로이해되는것도가능할것으로추정된다
.
일반적으로패류의중장선에축적된바이러스particles
수는 감염어류에비해적고또한유전적으로 유사한다양한sub- group
의Megalocytivirus strains
가축적될수있기때문에특이 적인primer
를사용한PCR
에서나타난amplicon
을분석하는 (B) Subgroups of Megalocytiviruses detected in Oysters in Gos- eong (GSOy1103)100 90 80 70 60 50 40 30 20 10
Normalised Fluorescence
84.8 85.0 85.2 85.4 85.6 85.8 86.0 86.2 86.4 86.6 86.8 87.0 87.2 87.4 87.6 87.8 88.0 88.2 88.4 88.6 88.8 89.0 89.2 89.4 deg.
84.6
100 90 80 70 60 50 40 30 20 10
Normalised Fluorescence
84.8 85.0 85.2 85.4 85.6 85.8 86.0 86.2 86.4 86.6 86.8 87.0 87.2 87.4 87.6 87.8 88.0 88.2 88.4 88.6 88.8 89.0 89.2 89.4 89.6 deg.
100 90 80 70 60 50 40 30 20 10
Normalised Fluorescence
84.8 85.0 85.2 85.4 85.6 85.8 86.0 86.2 86.4 86.6 86.8 87.0 87.2 87.4 87.6 87.8 88.0 88.2 88.4 88.6 88.8 89.0 89.2 89.4 89.6 deg.
60 40
20
Normalised Fluorescence 84.8 85.0 85.2 85.4 85.6 85.8 86.0 86.2 86.4 86.6 86.8 87.0 87.2 87.4 87.6 87.8 88.0 88.2 88.4 88.6 88.8 89.0 89.2 89.4 89.6 89.8
deg.
(A) Subgroups of Megalocytiviruses detected in Oysters in Jinha (JHOy1003)
100 90 80 70 60 50 40 30 20 10
Normalised Fluorescence
84.8 85.0 85.2 85.4 85.6 85.8 86.0 86.2 86.4 86.6 86.8 87.0 87.2 87.4 87.6 87.8 88.0 88.2 88.4 88.6 88.8 89.0 89.2 89.4 deg.
84.6
100 90 80 70 60 50 40 30 20 10
Normalised Fluorescence
84.8 85.0 85.2 85.4 85.6 85.8 86.0 86.2 86.4 86.6 86.8 87.0 87.2 87.4 87.6 87.8 88.0 88.2 88.4 88.6 88.8 89.0 89.2 89.4 89.6 deg.
100 90 80 70 60 50 40 30 20 10
Normalised Fluorescence
84.8 85.0 85.2 85.4 85.6 85.8 86.0 86.2 86.4 86.6 86.8 87.0 87.2 87.4 87.6 87.8 88.0 88.2 88.4 88.6 88.8 89.0 89.2 89.4 89.6 deg.
40
20
Normalised Fluorescence 84.8 85.0 85.2 85.4 85.6 85.8 86.0 86.2 86.4 86.6 86.8 87.0 87.2 87.4 87.6 87.8 88.0 88.2 88.4 88.6 88.8 89.0 89.2 89.4 89.6 89.8
deg.
(C) Subgroups of Megalocytiviruses detected in Clams in Seosan (SSC1109)
100 90 80 70 60 50 40 30 20 10
Normalised Fluorescence
84.8 85.0 85.2 85.4 85.6 85.8 86.0 86.2 86.4 86.6 86.8 87.0 87.2 87.4 87.6 87.8 88.0 88.2 88.4 88.6 88.8 89.0 89.2 89.4 deg.
84.6
100 90 80 70 60 50 40 30 20 10
Normalised Fluorescence
84.8 85.0 85.2 85.4 85.6 85.8 86.0 86.2 86.4 86.6 86.8 87.0 87.2 87.4 87.6 87.8 88.0 88.2 88.4 88.6 88.8 89.0 89.2 89.4 89.6 deg.
100 90 80 70 60 50 40 30 20 10
Normalised Fluorescence
84.8 85.0 85.2 85.4 85.6 85.8 86.0 86.2 86.4 86.6 86.8 87.0 87.2 87.4 87.6 87.8 88.0 88.2 88.4 88.6 88.8 89.0 89.2 89.4 89.6 deg.
80 60 40
20
Normalised Fluorescence 84.8 85.0 85.2 85.4 85.6 85.8 86.0 86.2 86.4 86.6 86.8 87.0 87.2 87.4 87.6 87.8 88.0 88.2 88.4 88.6 88.8 89.0 89.2 89.4 89.6 89.8
deg.
Fig. 2. Differentiation of the Megalocytivirus subgroups using high-resolution melting (HRM) analysis. Amplicons of the MCP gene of Megalocytiviruses in 2-step PCR from shellfish were cloned. Each purified plasmid from 5~7 separated colonies grown on a single cloning plate were used for HRM analysis.
Table 2. Distribution of subgroups in Megalocytivirus obtained from shellfish based on differentiation by high-resolution melting analysis.
Name Location Subgroup 2 Subgroup 3 Subgroup 4 JHOy1003 Eastern sea(Jinha) 1/6 0/6 5/6
GSOy1103 Southern
(Gosung)sea 3/5 0/5 2/5
SSC1109 Western
(Seosan)sea 6/7 0/7 1/7
HRM분석법을 이용한 Megalocytivirus의 유전적 분석
245
것은그
virus
의다양성분석에는충분한의미를부여하지못한다
.
그러므로다양한viral strains
모두에서나타나는conserved genomic region
에대한primer
를사용한PCR
을실시한후생 성된amplicon
의유전적특성또는variation
을분석하면detec- tion
된strains
모두를분석할수있는것과같은의미를지니는 빠르고간편한방법의활용이필요하다.
HRM
은PCR
의증폭산물로부터target
이되는유전자의유 전적 다양성을 신속하고 효율적으로 확인할수 있는기술로mutant
나variation
부위를 확인할 수 있다(Kristensen and Dobrovic, 2008; Tajiri-Utagawa et al., 2008; Wittwer et al., 2003).
따라서본연구에서는Megalocytivirus
의MCP gene
을target
으로한HRM
기법을사용하여패류내에축적된Mega- locytivirus
의subgroup
의분석을유전자염기배열분석없이 보다빠르고,
간단하며,
편리하게실시하고자하였다.
일반적으로패류에축적된바이러스는양적 문제뿐만아니 라 다른 유전자형의 바이러스가동시에 존재하기 때문에조 직
DNA
를주형으로한PCR amplicon
에대한HRM
의직접 적인적용은힘들다.
따라서본연구에서는4
종의subgroup
의MCP
유전자부위에서conserved region
을target
으로한새로 운primer
를제작하고이를사용하여2-step PCR
을실시한후결과물로나타난
amplicon
을pGEM-T Easy vector
에cloning
하였다.
나타난clones
를무작위로선별하여plasmid
를분리한 후cloned MCP gene
에대한PCR amplicon
의variation
에대한HRM
분석을실시하여subgroup
을확인하였다.
그결과,
모든 양성시료에는subgroup 2
와subgroup 4
가함께존재하고있는 것을확인할수있었다.
그러나subgroup 1
과subgroup 3
는검 출되지않아아직까지의국내발병사례에서이두가지type
에 의한사례가보고된바없다는결과와일치함을알수있었다.
본연구에서비록subgroup 1
에대한HRM
은따로실시하지 않았으나gene sequence analysis
에서subgroup 1
로분류할 수있는clones
는없었다(Fig. 3).
그러므로우리나라의수권은 우리나라양식현장에서나타나는subgroup 2
와subgroup 4
두 가지types
에노출되어있으며,
아직subgroup 1
과subgroup 3
은우리나라양식현장의수권에서비상존성바이러스종(ex- otic species)
으로분류할수있음을확인할수있었다.
향후수 입수산물의검역에서이에대한보다정밀한분석이항상이루 어져새로운subgroup
의국내유입방지책이있어야할것이다.
더욱흥미로운 것은시료를 채취한지역별로 나타난
sub-
group
들의비율이차이가있다는것이었다. JHOy1003
로부터 의2-step PCR amplicon-clones
중에는subgroup 4
가83.3%
Fig. 3. Nucleotide sequence alignment of the MCP genes of Megalocytiviruses in shellfish. Each cloned plasmid was purified from colonies on plate used for cloning of MCP gene amplicons in 2-step PCR and used for determination of nucleotide sequence.
의비율로나타났고
, SSC1109
로부터의2-step PCR amplicon- clones
중에는subgroup 2
가85.7%
의비율을차지하고 있어 동해지역(
진하)
은subgroup 4,
서해지역(
서산)
은subgroup 2
가 각각우점적임을확인할수있었다.
그리고GSOy1103
로부 터의2-step PCR amplicon-clones
중에는subgroup 2
가60%, subgroup 4
가40%
로나타나남해지역(
고성)
은subgroup 2
와subgroup 4
가서로유사한비율로나타나고있음을확인할수있었다
.
이러한결과는주변양식환경에따라넙치양식이주를이루고있는동해지역의경우넙치에감염되는
FLIV
가굴에축적되어서나타난다고생각되어지며또한동해지역과는달 리남해지역과서해지역의경우넙치양식뿐만아니라돌돔 양식을포함하여다양한어종의양식이이루어지고있으므로
subgroup 2
와subgroup 4
에속하는바이러스가매년어류에서 발생하여수권으로유출되어패류내로축적되고있는것으로 확인되어졌다.
기법적으로보면염기서열분석결과와본연구 의HRM
결과와비교하였을때결정된subgroup
들은서로동 일한결과를보였으며(Fig. 3),
이러한결과를통하여Megalo- cytivirus
의subgroup
분석에있어서HRM
분석기법이기존의 염기서열분석법보다신속하고효율적임을확인할수있었다.
결론적으로 국내에 서식하고 있는 패류의 중장선 내에서
Megalocytivirus
의오염존재의확인과함께subgroup
의결정 을위한HRM
분석방법을제안할수있었으며, Megalocytivi- rus
양성패류의중장선내에는subgroup 2, subgroup 4
가존재 하고있음을확인할수있었다.
향후패류및어류내감염바이러스의진단및유전적분석을위하여본연구의
HRM
기법의응용은한단계진보된기법임을확인하였다
.
사 사
이논문은부경대학교자율창의학술연구비
(2013
년)
의 지원 에의해연구되었습니다.
References
A
tmar RL, Neill FH, Romalde JL, Le Guyader F, Woodley CM, Metcalf TG and Estes MK. 1995. Detection of Norwalk vi- rus and hepatitis A viruses in shellfish tissues with the PCR.Appl Environ Microbiol 61, 3014-3018.
Do JW, Cha SJ, Kim JS, Jeong EJ, An EJ, Lee NS, Choi HJ, Lee CH, Park MS, Kim JW, Kim YC and Park JW. 2005.
Phylogenetic analysis of the major capsid protein gene of iri- dovirus isolates from cultured flounders Paralichthys oliva-
ceus in Korea. Dis Aquat Organ 64, 193-200. http://dx.doi.
org/10.3354/dao064193.
Gerba CP and Goyal SM. 1978. Detection and occurrence of en- teric viruses in shellfish: A review. J Food Prot 41, 743-754.
Goyal SM, Gerba CP and Melnick JL. 1979. Human enterovi- ruses in oysters and their overlying waters. Appl Environ Microbiol 37, 572-575.
Inouye K, Yamano K, Maeno Y, Nakajima K, Matsuoka M, Wada Y and Sorimachi M. 1992. Iridovirus infection of cul- tured red sea bream, Pagrus major. Fish Pathol 27, 19-27.
Imajoh M, Ikawa T and Oshima SI. 2007. Characterization of a new fibroblast cell line from a tail fin of red sea bream,
Pagrus major, and phylogenetic relationships of a recent
RSIV isolate in Japan. Virus Res 126, 45-52. http://dx.doi.org/10.1016/j.virusres.2006.12.020.
Jeong JB, Jun LJ, Yoo MH, Kim MS, Komisar JL and Jeong HD. 2003. Characterization of the DNA nucleotide sequenc- es in the genome of red sea bream iridoviruses isolated in Korea. Aquaculture 220, 119-133.
Jeong JB, Kim HY, Jun LJ, Lyu JH, Park NG, Kim JK and Jeong HD. 2008. Outbreaks and risks of infectious spleen and kid- ney necrosis virus disease in freshwater ornamental fishes.
Dis Aquat Organ 78, 209-215
Kim JW, Cho MY, Jin JW, Kim KH, Kim KI and Jeong HD.
2011. Detection of Megalocytivirus in shellfish using PCR with various DNA extraction methods. J Fish Pathol 24, 65- Kurita J and Nakajima K. 2012. Megalocytiviruses. Viruses 4, 73.
521-538.
Kristensen LS and Dobrovic A. 2008. Direct genotyping of sin- gle nucleotide polymorphisms in methyl metabolism genes using probe-free high-resolution melting analysis. Cancer Epidemiol Biomarkers Prev 17, 1240-1247. http://dx.doi.
org/10.1158/1055-9965.EPI-07-2531.
Oh MJ, Kitamura SI, Kim WS, Park MK, Jung SJ, Miyadai T and Ohtani M. 2006. Susceptibility of marine fish species to a Megalocytivirus, turbot iridovirus, isolated from turbot,
Psetta maximus (L.). J Fish Dis 29, 415-421.
Suzuki S and Nojima M. 1999. Detection of a marine birnavirus in wild molluscan shellfish species from Japan. Fish Pathol 34, 121-125.
Tajiri-Utagawa E, Hara M, Takahashi K, Watanabe M and Wakita T. 2008. Development of a rapid high-throughput method for high-resolution melting analysis for routine de- tection and genotyping of noroviruses. J Clin Microbiol 47, 435-440. http://dx.doi.org/10.1128/JCM.01247-08.
Vazquez-Boucard C, Alvarez-Ruiz P, Escobedo-Fregoso C, Anguiano-Vega G, Duran-Avelar Mde J, Pinto VS and Esc- obedo-Bonilla CM. 2010. Detection of white spot syndrome virus (WSSV) in the Pacific oyster Crassostrea gigas. J In- vertebr Pathol 104, 245-247.
Wittwer CT, Reed GH, Gundry CN, Vandersteen JG and Pryor RJ. 2003. High-resolution genotyping by amplicon melting analysis using LCGreen. Am Assoc Clin Chem 49, 853-860.