ISSR 표지에 의한 연속 ( Nelumbo )의 유연관계 분석
류재혁*·최갑림**·류재일***·이성춘*·천종은*·신동영*·배창휴*
†*순천대학교 바이오자원학부, **목원대학교 창업보육센터, ***한국생명공학연구원
Genetic Relationship Analysis of genus Nelumbo Accessions Based on Inter-Simple Sequence Repeats (ISSR)
JaiHyunk Ryu * , Gab Lim Choi ** , Jae Il Lyu *** , Sheong Chun Lee * , Jong Un Chun * , Dong Young Shin * and Chang-Hyu Bae *†
*Department of Bioresources Science, Sunchon National University, Suncheon, 540-742, Korea.
**Business Incubation Center, Mokwon University, Daejeon 302-318, Korea.
***Division of Biosystems Reserch, KRIBB, Daejeon 305-806, Korea.
ABSTRACT : The polymorphism and the genetic relationships among 32 genetic resources of genus Nelumbo from Korea, Japan, China, USA, India, Thailand and Gabong were thoroughly investigated and extensively examined using ISSR mark- ers. Out of 103 loci detected overall, 94 were identified to be polymorphic with a rate of 91.2%. The genetic similarity matrix revealed a wide range of variability among the 32 accessions, spanning from 0.227 to 0.833. The study findings indicate that the Nelumbo accessions have a high genetic diversity, and accordingly carry a germplasm qualifying as good genetic resources for cross breeding. According to the clustering analysis, different subspecies, N. nucifera and N. lutea , were divided into independent groups and all of the N. nucifera accessions could be classified into five categories. Compared to RAPD analysis, ISSR method showed a clearer picture of polymorphism among the accessions and exhibited a definite distinction even among the subspecies. In this respect, ISSR analysis is considered to be more effective in differentiating the accessions and subspecies of the genus Nelumbo than RAPD test.
Key Words : Nelumbo nucifera , ISSR, Cluster Analysis, Genetic Similarity
서 언
연
( Nelumbo nucifera Gaertn.)
은 부엽식물에 속하는 수생식물로주로인도와중국을중심으로열대
,
온대지방에널리분포하며 연못에서 자라고 논밭에서 재배하며
(Borsch and
Bartholott, 1994; Dahlgrea and Rasmussen, 1983)
관상용,
약용
,
식용으로 널리 이용되고있다.
특히 괴경의끝 부분이굵어져 식용으로 사용하는 연근을 우절
(Nelumbinis
Rhizomatis Nodus)
이라하여 지혈제(
止血劑)
로사용되며,
종 자는연자육(Nelumbo Seed)
이라 하여유정(
遺精),
대하(
帶下
)
등의 치료에 사용하고 있다(Mukherjee et al ., 1997;
Lee et al., 2006a; Lee et al ., 2006b).
연속은수련과
(Nymphaeaceae)
에포함되어통용되어왔으나 최근 분자계통분석에 근거하여(Barkman et al ., 2000; Lee and Wen, 2004)
연과(Nelumbonaceae)
연속으로 분류되며,
연속은 북미 원산종
( N. lutea )
과 동아시아 원산종( N.
nucifera )
이있다(Borsch and Barthlott, 1994).
ISSR
은 단순한 반복 서열에 의한 분자 마커로 기능은 알수없지만반복서열이게놈에위치하여종이나집단에특이적 으로 판정가능하며
(Fang and Roose, 1997), SSR
마커와는 달리목표로하는 사전염기서열의지식이필요하지않아쉽 게이용가능하다(Tsumura et al ., 1996).
또한ISSR
분석은 프라이머의높은결합 온도와긴염기서열때문에RAPD
방 법 보다 정확하고 재현성이 우수하다(Esselman et al .,
1999). ISSR
이 목표로 하는 반복 서열은 진핵 게놈 전체에걸쳐 풍부하지만 빠르게 진화하는 것으로 알려져 있다
(Godwin et al ., 1997).
따라서ISSR
분석은 집단유전 연구,
특히유연관계가근연한종이나집단의유전적다양성연구에 유용한방법이다
(Russel et al ., 1997).
ISSR
마커를이용한연의유전분석에관한연구는중국및†
Corresponding author: (Phone) +82-61-750-3214 (E-mail) [email protected]
Received 2010 February 9 / 1st Revised 2010 March 5 / 2nd Revised 2010 March 29 / Accepted 2010 April 5
연속의 ISSR 분석
87 미국 품종과 교배품종의 유전적 다양성 분석
(Chen et al ., 2008),
영양번식,
종자번식으로 육성된중국내연품종의유연관계 분석
(Tian et al ., 2008),
중국의 북부지방과 중부지방 야생종의 계통의 유전적 다양성 분석(Xue et al ., 2006;
Han et al ., 2007b)
등다양한 연구가진행되었다.
국내의경우
RAPD
방법에따른연품종및수집종간지역적분화양상에 대한 연구가 이루어진 바 있으나
(Kim et al., 1998)
ISSR
마커를이용한수집계통의유전적다양성과유연관계분석은미진한실정이다
.
따라서본연구는연의신품종육종에필요한기초자료를제공할목적으로국내수집종과외국에서
도입되어국내에서재배되고있는품종을대상으로
ISSR
을분석하여유전적유사도를평가하고
,
국내수집종과외래도입종 간의유연관계를분석하고자수행되었다.
재료 및 방법
1. 식물재료
본연구에서는 국내 수집종
(7
계통),
일본 도입종(5
계통),
중국 도입종
(9
계통),
미국 도입종(8
계통),
인도 도입종(1
계 통),
태국도입종(1
계통),
아프리카가봉도입종(1
계통)
을포 함한총32
계통을공시하였다(Table 1).
국내 수집종은해남(
인취사),
강진(
금당지),
전주(
다가동련),
무안(
회산지),
대구(
안심습지),
시흥(
관곡지),
양양(
양양지)
등자생지에서직접수집하였고
,
도입종과함께실험포장용기(45 × 90
㎝)
에이식 하여실험재료로사용하였다(Choi, 2009).
2. DNA 추출
DNA
는CTAB
방법(Fang et al ., 1992)
에 따라 추출하여 Table 1.The origin of the N. nucifera and N. lutea accessions used in this study.
No. subspecies Cultivars name or locality
†Collected region Origin
1 N. nucifera subsp. nucifera Bailian Jeonnam, Gokseong China
2 N. nucifera subsp. nucifera CiLian Jeonnam, Gokseong China
3 N. nucifera subsp. nucifera 'Inchisa' Jeonnam, Heanam Korea
4 N. nucifera subsp. nucifera Jinbihuihuang Jeonnam, Gokseong China
5 N. nucifera subsp. nucifera Redscarf Jeonnam, Gokseong USA
6 N. nucifera subsp. nucifera GuokLian Gyeonggi, Icheon China
7 N. nucifera subsp. lutea Lutia Gyeonggi, Icheon USA
8 N. nucifera subsp. nucifera First Lady Gyeonggi, Icheon USA
9 N. nucifera subsp. nucifera Perry D. Slocum Gyeonggi, Icheon USA
10 N. nucifera subsp. nucifera Giant Sunburst Gyeonggi, Icheon USA
11 N. nucifera subsp. nucifera Charles Tomas Gyeonggi, Icheon USA
12 N. nucifera subsp. nucifera 'kumdangji' Gyeonggi, Icheon Korea
13 N. nucifera subsp. nucifera 'Dagadonglian' Gyeonggi, Icheon Korea
14 N. nucifera subsp. nucifera Perry R..Slocum Gyeonggi, Icheon USA
15 N. nucifera subsp. nucifera 'Gwangokji' Gyeonggi, Siheung Korea
16 N. nucifera subsp. nucifera 'Yangyangji' Gangwon, Yangyang Korea
17 N. nucifera subsp. nucifera 'whasanji' Jeonnam, Muan Korea
18 N. nucifera subsp. nucifera 'Ansimji' Deagu, Ansimdong Korea
19 N. nucifera subsp. nucifera Daihe Gyeonggi Uiwang Japan
20 N. nucifera subsp. nucifera Zhenru Gyeonggi, Uiwang Japan
21 N. nucifera subsp. nucifera JinLun Gyeonggi, Uiwang Japan
22 N. nucifera subsp. nucifera Baiqunzi Gyeonggi, Uiwang Japan
23 N. nucifera subsp. nucifera kyeongbokgungLian Gyeonggi, Uiwang Japan
24 N. nucifera subsp. nucifera Gabong Gyeonggi, Uiwang Gabong
25 N. nucifera subsp. nucifera Daisuijin Gyeonggi, Uiwang China
26 N. nucifera subsp. nucifera Sharon Gyeonggi, Uiwang USA
27 N. nucifera subsp. nucifera Xuanwulian Gyeonggi, Uiwang China
28 N. nucifera subsp. nucifera Shenshuihonglian Gyeonggi, Uiwang China
29 N. nucifera subsp. nucifera Ziyuolian Gyeonggi, Uiwang China
30 N. nucifera subsp. nucifera Chewanbashou Gyeonggi, Uiwang China
31 N. nucifera subsp. nucifera IndiaLian Gyeonggi, Uiwang India
32 N. nucifera subsp. nucifera Taegoklian Gyeonggi, Uiwang Thailand
†
The name of this column is used to represent Nelumbo cultivars (without apostrophe) or locality of samples (with apostrophe).
agarose gel (0.8%)
에서전기영동후밴드를확인하고, Nanodrop Spectrophotometer (Thermo Fisher Scientific, Netherlands)
를이용하여농도를측정하였다
.
3. ISSR 분석
국내 수집종과 도입종간의 유연관계 분석을 위해
20
개의ISSR
프라이머(UBC primer Set No. 9, University of British Columbia, Canada)
로예비실험후밴드가명확하고다형성이우수한
10
개의프라이머를선택하여분석에사용하였다(Table 2). PCR
반응액의 조성은총25
㎕로주형DNA 100 ng/
㎕, Taq polymerase 1 unit, 10 × PCR reaction buffer (10 mM Tris-HCl pH 8.3; 50 mM KCl; 1.5 mM MgCl
2), 0.2 mM dNTP, 10 pmol
프라이머를혼합하여사용하였다. PCR
반응은Gene Amp 2700 (Applied Biosystem)
을 사용하여94
℃에서5
분간초기변성시킨후94
℃에서1
분간predenaturation, 55
℃에서
1
분간annealing, 72
℃에서2
분간extension
을39
회 수 행한후최종적으로72
℃에서8
분간반응시킨후종료하였다. PCR
완료 후 증폭산물15
㎕ 를agarose gel (1.5%)
에loading
하여 전기영동(50 V, 60
분간)
한 후EtBr
로 염색, Gel Doc XR (Bio-rad, USA)
을사용하여주요밴드를확인하였다.
4. Data 분석
ISSR
로분석된DNA
밴드양상은이진수를이용하여밴드유·무에 따라
1
과0
으로data
를 표시하여 통계 프로그램[SPSS V.12 (SPSS Inc., USA)]
에 입력하였고, Jaccard
계수(Jaccard coefficient)
로유전적유사도지수를 산출하고 평균연 결법(Average Linkage Method)
으로군집분석을실시하였다.
결과 및 고찰
1. 연 수집계통 간의 DNA 밴드 양상
10
개의 프라이머를이용하여국내 수집종(7
계통)
및도입종
(25
종)
의ISSR
분석결과는다음과같다.
20
개의ISSR
프라이머중재현성이높은10
개의프라이머(Table 2)
를사용한결과,
연의 수집 계통간증폭된DNA
산 물의 각프라이머별총밴드 수는4
개(UBC 841)
에서15
개(UBC 835)
로한프라이머조합 당평균 밴드수는10.3
개였 다.
다형성 밴드 수는 최소3
개(UBC 841)
에서 최대13
개(UBC 854, UBC 835)
로평균9.4
개의 다형성밴드가검출되었다
.
이는동일한수집종에대하여RAPD
분석결과를통해얻어진다형성밴드수가평균
9.66
개로Choi (2009)
의결과와유사하며
,
중국중부지방야생종N. nucifera (89
계통)
에대한ISSR
분석으로 검출된 프라이머 당 평균 다형성 밴드수인12.33
개보다낮은수준이었다(Han et al ., 2007a).
연속 품종간에서 검출된 특이적 증폭 산물의 경우
UBC 827
프라이머[(AC)
8G]
에서N. lutea
품종의 특이 밴드3
개 가 증폭되었으며(Fig. 1-A), UBC 835
프라이머[(AG)
8YC)]
에서는
N. nucifera
품종특이적증폭산물1
개가검출되었다(Fig. 1-B).
이러한 결과는Tian
등(2008)
이연 품종103
계 통의ISSR
분석에서2
개의N. lutea
품종 특이적인 밴드를 검출하고, N. nucifera
품종특이적밴드를 검출한결과와유 사한빈도이다.
수집종간밴드의다형성비율은
Table 2
에서와같이증폭된총
103
개의 밴드 중다형성 밴드가94
개로91.2%
의다형성을나타내었다
.
이러한결과는본연구에서사용한연수집계통과 동일한 재료로
RAPD
분석을 하였을때 총68
개밴드중
58
개의 다형성 밴드가 검출되어85.2%
다형을 나타낸Choi (2009)
의결과보다높은다형성을나타낸결과이다.
이 는 국화(Wolff et al ., 1995),
가시오갈피(Hong et al ., 2000),
동과(Verma et al ., 2007)
등 다른 약용작물에서ISSR
분석이RAPD
방법보다높은다형성을나타낸연구결과와유사하다
.
여러국가에서수집된연속품종에대한
ISSR
분석결과다형성 비율이
3
개국(
중국,
미국,
일본)
의38
개 수집종에서 Table 2.Sequences of the ISSR primers, number of total bands and number of polymorphic bands amplified by each primer across all 32
accessions.
Primer Sequence G+C (%) No. of total band No. of polymorphism band Polymorphism rate (%)
UBC811 (GA)
8C 52.9 11 10 90.9
UBC824 (TC)
8G 52.9 13 13 100
UBC827 (AC)
8G 52.9 12 10 83.3
UBC835 (AG)
8YC 50.0 12 12 100
UBC841 (GA)
8YC 50.0 4 3 75
UBC843 (CT)
8RA 44.4 9 8 88.8
UBC856 (AC)
8YA 44.4 11 11 100
UBC862 (AGC)
566.7 10 9 90
UBC866 (CTC)
566.7 11 9 81.8
UBC889 DBD(AC)
741.2 10 9 90
Total 103 94 91.2%
†
B: (C, G, T), D: (A, G, T), R: (A, G), Y: (C, T)
연속의 ISSR 분석
89
76.8% (Han et al ., 2007a), 2
개국(
중국,
미국) 92
품종에서55.8% (Chen et al ., 2008),
그리고중국내103
개수집종에서 는37.63% (Tian et al ., 2008)
로 원거리에서수집된 수집종일수록유전적 다형성이높게 나타났다
.
또한 중국 중부지방야생종 연속
89
계통에 대하여ISSR
분석 결과 다형성이90.2%
로 높게 나타났다(Han et al ., 2007b).
이러한 연구결 과는근거리수집종간보다는원거리에서수집된집단일수록 변이가많다는 것을 보여준다.
연속의 유전적다형성은백악 기부터지역별로축적된다양한변이가오랜기간에걸쳐선 발육종되어확립된지역종들이각각원거리에위치하여높은 유전적다형성을갖는 것으로유추 된다(Han et al ., 2007b;
Chen et al ., 2008; Tian et al ., 2008).
따라서 본연구 결과 높은다형성은지리적으로원거리에있는수집종들간에축적 된변이에기인한것으로사료된다.
2. 유전적 유사도 지수와 군집분석
연의국내수집종과도입종유전자원을육종적측면에서활
용하고자유전적유사도지수를산출하고
(Table 3)
군집분석을실시하였다
(Fig. 2).
총
32
계통간의 유전적유사도지수는 최저0.227 [7
번(
미 국품종), 30
번(
중국품종)]
에서 최고0.833 [25
번(
중국품종),
27 (
중국품종)]
이었고, 32
개 전체종의 유전적 유사도 평균은0.530
이었다.
이러한결과는본연구와같은국내수집종들을RAPD
분석했을 때 나타난 유전적 유사도 지수인0.750~
0.932 (Choi, 2009)
보다 폭넓게 나타난 결과이다.
또한Kim
등
(1998)
이국내 수집종(36
계통)
을대상으로RAPD
결과를분석한유전적 유사도지수인
0.785~1.000
과Han
등(2007
a)
이
ISSR
을이용하여중국,
일본,
미국의연속수집종(38
계통)
에서산출한유전적유사도지수인
0.785~0.968
에비하여폭넓은 수준의 유전적 유사도를 나타낸 것이다
.
한편Kwon
등(1998)
이부추의 원연교배육종재료로서 활용 가능한 유전적유사도 지수로 보고한
0.231~0.713
과유사한 결과이다.
이와같이국내 수집종과도입종사이에넓은 범위의유전적유사 도지수를보여유전적다양성이높은것으로나타났기때문 에 유사도지수가 낮은 계통들은교배 육종에서육종소재로 활용이 가능하다고 판단된다
(Baigalmaa et al ., 2009; Kim et al ., 2008; Kwon et al., 1998; Lyu et al ., 2003).
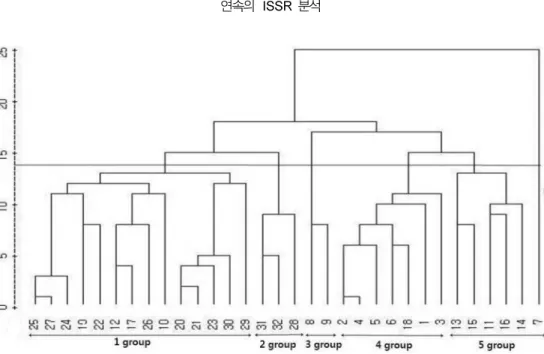
평균연결법
(Average Linkage Method)
으로 덴드로그램을 작성하여32
개수집종을군집분석한결과(Fig. 2),
유전적유사도 지수
0.352
에서N. nucifera
품종전체와N. lutea
품종 이나뉘어졌으며,
유전적유사도지수0.592
에서총수집계통 은5
개의N. nucifera
그룹과1
개의N. lutea
그룹으로 나눠 Fig. 1.ISSR electrophoresis profiles for 32 accessions of
N. nuciferaand
N. luteashowing
N. nuciferadeficient or additive DNA
bands (arrows of plate A and B). Amplification products were separated on agarose gels (1.5%) in 1xTAE buffer and
photographed using gel documentation system (Bio-rad, USA). Lane M : 100 bp marker, lanes 1~32 : accessions are
included in Table 1.
연속 의 IS SR 분석 90
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32
1 1.000
2 0.729 1.000
3 0.534 0.672 1.000
4 0.672 0.833 0.764 1.000
5 0.656 0.730 0.600 0.730 1.000
6 0.591 0.712 0.587 0.712 0.723 1.000
7 0.338 0.388 0.361 0.354 0.392 0.373 1.000
8 0.500 0.533 0.681 0.586 0.542 0.532 0.409 1.000
9 0.537 0.492 0.560 0.492 0.525 0.446 0.415 0.698 1.000
10 0.556 0.438 0.411 0.415 0.422 0.397 0.329 0.451 0.460 1.000
11 0.530 0.557 0.524 0.580 0.612 0.623 0.392 0.596 0.475 0.400 1.000
12 0.525 0.464 0.443 0.464 0.515 0.444 0.342 0.456 0.414 0.627 0.613 1.000
13 0.561 0.586 0.485 0.586 0.618 0.562 0.493 0.576 0.533 0.431 0.692 0.500 1.000
14 0.458 0.527 0.493 0.569 0.623 0.568 0.443 0.439 0.446 0.397 0.647 0.529 0.520 1.000
15 0.485 0.536 0.453 0.514 0.522 0.473 0.446 0.517 0.475 0.517 0.615 0.590 0.698 0.493 1.000
16 0.536 0.606 0.507 0.629 0.614 0.603 0.386 0.500 0.462 0.391 0.662 0.479 0.691 0.648 0.571 1.000
17 0.548 0.529 0.422 0.486 0.515 0.486 0.364 0.410 0.371 0.623 0.561 0.759 0.479 0.551 0.587 0.459 1.000
18 0.609 0.682 0.607 0.682 0.667 0.727 0.366 0.500 0.438 0.453 0.618 0.500 0.577 0.676 0.574 0.691 0.544 1.000
19 0.632 0.554 0.492 0.554 0.538 0.529 0.325 0.509 0.464 0.566 0.538 0.614 0.500 0.507 0.644 0.458 0.667 0.619 1.000
20 0.475 0.439 0.464 0.418 0.446 0.485 0.297 0.481 0.434 0.604 0.492 0.686 0.391 0.400 0.492 0.356 0.618 0.477 0.654 1.000
21 0.473 0.413 0.434 0.391 0.419 0.460 0.268 0.479 0.429 0.578 0.443 0.600 0.364 0.394 0.417 0.348 0.566 0.475 0.538 0.805 1.000
22 0.586 0.471 0.450 0.493 0.478 0.515 0.278 0.519 0.473 0.640 0.523 0.625 0.507 0.411 0.548 0.465 0.593 0.530 0.685 0.700 0.580 1.000
23 0.517 0.478 0.483 0.478 0.508 0.545 0.316 0.528 0.509 0.528 0.531 0.636 0.449 0.457 0.508 0.431 0.550 0.538 0.636 0.750 0.773 0.618 1.000
24 0.469 0.478 0.435 0.457 0.485 0.522 0.355 0.500 0.456 0.615 0.507 0.576 0.537 0.438 0.661 0.493 0.574 0.537 0.550 0.611 0.558 0.673 0.625 1.000
25 0.500 0.576 0.492 0.529 0.537 0.597 0.382 0.536 0.466 0.593 0.585 0.583 0.544 0.507 0.667 0.543 0.607 0.615 0.610 0.648 0.566 0.679 0.632 0.811 1.000
26 0.525 0.485 0.419 0.443 0.515 0.486 0.325 0.431 0.464 0.627 0.471 0.736 0.437 0.465 0.540 0.400 0.638 0.500 0.614 0.720 0.633 0.596 0.698 0.632 0.583 1.000
27 0.516 0.544 0.460 0.500 0.529 0.612 0.359 0.475 0.433 0.611 0.600 0.574 0.536 0.543 0.629 0.514 0.597 0.631 0.627 0.667 0.556 0.667 0.649 0.764 0.833 0.600 1.000
28 0.547 0.529 0.469 0.551 0.536 0.594 0.333 0.435 0.443 0.459 0.631 0.531 0.543 0.571 0.515 0.542 0.507 0.636 0.607 0.533 0.509 0.540 0.627 0.478 0.554 0.581 0.569 1.000
29 0.473 0.435 0.434 0.413 0.467 0.438 0.286 0.479 0.556 0.543 0.492 0.538 0.429 0.460 0.491 0.409 0.456 0.500 0.569 0.609 0.581 0.549 0.660 0.500 0.566 0.600 0.556 0.593 1.000
30 0.482 0.422 0.472 0.422 0.406 0.446 0.227 0.490 0.412 0.553 0.500 0.608 0.373 0.424 0.450 0.397 0.545 0.508 0.577 0.727 0.795 0.588 0.739 0.509 0.574 0.608 0.536 0.571 0.628 1.000
31 0.484 0.449 0.475 0.493 0.456 0.537 0.278 0.491 0.397 0.464 0.623 0.542 0.554 0.515 0.500 0.529 0.516 0.603 0.596 0.635 0.646 0.607 0.648 0.533 0.567 0.569 0.610 0.702 0.519 0.653 1.000
연속의 ISSR 분석
91 졌다
. 1
그룹에는 전체 일본5
계통(No. 19, 20, 21, 22, 23)
과 중국4
계통(No. 25 27, 29, 30),
국내 수집2
계통(No. 12, 17),
미국2
계통(No. 10, 26),
가봉1
계통(No.
24)
이 포함되었고, 2
그룹은 인도(No. 31),
태국(No. 32),
중국
(No. 28)
이각각1
계통씩속하였으며, 3
그룹은 미국2
계통
(No. 8, 9)
이포함되었다. 4
그룹은중국4
계통(No. 1, 2, 4, 6),
국내 수집2
계통(No. 3, 18),
미국1
계통(No. 5)
이포함되었다
. 5
그룹은 국내 수집3
계통(No. 13, 15, 16)
과미국
2
계통(No. 11, 14)
이포함되었다.
수집된 국가별로 보면일본에서도입한5
개계통은모두1
그룹에속하고,
중국도입
9
계통은1
그룹과4
그룹에 각각 군집을 이루었다.
그러나국내 수집
7
계통은1
그룹에3
개, 4
그룹에는2
개, 5
그룹에
3
개씩각각분포하여군집을이루었다.
또한미국8
품 종중 N. nucifera7
계통은 각각1, 3, 4, 5
그룹에 나뉘어군 집을형성하였다.
N. lutea는 N. nucifera품종과같은 그룹을이루지 않았으며
, Choi (2009)
가 본 연구와 동일한 시료로RAPD
에따른군집분석의결과로나타난유전적유사도지수0.90
에서총32
수집계통이6
그룹으로분류된것과는일치하지않았다
.
이는RAPD
결과에서(Choi, 2009)
아종이 다른 N.lutea와 N. nucifera 품종으로 구분되지 않았으나 본 연구의
ISSR
분석에서는명확하게구분되었기때문이다.
따라서연속 의분자표지를이용한 분류방법은RAPD
분석 보다는ISSR
분석이효과적인 것으로판단되었다
.
또한본연구에서같은 그룹으로 분류된 중국4
계통(No. 25 27, 29, 30)
과 일본3
계통
(No. 19, 21, 22),
미국2
계통(No. 10, 26)
과국내 수 집2
계통(No. 12, 17),
그리고 미국2
계통(No. 8, 9)
이RAPD
분석에서도단일그룹에속한것은일치하였다.
이상의 결과와같이 본연구에서 수집된 N. nucifera 종은 유전적 다형성
(polymorphism)
이91.2%
로높았으며 넓은범 위의유전적유사도지수를보인 것으로보아유전적으로매 우다양한 것으로나타났다.
이러한유전적 다양성에기초하여신품종육종을위한육종소재로활용이가능할것으로사 료된다
.
또한동일한연수집계통을재료로RAPD
분석한경 우(Choi, 2009)
보다ISSR
분석에서높은 다형성을 나타내 고, RAPD
에서 아종이 다른 N. lutea와 N. nucifera 간은구분되지 않았으나
ISSR
분석에서는 명확하게 구분되었으므로RAPD
분석에비하여ISSR
분석이보다효과적인것으로판단되었다
.
감사의 글
이논문은
2005
년순천대학교학술연구비공모과제로 수행된연구결과의일부이며연구비지원에감사드립니다
.
LITERATURE CITED
Baigalmaa J, Kim MK, Noh JH, Sun H and Yang DC. (2009).
Phylogenetic analysis of schizonepeta spike on the basis of DNA sequences. Korean Journal of Medicinal Crop Science.
17:46-53.
Barkman TJ, Chenery G, McNeal JR, Lyons-Weiler J, Ellisens WJ, Moore G, Wolfe AD and Pamphillis CW. (2000).
Independent and combined analyses of sequences from all three genomic compartments converge on the root of flowering plant phylogeny. Proceedings of the National Academy of Sciences.
97:13166-13171.
Fig. 2.