313 책임저자:김영민, 305-811, 대전시 유성구 전민동 461-6
한남대학교 생명나노과학대학 생명과학과 Tel: 042-629-8753, Fax: 042-629-8751 E-mail: [email protected]
접수일:2010년 12월 3일, 1차 수정일:2010년 12월 7일, 2차 수정일:2010년 12월 9일, 게재승인일:2010년 12월 13일
Correspondence to:Young Min Kim
Department of Biological Sciences, College of Life Science and Nano Technology, Hannam University, 461-6, Jeonmin-dong, Yuseong-gu, Daejeon 305-811, Korea
Tel: +82-42-629-8753, Fax: +82-42-629-8751 E-mail: [email protected]
B16F10 피부암 세포에서 AMPK-VASP 신호경로를 경유한 셀레늄의 Apoptosis 효과
한남대학교 생명나노과학대학 1생명과학과, 2식품영양학과
박송이1ㆍ이솔화1ㆍ박옥진2ㆍ김영민1
Apoptotic Effects of Selenium via AMPK-VASP Signal Pathway in B16F10 Melanoma Cells
Song Yi Park1, Sol Hwa Lee1, Ock Jin Park2 and Young Min Kim1
Departments of 1Biological Sciences, 2Food and Nutrition, College of Life Science and Nano Technology, Hannam University, Daejeon 305-811, Korea
A vasodilatator-stimulierte phosphoprotein (VASP), Ena/VASP family, is connected to actin. It is associated with the signal pathway involving the proliferation and migration of cells, and VASP regulates cells-cells contacts. VASP in the blood acts as cAMP-and cGMP-dependent protein kinase. According to the positions of amino acid at Ser-157, Ser-239 and Thr-278, it is regulated by AMP-activated protein kinase (AMPK) that acts as Ser-Thr kinase. Recently the VASP is known to the new substrate of AMPK.
The AMPK plays a role of the energy sensor. If AMPK is activated in cancer cells, apoptosis is induced in them. However, activation of mTOR is known to be promoted cancer growth. In addition, Akt, which is a kind of Ser-Thr kinase, is responsible for proliferation, differentiation, and growth of cells, and it inhibits apoptosis in cancer cells and is known to engage in angiogenesis and metastasis. In this study, when selenium is treated in B16F10 cancer cells, we suggest that the growth of B16F10 cells is inhibited.
We determined with MTT assay, Fluorescence-Activated Cell Sorting (FACS), and Western blotting.
Finally, it is observed whether these inhibitory effects is due to induction of apoptosis, and we have tried to evaluate the anti-tumor effect of selenium, through VASP, AMPK, mTOR and Akt signal pathway.
(Cancer Prev Res 15, 313-319, 2010)
Key Words: Selenium, Apoptosis, B16F10 melanoma cells, AMPK, VASP
서 론
지구의 온난화 현상으로 인해 기온이 점점 상승하고, 햇볕에 대한 노출이 점점 많아짐에 따라 피부암 발생이 증가하고 있으며 미국의 경우에는 20년 전과 비교했을 때 발생률이 2배나 증가하였다. 피부암 중에서 악성 흑
색종(malignant melanoma)은 전체 피부암의 4% 정도이지 만 피부암 사망자의 79%를 차지할 정도로 독성이 매우 강하다.1) 이처럼 악성 흑색종은 전 세계적으로 발생율이 급속히 늘어나고 있는 실정이며, 전이가 매우 급속하게 진행되고, 보통 암세포가 전이되지 않는 뇌, 심장까지 영 향을 미칠 만큼 높은 전이율을 나타낸다.
Ena/VASP family인 vasodilatator-stimulierte phosphoprotein
(VASP)는 초파리 단백질인 enabeled (Ena)와 구조적, 기능 적으로 많은 유사성을 보이기 때문에, 이러한 단백질군 을 Ena/VASP family라고 하고, Ena를 포유동물에서는 Mena라고 한다.2) Ena/VASP family는 오랜 진화과정을 통 하여 높게 보존되어 왔으며, 서로 다른 2개의 영역 EVH1 및 EVH2로 구성되어 있다.3) VASP는 actin, profilin, vincu- lin, talin 등과 연결되어 세포의 증식과 이동, 세포-세포 접촉을 조절하며 세포의 형태를 변화시키는 다양한 신 호경로와 연결되어 있다. VASP는 focal adhesion proteins과 vinculin, zyxin을 연결시키고, profilin과 focal adhesion pro- teins을 중재하는 기능을 한다.4) 특히 VASP는 혈액의 혈 소판에서 cAMP- 및 cGMP-의존성 단백질 키나아제의 기 질로서 작용하며, Ser-239 부위에서 VASP의 인산화는 내 피세포에서 nitric oxide/cyclic GMP 신호의 생화학적 마커 (biochemical marker)라고 보고되었다.5) VASP는 아미노산 의 위치에 따라 Ser-157, Ser-239, Thr-278 부위에서 단백 질 키나아제에 의해 조절되며, Ser-Thr 키나아제 역할을 하는 AMP-activated protein kinase (AMPK)가 여기에 관여 한다. 특히 AMPK에 의한 VASP Thr-278 위치에서의 인산 화는 액틴-세포골격의 재배치가 일어나는 과정에 분자 적 신호를 보낸다. 즉 VASP는 AMPK의 새로운 기질인 것으로 밝혀졌다.6)
AMPK는 hypoxia, 영양분 결핍, 운동, heat shock 등과 같은 스트레스에 의해 세포 내에서 ATP가 결핍되면 활 성화된다. AMPK는 세포에서 에너지 센서로서의 역할을 하며, 암세포에서 AMPK가 활성화되면 apoptosis가 유도 되는 것으로 알려져 있다.7) AMPK와 mammalian target of rapamycin (mTOR)은 세스트린(sestrins) 유전자의 지시를 받아, 세스트린이 부족하게 되면 AMPK가 저활성화되 고, 표적단백질인 TOR가 과다하게 활성화되어 노화가 촉진된다.8) Akt는 Ser-Thr 키나아제의 일종으로 세포의 증식, 분화, 성장에 관여하며, IGF-1에 의해서 활성화된 PI3K/Akt 신호경로는 Bad나 Caspase-9와 같은 pro-apop- totic protein의 발현을 저해하여 apoptosis를 억제하는 것 으로 알려져 있다.9,10)
과일, 채소, 녹차 등에 함유된 천연 식물성 화학물질인 파이토케미컬(phytochemical)의 일종인 셀레늄(selenium)이 Akt 신호경로의 활성을 저해함으로써 in vitro 실험에서 암세포의 증식을 억제하고 apoptosis를 유도하며, in vivo 실험에서도 종양의 성장을 억제하는 것으로 밝혀졌다.11) 이는 Akt 신호경로의 조절이 암세포의 비정상적인 증식 을 억제할 수 있는 중요한 신호분자로 작용할 수 있다는 것을 의미하는데, Akt는 암세포의 apoptosis를 억제하고 혈관신생과 전이에도 관여하는 것으로 알려져 있다.12)
mTOR는 PI3K/Akt 신호경로의 하위경로로서 PI3K에 의 해 활성화된 Akt가 mTOR를 활성화시킴으로서 세포의 증식과 분화를 촉진시키는 것으로 알려져 있다.13) 따라서 본 연구에서는 B16F10 피부암 세포에 다양한 농도 별로 셀레늄을 처리했을 때, B16F10 세포의 증식이 억제되는지를 밝혀내고, 이러한 암세포의 증식 억제 효 과가 apoptosis의 유도에 의한 것인지를 VASP, AMPK, mTOR, Akt 신호경로를 통하여 알아보고자 하였다.
재료 및 방법 1. 재료
본 실험에서 사용된 셀레늄은 Sigma (Sigma Aldrich, St.
Louis, MO)에서 구입하여 증류수에 녹인 뒤 10 mM stock 으로 만들어 -20oC에 보관하여 사용하였다. 3-(4,5-dime thylthiazol-2-yl)-2.5-diphenyltetrzolium bromide (MTT)는 Sig- ma (Sigma Aldrich, St. Louis, MO)에서 구입하여 5 mg/ml stock으로 만들어서 사용하였다. 또한, Compound C와 rapa- mycin은 Calbiochem (San Diego, CA)에서 구입하여 DMSO에 녹인 뒤 각각 10 μM과 50 nM stock으로 만들어 사용하 였다.
2. 세포 배양
B16F10 세포는 한국세포주은행에서 분양 받았으며, 10% 우태아혈청(fetal bovine serum, FBS, Gibco BRL, Grand Island, NY, USA)과 1% antibiotics (100 mg/l streptomycin, 100 U/ml penicillin)가 포함된 RPMI 1640 배지(WelGENE Inc., Seoul, Korea)를 사용하여 5% CO2, 37oC 조건하에서 배양하였다. 매 48시간마다 Trypsin-EDTA (WelGENE Inc., Seoul, Korea)를 이용하여 세포를 부유상태로 만든 다 음 세포를 1×106개/ml로 분주하고 계대하여 실험에 사용 하였다.
3. MTT assay에 의한 암세포의 생존율 측정
세포배양용 12 well plate에 B16F10 세포를 1×104개/ml 로 분주하고 24시간 동안 배양시킨 후 셀레늄을 처리하 였다. Rapamycin 처리시에는 rapamycin을 30분 먼저 처리 한 후 셀레늄을 처리하였으며 3-(4,5-dime thylthiazol-2-yl)- 2,5-diphenyltetrazolium bromide (MTT) 용액(5 mg/ml)을 30 μl씩 첨가하여 1시간 동안 CO2 incubator에서 배양하였 다. MTT시약이 들어있는 배지를 제거한 후에 DMSO를 150 μl 넣어 well에 생성된 formazan을 모두 녹인 후, 96 well plate에 100 μl씩 옮겨서 Microplate Reader (BIO-RAD Laboratories, Inc. USA)로 595 nm에서 흡광도를 측정하였다.
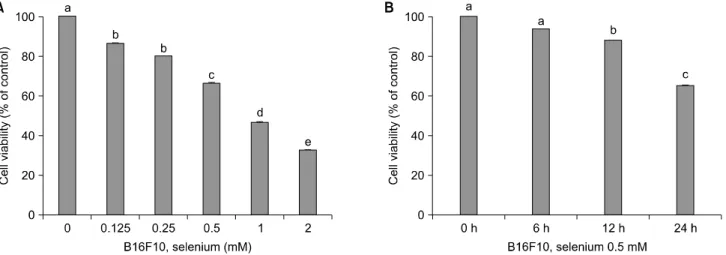
Fig. 1. Selenium inhibits cell proliferation in B16F10 melanoma cells. Cell viability was measured by MTT assay. Treatment of selenium inhibits cell proliferation. Cells were treated with 0.125∼2 mM of selenium (A). Cells were treated with selenium 0.5 mM in 6∼24 h (B). a∼ep<0.05 (each experiment’s n=3).
측정은 모두 세 번 하였으며, 그에 따르는 평균값과 표준 오차는 Microsoft Excel program을 이용하여 분석하였다.
4. Fluorescence-Activated Cell Sorting (FACS)에 의한 apoptosis 관찰
Apoptosis는 FITC-Annexin V apoptosis detection kit (BD PharmingenTM, San Diego, CA, USA)를 사용하여 측정하였 다. AnnexinV-PI staining을 하기 위해, B16F10 세포에 셀 레늄을 농도별(0 mM, 0.5 mM, 1 mM, 2 mM)로 처리한 후 24시간 배양하였다. 처리된 세포는 차가운 PBS로 세 척하고 나서 trypsin-EDTA로 모은 다음, 1×106 cells/ml의 농도에서 binding buffer로 suspension하였다. 그 다음에 1×105개의 B16F10 세포를 Annexin V-FITC와 prodipium io- dide (PI)로 15분간 염색한 후, 1시간 정도 flow cytometry (Becton-Dickinson Biosciences, Drive Frankline Lages, NJ, USA)로 분석하였다.
5. Western blotting
셀레늄을 농도별로 처리한 후 6시간 동안 CO2 in- cubator에서 배양한 후, 단백질을 추출하기 위하여 RIPA lysis buffer [50 mM Tris-HCl (pH 8.0), 150 mM NaCl, 1%
NP 40, 0.5% sodium deoxycholate, 1 mM phenylmethysul- fonyl]를 각 well에 150 μl씩 첨가하여 반응시킨 후, 14,000 rpm, 4oC에서 20분 동안 원심분리 한 후 상등액을 취하였다. 그런 다음, ELISA-reader 595 nm에서 흡광도를 측정하여 각 표본의 단백질 농도를 결정하였다. 5×
Laemmi sample buffer (loading dye; 250 mM Tris-Cl (pH 6.8), 40% glycerol, 4% β-mercaptoethanol, 0.08% bromophenol
blue, 8% SDS)와 B16F10 세포에서 추출한 단백질을 혼합 하여 표본을 제작한 후, p-AMPK, VASP, p-mTOR, p-Akt, β-actin의 1차 항체를 Cell Signaling Technology (Beverly, MA, USA)로부터 구입하여 각각 희석한 후, 반응시켰다.
2차 항체는 GenDEPOT (Barker, TX)에서 구입하여 각각 희석하여 1시간 반응시킨 다음, LAS-4000 (Fuji Film, Japan)을 이용하여 감광하였다.
6. 통계처리
통계 프로그램인 SPSS 17.0을 사용하여 실험설계에 대 한 분산분석은 ANOVA로, 각 처리군들 과의 비교는 일 원배치 분산분석을 실시하여 검정하였다. 각 자료는 3번 이상의 반복된 실험을 통하여 얻어진 결과로 검정하였 고 p<0.05인 경우 통계적으로 유의하다고 판정하였다.
결과 및 고찰
1. B16F10 세포의 증식에 미치는 셀레늄의 효과
셀레늄이 B16F10 세포의 증식을 억제하는지를 관찰하 기 위해 MTT assay 실험을 수행하였다. Fig. 1A에서 보는 바와 같이, 셀레늄을 농도별(0.125 mM, 0.25 mM, 0.5 mM, 1 mM, 2 mM)로 처리했을 때, 농도가 증가할수록 B16F10 세포의 증식이 억제되는 것을 확인하였다. 즉 B16F10 세 포에 있어서 셀레늄 0.125 mM 처리시에는 약 86%, 0.25 mM 처리시에는 약 80%, 0.5 mM 처리시에는 약 67%, 1 mM 처리시에는 약 47%, 2 mM 처리시에는 약 32% 정도 의 생존율이 나타난 것으로 확인되었다. 이처럼 셀레늄 을 농도별로 처리했을 때, B16F10 세포의 생존율이 뚜렷
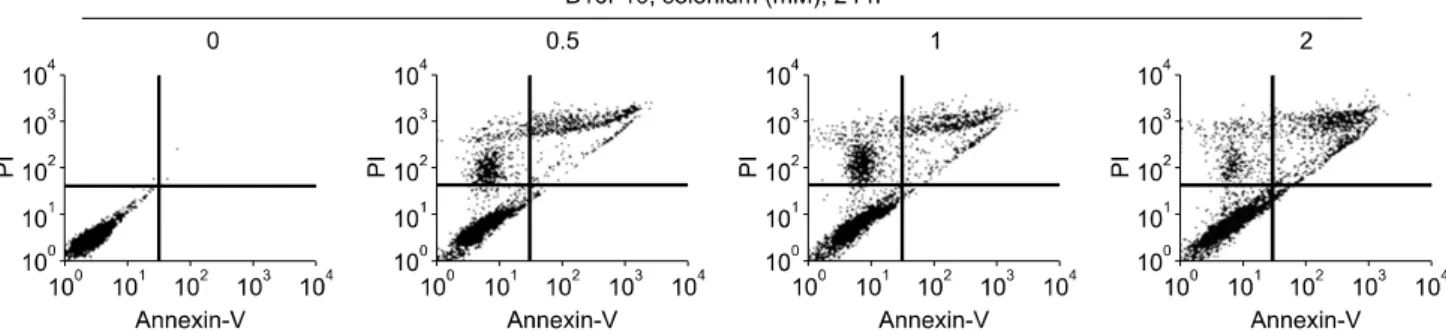
Fig. 2. Treatment of selenium induces apoptosis in B16F10 melanoma cells. Cell were treated with 0.5∼2 mM of selenium for 24 h. Apoptosis was measured by Annexin V-fluorescein isothiocyanate and popidium iodide (PI).
Fig. 3. Selenium effects on p-mTOR, VASP, p-AMPK, and p-AKT in B16F10 melanoma cells. Cells were treated with 0.125∼2 mM of selenium. Protein levels of them were determined by Western blotting.
하게 감소되는 것을 알 수 있었으며, 셀레늄 1 mM 이상 처리시에는 피부암 세포의 생존율이 50% 미만인 것으로 관찰되었다. 또한 셀레늄의 농도를 0.5 mM로 일정하게 처리하고, 시간의 경과(6 h, 12 h, 24 h)에 따른 B16F10 세포의 생존율을 측정한 결과, 6시간이 지난 후에는 약 93%, 12시간이 지난 후에는 약 87%의 미미한 차이를 보 였지만, 24시간이 지난 후에는 약 65%의 생존율을 나타 냈다.
2. 셀레늄을 통한 B16F10 세포의 apoptosis 유도 효과
이와 같은 암세포의 증식 억제 효과가 apoptosis 유도에 의해 일어난 것인지를 알아보기 위해 Annexin V-propo- dium iodide staning을 이용하여 apoptosis가 일어난 세포를 측정하였다. Fig. 2에서와 같이, 셀레늄을 0.5 mM, 1 mM, 2 mM로 24시간 동안 처리했을 때, MTT assay에서의 실 험결과와 마찬가지로 셀레늄의 농도가 더 높아질수록 더 많은 apoptosis가 유도되는 효과를 확인할 수 있었다.
3. 셀레늄이 VASP, p-AMPK, p-mTOR, p-Akt의 신 호경로에 미치는 효과
셀레늄을 MTT assay에서와 동일한 농도(0.125 mM, 0.25 mM, 0.5 mM, 1 mM, 2 mM)로 처리했을 때, VASP, p-AMPK, p-mTOR, p-Akt의 신호경로를 알아보기 위해 Western blotting을 실시하였다. Fig. 3에서 보이는 것과 같 이 셀레늄의 농도가 높아질수록 p-AMPK의 활성이 증가 하였다. AMPK의 활성이 증가하게 되면 COX-2의 발현 이 감소하게 되고, COX-2의 조절을 받는 VEGF가 결국 감소됨에 따라 angiogenesis가 억제됨으로써 암의 전이가 억제되는 것으로 알려져 있다.7,14) 반면에 셀레늄의 농도 가 높아질수록 VASP의 활성은 감소하는 것으로 관찰되었 다. Han 등은 유방암 세포의 이주와 침윤에 있어서 VASP 와 RHO family의 구성원인 Rac1의 발현이 암의 전이에 어
떠한 영향을 미치는지를 VASP-siRNA, Rac1-siRNA를 이용 하여 연구한 결과, VASP와 Rac1의 발현 수준이 높을수록 종양의 성장이 빠르게 진행된다고 발표하였다. 즉 VASP 의 발현이 약해질수록 종양의 성장은 느리게 진행되는 것이다.15) 또한 셀레늄의 농도가 높아질수록 p-mTOR와 p-Akt의 발현은 점차로 감소되었다. Akt-mTOR pathway 는 여러 종류의 암에서 세포의 성장과 분화에 영향을 미 치고 암세포에서 apoptosis가 유도되는 것을 방해한다. 세 포막에 존재하는 PI3K가 활성화되면 Akt가 활성을 나타 내어 BAD, GSK3β 등 여러 단백질을 인산화시킴으로써 암세포의 증식을 조절하는 것으로 알려져 있다.16) PI3K/
Akt 신호 경로는 mTOR의 상위조절자이며 Akt의 활성화 로 mTOR의 활성이 더욱 효과적으로 작용하는 것으로 보인다.17) 따라서 Fig. 3에서와 같이 셀레늄의 농도가 높 아질수록 p-mTOR와 p-Akt의 발현이 감소하는 것은 암세 포의 증식이 억제되는 것이라고 할 수 있다.
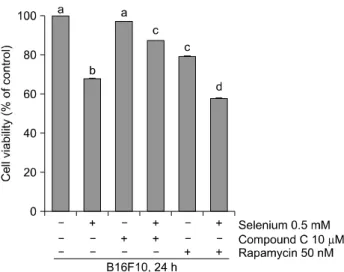
Fig. 4. Treatment of selenium, Compound C, and rapamycin.
0.5 mM selenium was treated with 10 μM of Compound C or 50 nM of rapamycin. B16F10 melanoma cells were pre-treated with 10 μM of Compound C or 50 nM of rapamycin. Cell viability was measured by MTT assay. a∼dp<
0.05 (each experiment’s n=3).
Fig. 5. Treatment of selenium, Compound C, and rapamycin.
0.5 mM selenium was treated with 10 μM of Compound C or 50 nM of rapamycin. B16F10 melanoma cells were pre- treated with 10 μM of Compound C or 50 nM of rapamycin.
Protein levels of p-mTOR, VASP and p-AMPK were detected by Western blotting.
4. AMPK 및 mTOR 저해제가 B16F10 세포의 증식에 미치는 영향
AMPK의 저해제인 Compound C와 mTOR의 저해제인 rapamycin을 B16F10 세포에 각각 처리했을 때, B16F10 세 포의 증식에 미치는 영향을 알아보기 위해 MTT assay를 실시하였다. 우선 Fig. 4에 나타난 바와 같이, 셀레늄 0.5 mM을 단독으로 처리하였을 때 B16F10 세포의 생존율이 약 68%로 나타났다. 그러나 Compound C 10 μM를 처리하 고 셀레늄 0.5 mM을 처리했을 때에는 B16F10 세포의 생 존율이 약 90% 정도가 되었다. 이는 AMPK의 저해제인 Compound C에 의해 셀레늄이 조절하는 B16F10 세포의 세 포증식억제 효과가 저해된 것으로 보인다. 한편 셀레늄 을 처리하지 않고 Compound C만 처리했을 때에는 B16F10 세포의 생존율에 아무 변화도 일어나지 않았다.
이러한 실험 결과는 셀레늄이 B16F10 세포의 생존율에 많은 영향을 미치며, 이러한 영향은 AMPK의 활성을 조 절함으로서 일어나는 것으로 여겨진다.7) 또한 AMPK의 활성을 저해했을 때에는 셀레늄이 B16F10 세포의 생존 율에 영향을 미치지 못하는 것으로 보인다.
마찬가지 방법으로 셀레늄 0.5 mM을 처리하고 mTOR 의 저해제인 rapamycin 50 nM을 처리했을 때에는 B16F10 세포의 생존율은 약 57% 정도가 되었다. 이러한 결과는 셀레늄 0.5 mM을 처리했을 때 B16F10 세포의 생존율 68%와 비교했을 때, 암세포의 증식에 관여하는 mTOR의
활성이 저해되어 B16F10 세포의 증식을 억제했기 때문 으로 보인다. 또한 셀레늄을 처리하지 않고 rapamycin만 단독으로 처리했을 때에는 B16F10 세포의 생존율이 약 80%에 이르렀으며, 이는 mTOR의 활성이 저해되어 세포 의 증식이 억제되었기 때문인 것으로 여겨진다.17)
5. AMPK 및 Akt 저해제에 의한 VASP, p-AMPK, p-mTOR의 신호경로의 조절
Compound C와 rapamycin을 B16F10 세포에 각각 처리 했을 때, 이 같은 저해제에 따른 VASP, p-AMPK, p-mTOR 단백질의 발현 양상을 알아보기 위해 Western blotting을 실시하였다. 그 결과, Fig. 5에서 나타난 바와 같이, Com- pound C 10 μM를 처리하고 셀레늄 0.5 mM을 처리했을 때에는 p-mTOR, VASP의 발현이 증가하였으나 p-AMPK 의 활성은 감소하였다. p-AMPK의 활성이 감소한 것은 AMPK의 활성이 AMPK의 저해제인 Compound C에 의해 영향을 받았으며, 이러한 실험 결과는 동일한 조건에서 실시한 Fig. 4의 MTT assay 결과와 일치한다.7) 한편 셀레 늄을 처리하지 않고 Compound C만 처리했을 때에도 p-mTOR, VASP의 발현은 증가하였지만 AMPK의 활성은 Compound C에 의해 영향을 받아 감소하였다.7) 이와 같 은 결과를 통해 AMPK가 VASP와 mTOR의 조절에 영향 을 미치는 중요분자로서 작용할 수 있음을 확인하였다.
Fig. 4의 MTT assay 결과와 비교해 보았을 때 AMPK를 저해하면 세포 생존율이 증가하며, 이에 따라 p-mTOR, VASP의 발현 역시 증가하는 것으로 보인다.7,15,17) 마찬가지로 mTOR의 저해제인 rapamycin 50 nM을 처 리하고 셀레늄 0.5 mM을 처리했을 때, 대조군에 비하여 AMPK의 활성이 감소하였고, p-mTOR의 발현 또한 감소
하였다. 이는 Fig. 4의 MTT assay 결과와 일치하며, mTOR 가 활성화되지 못하여 apoptosis가 유도된 것으로 추측된 다.7) 또한 셀레늄을 처리하지 않고 rapamycin만 처리했을 때에도 역시 p-AMPK와 p-mTOR의 발현이 감소되었다. 선 행연구들에서 암세포에 셀레늄을 처리하였을 때 AMPK 가 활성화 되며, 활성화된 AMPK는 mTOR의 활성을 저 해하는 것으로 보고되었다.11) 그러므로 이와 같은 실험 결과를 종합해 본다면, B16F10 세포에 셀레늄을 처리하 였을 때 AMPK가 활성화되며, 활성화 된 AMPK는 VASP 와 p-mTOR의 발현을 조절하는 것으로 보인다. 이는 셀 레늄이 유도하는 apoptosis에 있어서 AMPK가 중요한 신 호전달 분자로서 작용할 수 있음을 의미한다.
결 론
Ena/VASP family인 vasodilatator-stimulierte phosphoprotein (VASP)는 액틴 등과 연결되어 세포의 증식과 이동, 세포- 세포 접촉을 조절하는 다양한 신호경로에 관여한다.
VASP는 혈액에서 cAMP- 및 cGMP-의존성 단백질 키나 아제의 기질로서 작용하며, 아미노산의 위치에 따라 Ser- 157, Ser-239, Thr-278 부위에서 단백질 키나아제에 의해 조절되는데, Ser-Thr 키나아제 역할을 하는 AMP-activated protein kinase (AMPK)가 여기에 관여한다. 특히 AMPK에 의한 VASP Thr-278 위치에서의 인산화는 액틴-세포골격 의 재배치가 일어나는 과정에 분자적 신호를 보내며, 최 근에 VASP는 AMPK의 새로운 기질인 것으로 밝혀졌다.
AMPK는 세포에서 에너지 센서로서의 역할을 하며, 암 세포에서 AMPK가 활성화 되면 apoptosis가 유도되는 반 면, mammalian target of rapamycin (mTOR)이 활성화 되면 암의 증식을 촉진하는 것으로 알려져 있다. 또한 Akt는 Ser-Thr 키나아제의 일종으로 세포의 증식, 분화, 성장에 관여하며, 암세포의 apoptosis를 억제하고 혈관신생과 전 이에도 관여하는 것으로 알려져 있다. 본 연구에서는 B16F10 피부암 세포에 식물에 포함된 파이토케미컬의 일종인 셀레늄을 처리했을 때, MTT assay, Fluorescence- Activated Cell Sorting (FACS), Western blotting을 통하여 B16F10 세포의 증식이 억제되는지를 관찰하고, 이러한 증식 억제 효과가 apoptosis의 유도에 의한 것인지를 밝혔 으며, 암 증식에 관련된 VASP, AMPK, mTOR, Akt 신호경 로를 통하여 셀레늄의 항암 효과를 알아보고자 하였다.
감사의 글
이 논문은 2010년도 한남대학교 학술연구조성비 지원
에 의하여 연구되었음.
참 고 문 헌
1) Bedogni B, Powell MB. Hypoxia, melanocytes and melanoma- survival and tumor development in the permissive microen- vironment of the skin. Pigment Cell Melanoma Res 22, 166-174, 2009.
2) Krause M, Leslie JD, Stewart M, Lafuente EM, Valderrama F, Jagannathan R, Strasser GA, Rubinson DA, Liu H, Way M, Yaffe MB, Boussiotis VA, Gertler FB. Lamellipodin, an Ena/VASP ligand, is implicated in the regulation of lamel- lipodial dynamics. Dev Cell 7, 462-463, 2004.
3) Reinhard M, Jarchau T, Walter U. Actin-based motility: stop and go with Ena/VASP proteins. Trends Biochem Sci 26, 243- 249, 2001.
4) Price CJ, Brindle NP. Vasodilator-stimulated phosphoprotein is involved in stress-fiber and membrane ruffle formation in endothelial cells. Arterioscler Thromb Vasc Biol 20, 2051-2056, 2000.
5) Ibarra-Alvarado C, Galle J, Melichar VO, Mameghani A, Schmidt HH. Phosphorylation of blood vessel vasodilator-sti- mulated phosphoprotein at serine 239 as a functional bio- chemical marker of endothelial nitric oxide/cyclic GMP signa- ling. Mol Pharmacol 61, 312-319, 2002.
6) Blume C, Benz. PM, Walter U, Ha J, Kemp BE, Renné T.
AMP-activated Protein Kinase Impairs Endothelial Actin Cytoskeleton Assembly by Phosphorylating Vasodilator-stimu- lated Phosphoprotein. J Bio Chem 282, 4610-4012, 2007.
7) Hwang JT, Kim YM, Baik HW, Lee SK, Ha JH, Park OJ.
Selenium regulates COX-2 and ERK signaling pathways by activating AMPK in colon cancer cells. Cancer Research 66, 10057-10063, 2006.
8) Lee JH, Budanov AV, Park EJ, Birse R, Kim TE, Perkins GA, Ocorr K, Ellisman MH, Bodmer R, Bier E, Karin M. Sestrin as a feedback inhibitor of TOR that prevents age-related pathologies. Science 327, 1223-1228, 2010.
9) Carnero A. The PKB/AKT pathway in cancer. Curr Pharm Des 16, 34-44, 2010.
10) Milacic V, Banerjee S, Landis-Piwowar KR, Sarkar FH, Majumdar AP, Dou QP. Curcumin inhibits the proteasome activity in human colon cancer cells in vitro and in vivo. Cancer Research 68, 7283-7292, 2008.
11) Lee YK, Park SY, Kim YM, Kim DC, Lee WS, Surh YJ, Park OJ. Suppression of mTOR via Akt-dependent and -indepen- dent mechanisms in selenium-treated colon cancer cells: in- volvement of AMPK alpha1. Carcinogenesis 31, 1092-1099, 2010.
12) Kim D, Chung J. Akt: Versatile Mediator of Cell Survival and Beyond. J Biochem Mol Biol 35, 105-115, 2002.
13) Guo Y, Liang X, Lu M, Weng T, Liu Y, Ye X. Mammalian target of rapamycin as a novel target in the treatment of hepatocellular carcinoma. Hepatogastroenterology 57, 913-918,
2010.
14) Veikkola T Alitalo K. VEGF's Receptors and angiogenesis.
Semin Cancer Biol 9, 211-220, 1999.
15) Han G, Fan B, Zhang Y, Zhou X, Wang Y, Dong H, Wei Y, Sun S, Hu M, Zhang J, Wei L. Positive regulation of migration and invasion by vasodilator-stimulated phospho- protein via Rac1 pathway in human breast cancer cells. Oncol
Rep 20, 929-939, 2008.
16) Brazil DP, Yang ZZ, Hemmings BA. Advances in protein kinase B signalling: Aktion on multiple fronts. Trends Biochem Sci 29, 233-242, 2004.
17) Guertin DA, Sabatini DM. An expanding role for mTOR in cancer. TRENDS in Molecular Medicine 11, 353-361, 2005.