340 책임저자:김영민, 305-811, 대전시 유성구 전민동 461-6
한남대학교 생명나노과학대학 생명과학과 Tel: 042-629-8753, Fax: 042-629-8751 E-mail: [email protected]
접수일:2011년 10월 13일, 1차 수정일:2011년 10월 18일, 2차 수정일:2011년 10월 24일, 게재승인일:2011년 10월 27일
Correspondence to:Young Min Kim
Department of Biological Sciences, College of Life Science and Nano Technology, Hannam University, 461-6, Jeonmin-dong, Yuseong-gu, Dae- jeon 305-811, Korea
Tel: +82-42-629-8753, Fax: +82-42-629-8751 E-mail: [email protected]
HT - 29 대장암 세포에서 mTOR - VASP 신호경로 조절을 통한 쿼세틴의 Apoptosis 효과
한남대학교 생명나노과학대학 1생명과학과, 2식품영양학과
이세희1ㆍ김인섭1ㆍ박송이1ㆍ박옥진2ㆍ김영민1
Quercetin Induces Apoptosis via Regulation of mTOR-VASP Signaling Pathway in HT-29 Colon Cancer Cells
Se Hee Lee1, In-Seop Kim1, Song Yi Park1, Ock Jin Park2 and Young Min Kim1 Departments of 1Biological Sciences, 2Food and Nutrition,
College of Life Science and Nano Technology, Hannam University, Daejeon 305-811, Korea
Quercetin, one kind of phytochemical, is known to an important anti-cancer flavonoid. In this study, we suggest that quercetin-treated HT-29 colon cancer cells inhibited cell proliferation and induced apoptosis through AMPK, mTOR, VASP and COX-2 signaling pathway. AMP-activated protein kinase (AMPK) acts as an intracellular energy sensor and activated by depletion status of ATP. AMPK has been recognized as an important up-regulation signaling mediator involved in the down-regulation of the mTOR and VASP pathway. Mammalian target of rapamycin (mTOR) plays a central role in regulating of cell proliferation. mTOR signaling pathway is immoderately activated in many type of cancer. We determined with MTT assay, Western blotting and FACS. Treatment of Compound C (AMPK inhibitor) resulted in increased cell proliferation of cancer cells and showed that mTOR, COX-2 and VASP are regulated by AMPK. But, COX-2, VASP are directly controlled by quercetin in AMPK-independent pathway. Also, treatment of rapamycin (mTOR inhibitor), celecoxib (COX-2 inhibitor) and co-treatment with quercetin resulted in greater decrease of cell proliferation in colon cancer cells. In addition, mTOR can modulate the expression of COX-2, and rapamycin functions as an activator of AMPK. The results indicate that quercetin decreases cell proliferation and induces apoptosis by inhibiting mTOR and COX-2. The inhibition of mTOR (or rapamycin) can control the expression of VASP, AMPK. (Cancer Prev Res 16, 340-347, 2011)
Key Words: AMPK, mTOR, VASP, Quercetin, Apoptosis, HT-29 colon cancer cells
서 론
전 세계적으로 암에 대한 연구는 끊임없이 진행되고 있으나, 암 발생과 그로 인한 사망률은 계속하여 증가하
고 있으므로 다양한 생물화학적 방법을 통해 암을 예방 하고 치료하려는 연구가 수행되고 있다. 대장암은 주로 노령인구에서 발생이 증가하는 것으로 알려져 있으며, 우리나라의 인구가 점차 고령화되고 음식문화가 서구화 되어감에 따라 사회적으로 매우 주목 받고 있는 질환으
로서 이를 예방하고 치료하기 위한 방법을 찾는 연구에 대한 관심이 높아지고 있다.1)
식물에서 추출한 파이토케미컬(phytochemical)이 세포 내 신호전달 분자들을 조절함으로써 암세포의 증식과 전이를 억제한다는 연구가 진행되어 왔으며, 이들이 조 절하는 신호경로에 대한 관심이 점차 높아지고 있다. 파 이토케미컬의 한 종류인 쿼세틴(quercetin)은 붉은 양파에 서 추출할 수 있는 물질로 항염 및 항암효과를 나타낸 다.2) 최근 연구에 의하면 쿼세틴이 MAPKs, Akt/protein kinase B 등의 발현을 조절하는 것으로 밝혀졌으며, COX-2와 같은 표적 분자의 조절뿐만 아니라 암세포의 생존, 증식, 사멸을 조절할 수 있는 것으로 밝혀졌다.3) AMP-activated protein kinase (AMPK)는 세포의 에너지 대사 상태를 감지하는 역할을 하며, 세포 내 ATP가 고갈 되거나 AMP의 level이 높은 상태일 때 활성화되며, 쿼세 틴이 지방세포에서 AMPK를 활성화시켜 apoptosis를 유 도함이 보고되었다.4,5) AMPK의 활성은 caspase를 포함한 p53과 p21과 같은 상위조절자에 의해 일어나며, 활성화 된 AMPK가 COX-2, mTOR, Akt와 같은 암세포의 증식과 성장과 관련된 분자를 억제시킴으로써 다양한 암세포에 서 apoptosis를 유도하는 것으로 알려져 있다.6)
mTOR (mammalian target of rapamycin)는 AMPK에 의해 서 조절되며, 상위조절 단백질인 Akt에 의하여 활성화되 는 것으로 밝혀졌다. Akt-mTOR 경로는 많은 암에서 과 활성화되어 있으며, mTOR는 세포의 성장이나 증식에 영향을 미치는 중요한 조절자로서 종양형성에 관여하는 단백질로 알려져 있다.7) 특히 HT-29 대장암 세포에서 mTOR의 발현을 저해하였을 때, 세포 성장인자의 자극 에 의해 암세포의 성장이 저해되는 것이 보고되었다.8) COX (cyclooxygenase)는 프로스타노이드(prostanoid)라고 불리는 중요한 생물학적 매개체의 형성을 위한 효소로 써, 프로스타글란딘(prostaglandin)의 합성에 COX의 두 가 지 isoform인 COX-1과 COX-2가 관여한다.9,10) COX-1은 대부분의 조직에서 housekeeper enzyme으로 항상성 유지 기능에 관여하며, COX-2는 interleukin-1 (1L-1)과 같은 cy- tokine이나 다양한 성장인자와 종양 촉진인자 등의 자극 에 의해 과발현되며 특히 암 조직이나 염증조직에서 높 게 나타난다.11) 또한 COX-2는 대장암세포의 생존에 관 여하는 주요 단백질로 알려져 있으므로 COX-2의 저해 는 대장암을 조절하는 효과적인 방법 중의 하나로 밝혀 졌다.12)
VASP (vasodilator-stimulated phosphoprotein)는 Ena/VASP Family라 하며, 이는 초파리의 단백질인 enabeled (Ena)와 구조ㆍ기능적으로 유사성을 보인다.13) VASP는 세포의
증식과 이동, 세포와 세포간의 접촉을 조절하며 세포의 형태변화와 관련된 다양한 신호경로와 연결되어 있다.14) VASP의 인산화는 VASP의 아미노산 위치에 따른 세 부 위에서 단백질 키나아제에 의해 조절되며 Ser-Thr 키나 아제 역할을 하는 AMPK가 여기에 관여한다. AMPK에 의한 VASP Thr-278 위치에서의 인산화는 액틴과 세포골 격 사이의 재배치 과정에 분자적, 세포적인 신호를 보낸 다.15)
본 연구에서는 HT-29 대장암 세포에 쿼세틴을 처리했 을 때, 암세포의 증식이 억제되는지를 알아보고자 하였 다. 또한 쿼세틴을 처리했을 때, AMPK, COX-2, mTOR, VASP 단백질들이 어떠한 영향을 받으며, 또한 어떠한 연 관이 있는지를 알아보기 위하여 COX-2와 mTOR의 저해 제를 단독으로 처리하거나 이들을 각각 쿼세틴과 병행 처리하여 이들 단백질들의 활성과 저해효과를 알아보고 자 하였다.
재료 및 방법 1. 재료
본 실험에서 사용된 쿼세틴은 Sigma (Sigma Aldrich, St.
Louis, MO)에서 구입하여 Dimethyl sulfoxide (DMSO)에 녹 인 뒤 50 mM stock으로 만들어 -20oC에 보관하여 사용 하였다. 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrzolium bro- mide (MTT)는 Sigma에서 구입하여 5 mg/ml stock으로 만 들어서 사용하였다. Celecoxib는 Calbiochem (San Diego, CA)에서 구입하여 100 mM stock으로 만들어 사용하였고, rapamycin은 Calbiochem에서 구입하여 100 ng/ml stock으 로 만들어 -20oC에 보관하여 사용하였다.
2. 세포배양
본 실험에 사용된 HT-29세포는 America Type Culture Collection (ATCC, Gaithersburg, MD)에서 분양 받았으며, 10% 우태아혈청(fetal bovine serum, FBS)과 1% antibiotics (100 mg/l streptomycin, 100 U/ml penicillin)가 포함된 RPMI 1,640 배지(WelGENE lnc., Seul, Korea)를 사용하여 5%
CO2, 37oC 조건 하에서 배양하였다. 매 48시간마다 Tryp- sin-EDTA (WelGENE lnc., Seul, Korea)을 이용하여 세포를 부유상태로 만든 다음 세포를 1×105개로 분주하여 계대 하였다.
3. MTT assay에 의한 암세포의 생존율 측정
세포배양용 12 well plate에 HT-29 세포를 1×104개/ml로 분주하고 24시간 동안 배양시킨 후 쿼세틴을 처리하였
Fig. 1. Anti-proliferatory effects of quercetin in HT-29 colon cancer cells. Cell viability was measured by MTT assay. Treat- ment of quercetin inhibits cell proliferation. Cells were treated with quercetin 25∼100μM for 24 h. a∼cp<0.05 (each experiment’s n=3).
다. COX-2의 저해제인 celecoxib와 mTOR의 저해제인 ra- pamycin과 병행처리 시에는 celecoxib와 rapamycin을 30분 먼저 처리한 후 쿼세틴을 처리하였으며 MTT용액(5 mg/
ml)을 30μl씩 첨가하여 CO2 incubator에서 1시간 동안 배 양하였다. MTT solution이 들어있는 배지를 제거한 후에 DMSO 150μl를 넣어 well에 생성된 formazan을 모두 녹인 후, 96 well plate로 100μl씩 옮겨서 Microplate Reader (BIO- RAD Laboratories, Inc. USA)로 595 nm에서 흡광도를 측정 하였다.
4. Fluorescence-Activated Cell Sorting (FACS)에 의한 apoptosis 관찰
Apoptosis는 FITC-Annexin V apoptosis detection kit (BD Pharmingen, San Diego, CA, USA)를 사용하여 측정하였다.
AnnexinV-PI staining을 하기 위해 HT-29 세포에 쿼세틴을 농도별(0μM, 25μM, 50μM, 100μM)로 처리한 후 24시 간 배양하였다. 처리된 세포는 PBS로 세척한 후 try- sin-EDTA로 모은 다음, 1×106 cells/ml의 농도에서 binding buffer로 suspension하였다. 그런 다음 1×105개의 세포를 Annexin V-FITC와 prodipium iodide (PI)로 15분간 염색한 후, 1시간 정도 flow cytometry (Becton-Dickinson Bidscien- ces, Drive Frankline Lages, NJ, USA)로 분석하였다.
5. Western blotting
세포배양용 6 well plate에 HT-29세포를 1×105개/ml로 분주하여 24시간 동안 배양한 다음 쿼세틴을 처리하였 다. Celecoxib와 mTOR의 저해제인 rapamycin을 병행처리 시에는 celecoxib와 rapamycin을 30분 먼저 처리한 후 쿼세 틴을 처리하였으며, 모든 물질을 처리한 후 6시간 동안 CO2 incubator에서 배양하였다. 단백질을 추출하기 위하 여 RIPA lysis buffer {50 mM Tris-HCL (pH 8.0, 150 mM NaCl, 1% NP40, 0.5% sodium deoxycholate, 1 mM PMSF)}
를 well 당 150μl씩 처리하여 반응시켜, 14,000 rpm, 4oC 에서 20분간 원심분리를 한 후 상등액을 취하여 정량하 였다. 그 후에 8% acrylamide gel에 sample을 loading하여 전기영동을 실시한 후 nitrocellulose membrane로 단백질을 transfer하는 과정을 거친 다음 2% bovine serum albumin (BSA)를 이용하여 blocking한 후, 1차, 2차 항체를 반응시 키고 감광하여 실험결과를 측정하였다.
6. 통계처리
통계 프로그램인 SPSS 17.0을 사용하여 실험설계에 대 한 분산분석은 ANOVA로, 각 처리군들과의 비교는 일원 배치 분산분석을 실시하여 검정하였다. 각 자료는 3번
이상의 반복된 실험을 통하여 얻어진 결과로 검정하였 고 p<0.05인 경우 통계적으로 유의하다고 판정하였다.
결 과
1. 쿼세틴이 HT-29 세포의 증식에 미치는 영향
쿼세틴이 HT-29 세포의 증식에 미치는 영향을 알아보 기 위해 쿼세틴을 농도별로 처리하여 MTT assay를 통하 여 암세포의 생존율을 측정하였다. 쿼세틴을 25μM, 50 μM, 100μM로 24시간 처리한 결과, 농도가 증가함에 따라 점차 HT-29세포가 25μM에서 70.1%, 50μM에서 43.4%, 100μM에서 35.9%의 생존율을 보여 각각 29.9%, 56.6%, 64.1%로 증식이 억제되는 것을 확인하였다(Fig.
1).
2. 쿼세틴에 의한 HT-29 세포 apoptosis 유도 효과
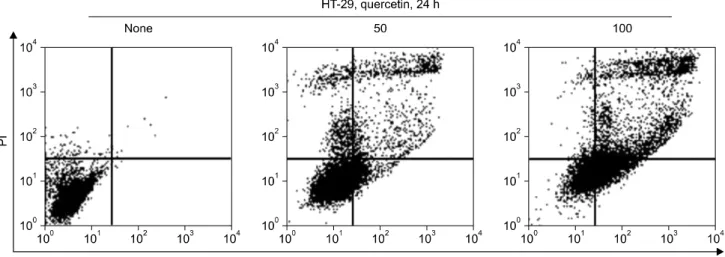
쿼세틴에 의한 HT-29 세포의 증식 억제 효과가 apopti- sis 유도에 의해 일어난 것인지를 확인하기 위해 Anne- xinV-PI staining을 통하여 apoptosis가 일어난 세포를 측정 하였다. 그 결과 Fig. 2에서와 같이 쿼세틴 50μM을 처리 하였을 때 22.12%, 100μM을 처리하였을 때 64.27%의 비율로 apoptosis가 유도되었음을 확인하였다.
3. 쿼세틴이 농도와 시간에 따른 AMPK, mTOR, COX-2, VASP의 발현
쿼세틴을 농도와 시간별로 처리했을 때 mTOR, AMPK,
Fig. 3. Quercetin effect on mTOR, AMPK, COX-2, VASP in HT-29 colon cancer cells. Cells were treated with quercetin 25∼
100μM for 6 h (A). Cells were treated with quercetin 50 μM for 3∼12 h (B). Protein levels of mTOR, AMPK, COX-2, VASP were determined by Western blotting.
Fig. 2. Apoptotic effects of quercetin were evaluated by fluorescence-activated cell-sorting analysis. Cells were treated with quercetin (control, 50μM, 100μM) for 24 h.
COX-2, VASP 단백질에 어떠한 영향을 미치는지 알아보 기 위하여 Western blotting을 실시하였다. 그 결과, Fig.
3A에서와 같이 25μM, 50μM, 100μM로 쿼세틴을 처리 하였을 때, 농도 의존적으로 mTOR, COX-2, VASP의 발 현이 감소하였으나, AMPK의 발현은 증가하는 것을 확 인할 수 있었다. 또한 시간의 변화(3 h, 6 h, 12 h)에 따라 mTOR, COX-2, VASP 발현이 감소하며 AMPK는 6시간째 가장 높은 발현을 나타내는 것을 확인하였다 (Fig. 3B).
4. AMPK 저해제가 HT-29세포의 증식 및 mTOR, COX-2, VASP 신호분자에 미치는 영향
AMPK의 저해제인 Compound C를 처리했을 때, 쿼세틴 이 HT-29세포의 증식에 미치는 영향을 알아보기 위해 MTT assay를 실시하였다. Fig. 4A에서와 같이 쿼세틴 50 μM과 Compound C 10μM을 각각 처리하였고, 또한 이 들을 동일한 농도로 병행 처리하였다. 쿼세틴 단독 처리 시 암세포가 약 70%의 생존율을 보였고, Compound C를 단독 처리했을 때 약 120%의 생존율을 보였으며, 이들을
병행 처리하였을 때에는 약 80%의 생존율을 보였다. 또 한 Compound C와 쿼세틴의 처리에 의한 AMPK-VASP 신 호경로 조절을 알아보기 위하여 Western blotting을 실시 하였다. Fig. 4B에서와 같이 AMPK가 저해되었을 때 VASP의 발현이 증가한 것으로 나타났으며, 이러한 실험 결과를 통해 AMPK가 mTOR를 조절하는 조절자로 작용 하는 것으로 여겨진다. 또한 HT-29 세포에서 쿼세틴에 의한 COX-2와 VASP의 직접적인 저해가 가능함을 확인 하였다.
5. COX-2와 mTOR의 저해제가 AMPK와 VASP의 활 성에 미치는 영향
COX-2 저해제인 celecoxib와 mTOR 저해제인 rapamycin 을 쿼세틴과 각각 또는 병행 처리하였을 때, HT-29 세포 증식에 미치는 영향을 알아보기 위해 MTT assay를 실시 하였다. Fig. 5A에서와 같이, 쿼세틴 50μM, celecoxib 40 μM, rapamycin 100 ng/ml을 각각 24시간 동안, 그리고 동 일한 농도로 병행 처리했을 때 HT-29세포의 증식을 확
Fig. 4. Co-treatment of Compound C and quercetin inhibits cell proliferation. Cells were pre-treatment with Compound C 10μM for 30 min and co-treated with quercetin 50μM for 24 h (A). Protein levels of the mTOR, AMPK, COX-2, VASP were determined by Western blotting (B). a∼dp<0.05 (each experiment’s n=3).
Fig. 5. Co-treatment of rapamycin and quercetin or celecoxib and quercetin inhibits cell proliferation. Cells were pre-treated with rapamycin 100 ng/ml or celecoxib 40μM for 30 min and co-treated with quercetin 50μM for 48 h. Cell viability were measured by MTT assay (A). Cells were pre-treated with rapamycin 100 ng/ml or celecoxib 40μM for 30 min and co-treated with quercetin 50μM for 6 h. Protein levels of mTOR, COX-2, AMPK, VASP were determined by western blotting (B). a∼dp<0.05 (each experiment’s n=3).
인하였다. 그 결과 celecoxib를 단독 처리하였을 때는 약 90%의 생존율을 보였고, 쿼세틴과 병행 처리하였을 때 는 약 75%의 생존율을 나타내었다. 그리고 rapamycin을 단독으로 처리하였을 때는 약 89%의 생존율을 나타내었 으며, 쿼세틴과 병행 처리하였을 때는 약 68%의 생존율 을 보였다.
마찬가지로 동일한 조건 하에서 mTOR, AMPK, COX- 2, VASP의 활성을 알아보기 위하여 Western blotting을 실
시하였다. Fig. 5B에서 나타낸 바와 같이 celecoxib를 단독 으로 처리하였을 때 COX-2의 발현이 감소되었고, 쿼세 틴과 병행 처리하였을 때에 발현이 더욱 감소된 것으로 보아 COX-2의 조절이 쿼세틴에 의해서도 가능하다는 것을 보여주었다. 또한 rapamycin을 단독으로 처리하였 을 때 mTOR의 발현이 감소되며, 쿼세틴과 병행 처리하 였을 때 발현이 더욱 감소한 것으로 보아 mTOR 역시 쿼세틴에 의해서 조절이 된다는 것을 알 수 있었다. 한편
rapamycin의 선 처리 후, 쿼세틴을 처리하였을 때 VASP의 발현이 더욱 감소되는 것으로 보아, mTOR의 활성이 VASP의 발현에 영향을 미칠 수 있는 것으로 확인되었다.
고 찰
그 동안 대표적인 서구형 암으로 알려져 온 대장암이, 국내에서도 식습관의 서구화 등으로 인하여 증가하고 있는 추세에 있다. 그러므로 과일과 채소에서 추출할 수 있는 식물성 화학물질인 파이토케미컬을 이용하여 세포 내 신호전달 단백질들을 조절함으로써 암세포의 증식 및 전이를 억제하고 apoptosis를 유도하려는 연구가 활발 히 진행되고 있다.
mTOR는 세포의 성장과 증식에 중요한 영향을 미치는 종양형성 단백질로 알려져 있으며, mTOR의 발현을 저 해하였을 때, 세포 성장인자의 자극에 의해 암세포의 성 장이 저해되는 것으로 보고되었다.7,8) COX-2는 주로 암 조직이나 염증조직에서 발현되며, COX-2의 저해가 대장 암을 조절하는 효과적인 방법 중의 하나로서 밝혀졌
다.11,12) VASP는 세포의 증식과 이동, 세포와 세포간의 접
촉을 조절하며 세포의 형태변화와 관련된 다양한 신호 경로와 연결되어 있다.14) AMPK는 세포에서 에너지 센서 역할을 하며, 특히 암세포에서는 AMPK의 활성이 apop- tosis를 유도하므로 그 역할에 대한 관심이 고조되고 있 다.6)
본 연구에서는 HT-29 세포에 쿼세틴을 처리했을 때 HT-29 세포의 증식이 억제가 되는지를 관찰하고 암세포 의 증식에 관련된 AMPK, VASP, COX-2, mTOR 신호경로 를 통하여 쿼세틴의 항암효과를 알아보고자 하였다. 이 를 위하여 쿼세틴을 농도별로 처리한 결과, Fig. 1에서 아무것도 처리하지 않은 대조군에 비하여 쿼세틴의 농 도가 높아짐에 따라 HT-29세포의 증식이 억제되는 것을 MTT assay를 통하여 확인하였다. 이러한 세포증식 억제 효과가 apoptosis에 의한 것임을 AnnexinV-PI staining을 통 하여 확인하였다(Fig. 2). 선행연구들에서도 쿼세틴을 처 리하였을 때 여러 암종에서 암세포의 증식이 억제되었 고, 특히 HT-29 세포에서 early apoptotic cell을 인지하는 Annexin V의 발현이 증가하는 것을 확인하였으며, MCF- 7 유방암 세포에 쿼세틴을 처리하였을 때 Hoechst33342 염색을 통하여 apoptotic body가 형성됨을 확인하였다.16) 쿼세틴을 농도별, 시간별로 Western blotting을 실시한 결과 Fig. 3에서와 같이 암세포 내에서 VASP, COX-2, mTOR의 발현 억제와 AMPK의 활성이 나타나는 것을 확 인하였다. 한편 AMPK가 VASP, COX-2, mTOR 신호경로
의 조절 인자로 작용하는지를 알아보기 위해 AMPK의 저해 상태에서의 이들 단백질의 발현을 관찰하였다. Fig.
4에서는 AMPK의 저해제인 Compound C와 쿼세틴을 각 각 그리고 병행 처리한 결과, Compound C가 암세포의 증 식 억제를 저해하는 것을 보여 주고 있다. Compound C를 단독 처리한 군에서는 mTOR, COX-2, VASP의 발현이 증 가하였으며, 이 신호경로들이 AMPK에 의해 조절된다는 것을 나타내 준다. 최근 연구에 의하면, B16F10 피부암 세포에 Compound C를 처리하였을 때 mTOR와 VASP의 발현은 증가하였지만 AMPK의 발현은 감소하는 것으로 보아, AMPK가 mTOR와 VASP의 발현에 영향을 미치는 것으로 알려져 왔다.17) 선행연구와 본 실험의 결과로 AMPK가 암세포의 apoptosis를 조절함에 있어서 중심적 인 역할을 한다는 것을 알 수 있었으며 AMPK의 저해 상태에서 COX-2와 VASP가 쿼세틴에 의해 직접적으로 저해된다는 것을 확인하였다.
한편 VASP가 암세포증식에 관여하는 mTOR와 VASP 또는 COX-2와 VASP의 연관성을 알아보고자 하였다.
mTOR의 저해제인 rapamycin과 COX-2의 저해제인 cele- coxib를 각각 단독으로 그리고 쿼세틴과 병행 처리하였 을 때, 단독 처리시보다 병행 처리하였을 때 더 강하게 암세포의 증식을 억제한다는 것을 확인하였다. 동일한 조건으로 Western blotting을 실시한 결과 rapamycin에 의 하여 COX-2, VASP의 발현이 감소되었으나 AMPK의 발 현은 증가하였다. 최근 연구에서 mTOR를 저해하였을 때 COX-2의 발현이 감소하여, mTOR가 COX-2를 조절하 는 조절 인자로서 작용할 수 있음이 밝혀졌으며, mTOR 의 저해제인 rapamycin이 AMPK의 활성에 관여하는 것으 로 보고되었다.18,19) 본 연구를 통하여 mTOR의 억제로 COX-2를 조절한다는 것과, mTOR는 AMPK의 영향을 받 는 down-regulation 신호 단백질이지만 그 저해제가 AMPK 를 조절할 수도 있다는 것은 선행연구와 일치한다. 또한 mTOR가 억제된 HT-29 세포에 쿼세틴을 처리하였을 때 VASP의 발현이 더욱 감소되는 것으로 보아 mTOR의 활 성이 VASP의 발현에 영향을 미칠 수 있다는 것을 알게 되었다.
이와 같이 본 연구를 통하여 세포 증식에 관여하는 COX-2, VASP가 AMPK가 저해된 상태에서도 쿼세틴에 의해 직접 저해가 일어날 수 있다는 것을 관찰하였다.
또한 AMPK가 암세포의 apoptosis에 중심적인 역할을 하 고 있지만, 쿼세틴에 의해 AMPK-independent 신호경로로 도 COX-2, VASP의 저해가 일어날 수 있음을 확인하였 다. 결과적으로 쿼세틴이 HT-29 대장암 세포에서 AMPK 에 의존하지 않고 암세포증식에 관여하는 단백질들의
발현을 저해시켜 항암효과를 나타낼 수 있음을 밝힐 수 있었다. 끝으로 rapamycin을 통한 mTOR의 저해가 AMPK 의 활성에 영향을 미치며, VASP의 발현에 영향을 줄 수 있다는 새로운 사실을 밝힐 수 있었으며, 따라서 mTOR 와 VASP 신호경로에 관한 연구는 향후 더 진행되어야 할 연구 과제로 여겨진다.
결 론
식물 화학물질인 파이토케미컬은 세포 내 신호전달 분자들을 조절함으로써 암세포의 증식과 전이를 억제한 다고 알려져 있다. 이 중에서도 쿼세틴은 유방암, 대장암 과 같은 여러 암종에서 항암효과가 있음이 밝혀졌다. 본 연구에서는 HT-29 대장암 세포에 쿼세틴을 처리하였을 때 암세포의 증식억제 효과와 apoptosis 유도 효과를 알아 보았고, 암세포 증식에 관여하는 mTOR, VASP, COX-2의 조절양상을 확인하였다. mTOR, VASP, COX-2는 AMPK 에 의해서 조절되며, mTOR, VASP, COX-2가 암세포 내 에서 활성화되면 암의 증식을 촉진하는 것으로 알려져 있다. 이러한 암세포 증식 관련 단백질들의 상호 연관성 을 알아보기 위하여 쿼세틴을 농도별, 시간별로 처리하 였을 때, 암세포의 증식이 억제되었고 apoptosis의 유도가 일어남을 확인하였다. 또한 AMPK 억제 상태에서 쿼세 틴을 처리하였을 때, 이들 단백질의 발현이 증가되는 것 으로 보아 AMPK에 영향을 받는 것으로 나타났으나 쿼 세틴에 의한 직접적인 저해도 일어날 수 있음을 확인하 였다. 마지막으로 rapamycin과 celecoxib를 단독 그리고 쿼 세틴과 병행 처리에 의한 관계를 알아본 결과, mTOR의 저해 또는 rapamycin에 의하여 VASP의 발현에 영향을 미 칠 수 있다는 사실을 알게 되었다.
감사의 글
본 연구는 지식경제부와 한국산업기술재단의 전략기 술양성사업 및 2011년 정부(교육과학기술부)의 재원으 로 한국연구재단의 지원을 받아 수행된 연구임(KRF- 2010-0021402).
참 고 문 헌
1) Grady WM, Markowitz SD. Genetic and epigenic alterations in colon cancer. Annu Rev Genomics Hum Genet 3, 101-128, 2002.
2) Russo GL. Ins and outs of dietary phytochemicals in cancer chemoprevention. Biochem Pharmacol 74, 533-544, 2007.
3) O’Leary KA, de Pascual-Tereasa S, Needs PW, Bao YP, O’Brien NM, Williamson G. Effect of flavonoids and vitamin E on cyclooxygenase-2 (COX-2) transcription. Mutat Res 551, 245-254, 2004.
4) Kahn BB. AMP-activated protein kinase: ancient energy gauge provides clues to modern understanding of metabolism.
Cell Metab 1, 15-25, 2005.
5) Ahn J, Lee H, Kim S, Park J, Ha T. The anti-obesity effect of quercetin is mediated by AMPK and MAPK signaling pathways. Biochem Biophys Res Commun 373, 545-549, 2008.
6) Jin Q, Feng L, Behrens C, Bekele BN, Wistuba II, Hong WK, Lee HY. Implication of AMP-activated protein kinase and Akt-regulated survivin in lung cancer chemopreventive activities of deguelin. Cancer Res 67, 11630-11639, 2007.
7) Markman B, Dienstmann R, Tabernero J. Targeting the PI3K/Akt/mTOR pathway--beyond rapalogs. Oncotartet 1, 530-543, 2010.
8) Lee YK, Park SY, Kim DC, Lee WS, Surh YJ, Park OJ.
Suppression of mTOR via Akt-dependent and -independent mechanisms in selenium-treated colon cancer cells: involve- ment of AMPKalpha 1. Carcinogenesis 31, 1092-1099, 2010.
9) Brunelle M, Sartin EA, Wolfe LG, Sirois J, Doré M.
Cylooxygenase-2 expression in normal and neoplastic ca- nine mammary cell lines. Vet Pathol 43, 656-666, 2006.
10) Yu Y, Fan J, Hui Y, Rouzer CA, Marnett LJ, Klein-Szanto AJ, FitzGerald GA, Funk CD. Targeted cyclooxygenase gene (ptgs) exchange reveals discriminant isoform functio- nality. J Biol Chem 282, 1498-1506, 2007.
11) Dempke W, Rie C, Grothey A, Schmoll HJ. Cyclooxyge- nase-2: a novel target for cancer chemotheraphy. J Cancer Res 127, 411-417, 2001.
12) Yamauchi T, Watanabe M, Kubota T, Hasegawa H, Ishii Y, Endo T, Kabeshima Y, Yorozuya K, Yamamoto K, Kitajima M. Cyclooxygenase-2 expression as a new marker for patients with colorectal cancer. Dis Colon Rectum 45, 98-103, 2002.
13) Krause M, Leslie JD, Stewart M, Lafuente EM, Valderrama F, Jagannathan R, Sttrasser GA, Rubinson DA, Lui H, Way M, Yaffe MB, Boussiotis VA, Gertler FB. Lamellipodin, an Ena/VASP ligand, is implicated in the regulation of lamelli- podial dynamics. Dev Cell 7, 462-463, 2004.
14) Price CJ, Brindle NP. Vasodilatator-simulierte phosphoprotein is involved in stress-fiber and membrane ruffle formation in endothelial cells. Arterioscler Thromb Vasc Biol 20, 2051-2056, 2000.
15) Blume C, Benz PM, Walter U, Ha J, Kemp BE, Renné T.
AMP-activated protein kinase impairs endothelial actin cyto- skeleton assembly by phosphorylating vasodilator-stimulated phosphoprotein. J Biol Chem 282, 4601-4612, 2007.
16) Van der Woude H, Gliszczyńska-Swigło A, Struijs K, Smeets A, Alink GM, Rietjens IM. Biphasic modulation of cell pro- liferation by quercetin at concentrations physiologically rele- vant in humans. Cancer Lett 200, 41-47, 2003.
17) Park SY, Lee SH, Park OJ, Kim YM. Apoptotic effects of
selenium via AMPK-VASP signaling pathway in B16F10 melanoma cells. Cancer Pre Res 15, 313-319, 2010.
18) Park SY, Jung DW, Kim YM, Park OJ. Modulation of apoptosis in HepG2 cells via the negative regulation of mTOR
and COX-2 with curcumin. Cancer Pre Res 16, 118-125, 2011.
19) Habib SL, Kasinath BS, Arya RR, Vexler S, Velagapudi C.
Novel mechanism of reducing tumourigenesis: Upregulation.
Eur J Cancer 46, 2806-2820, 2010.