Carrageenan으로 염증을 유도한 Stat 6 유전자제거 생쥐의 족삼리 침치료에 대한 시상하부 유전자의
마이크로어레이 프로파일
홍미숙
1⋅박히준
2⋅엄윤경
1⋅정경희
1⋅김수철
1,3⋅한미영
1,31경희대학교 의과대학 고황의학연구소, 2한의과대학 경혈학교실, 3의과대학 소아심장분과
Microarray profile of hypothalamic gene expression with acupuncture at acupoint ST36 in carrageenan induced inflammation in Stat 6 knockout mice
Mee-Suk Hong
1, Hi-Joon Park
2, Yoon-Kyung Um
1, Kyung-Hee Jung
1, Soo-Cheol Kim
1,3, Mi-Young Han
1,31
Kohwang Medical Research Institute, College of Medicine,
2
Dept.of Meridianand Acupuncture, College of Korean Medicine,
3
Div.of Pediatric Cardiology, College of Medicine, Kyunghee University Abstract
목 적 : Signal transducers and activators of transcription 6 (Stat 6) 유전자는 면역세포의 발달 에 있어서 중요한 유전인자이며, IL‐4와 같은 사이토카인에 의해 유전자 발현이 조절된다. 본 연구에서는 Stat 6 유전자 제거 생쥐와 정상 (wild type, W/T) 생쥐에 carrageenan으로 염증을 유도한 후 족삼리에 침치료를 시행하여 시상하부에서의 유전자 발현 양상을 분석하고자 하였다.
방 법 : BALB/c (W/T, n=12) and BALB/c‐Stat 6 유전자 제거 생쥐 (n=12)의 발뒤꿈치 표피에 1%
carrageenan을 30 ul 주사하여 염증을 유도하였다. 침은 염증 유도 30분 후에 족삼리(ST36)에 시침하였으며, 염증유도에 의한 부종 증가율은 매 시간마다 측정하여 총 5시간동안 측정하였다. 마이크로어레이는 Stat 6 유전 자 제거 생쥐를 염증 유발 군과 염증유발 후 침을 처치한 군으로 나누고, 시상하부를 적출하여 RNA를 분리한 뒤. 마이크로어레이 프로파일을 분석하였다.
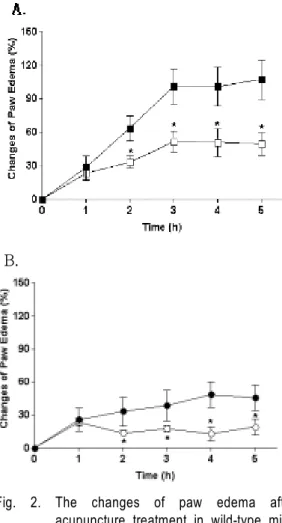
결 과 : 염증에 의한 부종증가율을 비교한 결과, Stat 6 유전자 제거 생쥐 그룹의 부종증가율이 W/T 생쥐 의 부종 증가율보다 약 50 % 정도 감소하였으며, 각 3, 4, 5시간째에 유의한 차이를 나타내었다 (각 p<0.05).
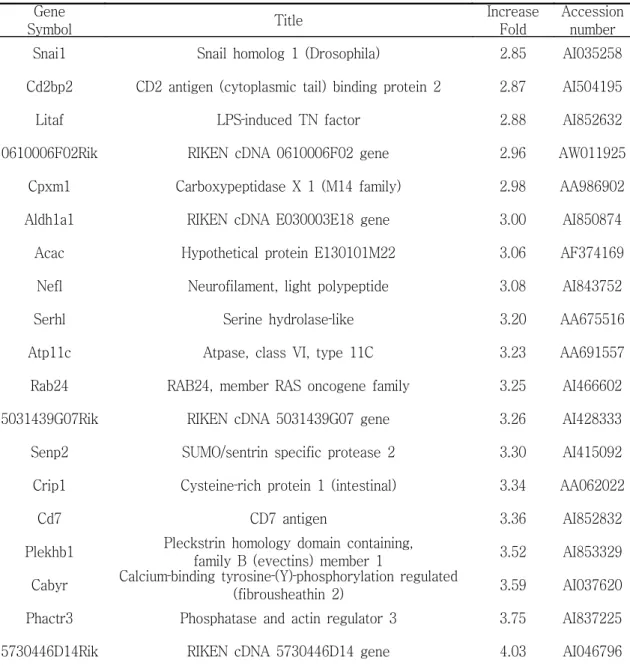
W/T생쥐군과 Stat 6 유전자 제거 생쥐군 모두에서, 침 처치군이 염증 유발 군에 비해, 염증 유발 2시간 후부터 유의한 감소를 나타내었다. 시상하부의 유전자 발현을 관찰한 결과, 39개의 유전자가 3배 이상 감소하였으며, 19 개의 유전자는 3배 이상 증가하였다.
결 론 : W/T 생쥐군과 Stat 6 유전자 제거 생쥐 모두에서 침의 진통효과는 나타나며, 이의 기전에는 시상 하부에서의 침 치료에 의한 염증관련 유전자들의 감소와, 항염증과 관련된 유전자들이 증가가 관여하는 것으로 보인다.
Key words : Stat 6 knockout mice, carrageenan, hypothalamus, microarray, ST36, acupuncture
I. Introduction
Signal transducer and activator of
⋅교신저자 : 홍미숙, 서울특별시동대문구회기동 1번지 경희대학교 의 과대학 약리학교실, Tel. 02-961-0303, Fax. 02-968-0560, E-mail: [email protected]
⋅본 연구는 과학기술부/한국과학재단 우수연구센터육성사업의 지 원으로 수행되었음(R11-2005-014).
⋅투고 : 2007/05/28 심사 : 2007/06/07 채택 : 2007/06/14