815
Copyright © 2016 The Korean Society of Fisheries and Aquatic Science pISSN:0374-8111, eISSN:2287-8815
서 론
높은 생체적합성을 장점으로 최근들어다양한생체 중합 체가의료 및 제약분야의 응용에널리 연구되어 지고 있다
(Niaounakis, 2015; Pomastowski et al., 2016; Tham and Yi,
2015).
특히키토산은이들중에서가장활발히연구가진행되고있는물질중의하나로키틴의탈아세틸화
(deacetylation)
에 의해유도된양이온을띤다당류중합체이다(Kim et al., 2008;
Vinsova and Vavrikova, 2008).
키틴은지구상에서자연적으 로발생하는가장풍부한생체중합체중의하나로,
해양및수 산자원의부산물인게(Brachyura),
새우(Alpheoidea)
및가재(Nephropidae)
등갑각류(Crustacean)
의껍질에서쉽게얻어진 다.
키틴은낮은수용성으로산업적응용이제한적이지만,
키토 산을비롯하여수용성이큰다양한키틴의유도체가활발히연 구되어보다유용한자원이되었다.
특히키토산은수용성이크 고,
천연물질에서유도되어생체적합성이커서약물및약물전달등에다양하게연구되어지고있다
(Taveira et al., 2009).
키틴은
N-
아세틸-D-
글루코사민(N-acetyl-D-glucosamine)
단위(unit)
가여러개모여선형의긴사슬을이루는데,
이키틴의탈 아세틸화에의해얻어진키토산분자는여러개의아미노기를 가지게된다(Kumar et al., 2005).
이들아미노기는pKa ≈ 6.5
로 산성도및환경에따라양성자화(protonation)
를통해키토산분 자를전기적으로양성을띠게만들고,
이러한물리적특성이키토산의수용성을더욱증가시켜물에잘녹게한다
(Krajewska
et al., 2011).
최근키토산과키토산의항균작용에대한연구가활발하였 으나
,
원자수준에서의항균작용의기작에대한연구는여전히 발표된바가없다(Choi et al., 2001; Taha and Swailam, 2002;
Tsai and Su, 1999).
따라서우리는전기적으로양전하를띠는 키토산올리고당분자가음전하를띠는세균막(bacterial mem-
brane)
과정전기적인력으로인해전기적으로중성인정상세포막은파괴하지않고선택적으로세균의세포막만을파괴한
키토산의 양성자화가 세균막과의 상호작용에 미치는 효과 연구
Gia Khuong Truong·이명기
1*
부경대학교 의생명기계전기융합공학협동과정, 1부경대학교 의공학과
Molecular Dynamics Simulation Studies of the Effects of the Protonation State of Chitosan in Interactions with Bacterial Membranes
Gia Khuong Truong and Myunggi Yi
1
*Interdisciplinary Program of Biomedical, Electrical and Mechanical Engineering, Pukyong National University, Busan 48513, Korea
1
Department of Biomedical Engineering, Pukyong National University, Busan 48513, Korea
The cationic biopolymer chitosan has several applications in medicine. Chitosan is the deacetylated derivative of chitin, the second most abundant naturally occurring polymer. Recent studies have investigated the relationship between chitosan and antibacterial activity. However, the molecular interactions and mechanisms have not been de- tailed. This study used molecular dynamics simulations to study interactions between chitosan and anionic bacterial membranes (POPE-POPG) and electrically neutral non-bacterial membranes (POPC). We calculated the free energy using umbrella sampling to compare the interactions between membranes and chitosan in different protonation states.
Fully protonated chitosan interacted most strongly with the bacterial membranes, but weakly with non-bacterial membranes. These results suggest that electrostatic interactions are the main mechanism of the antibacterial activity of chitosan, and they provide insights into the design of novel antibacterial and antimicrobial agents.
Key words: Chitosan, Antibacterial activity, Molecular dynamics, Electrostatic interaction
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial Licens (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
http://dx.doi.org/10.5657/KFAS.2016.0815 Korean J Fish Aquat Sci 49(6) 815-822, December 2016
Received 31 October 2016; Revised 30 November 2016; Accepted 5 December 2016
*Corresponding author: Tel: +82. 51. 629 .5773 Fax: +82. 51 .629 .5779
E-mail address: [email protected]
다는가설에따라
(Matsuzaki, 2009),
이들키토산올리고당분 자와세포막 사이의상호작용을 원자수준에서 해석하고 키 토산의항균작용의기작을이해하고자분자동역학전산모사(molecular dynamics simulations)
를이용한자유에너지(free energy)
계산을수행하였다.
재료 및 방법
전산모사 재료(Simulation systems)
본연구에서는다양한환경과가능성을고려하기위해키토산 올리고당의양성자화정도와세포막의종류에따라총
4
개의 분자동역학 전산모사를 수행하였다.
키토산올리고당 분자는 양성자화의정도에따라전부양성자화(fully protonated)
되거 나반만양성자화(half protonated),
그리고탈양성자화(deprot- onated)
된키토산으로총3
종류를사용하였다.
이후로이3
종 류의키토산올리고당분자를각각fully, half
그리고depro
로표 시하겠다.
세포막은두종류의인지질이중막(phospholipid bi- layer)
으로세균막은1-palmitoyl-2-oleoyl-sn-glycero-3-phos-
phoethanolamine (POPE)
지질 분자와 전기적으로 음전하를 띠는
1-palmitoyl-2-oleoyl-sn-glycero-3-phosphoglycerol
(POPG)
지질분자의조합으로구성된지질이중막으로모사되었고
,
일반세포막은전기적으로중성인1-palmitoyl-2-oleoyl- sn-glycero-3-phosphocholine (POPC)
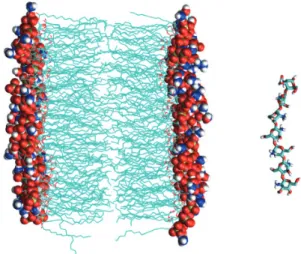
단일지질분자로구성 된지질이중막으로모사되었다(Picas et al., 2010a; Picas et al., 2010b) (Fig. 1).
키토산
본연구에서는
1
개의N-
아세틸-D-
글루코사민(
키틴)
과6
개 의D-
글루코사민(
키토산)
으로이루어진86%
의탈아세틸화된 키토산올리고당을사용하였고,
양성자화정도에따라3
종류 의 키토산올리고당을PRODRG
서버(http://davapc1.bioch.
dundee.ac.uk/cgi-bin/prodrg)
를 이용한3
차원 구조를모델링 에사용하였다(Schuttelkopf and van Aalten, 2004). Table 1
에 양성자화의정도에따른3
종류의키토산올리고당잔기서열을 각각나타내었다. Fully
의경우6
개의키토산잔기를모두양성 자화시켜총+6 e (e
는전자의전하량으로1.6×10
-19C
이다.)
의전하량을가지고, half
의경우2, 4,
및6
번의잔기에양성자 화를할당해양전하끼리의척력을고려하였으며,
총+3 e
의전하량을띠며
, depro
의경우전하를띠지않아전기적으로중성분자가된다
.
따라서본연구에사용된키토산올리고당의분자 량은각각1,175, 1,172, 1,169 g/mol
이된다.
지질 이중막(세포막)
세균의세포막은복잡한다층구조로되어있어현재까지분자 동역학전산모사로자세하게구현하지못하지만
,
최근에는보 다간단한음전하를띤지질이중막모형으로여러연구에서좋 은결과를얻고있다(Parkin et al., 2015; Silhavy et al., 2010).
따라서본연구의목적인키토산과세균막과의정전기적상호 작용을연구하기엔간단한지질이중막으로충분하다고고려 되었다
.
전산모사의수행속도를높이기위해이미열역학적 평형상태에 이른(pre-equilibrated) 128
개의POPC
와POPG
지질분자로이루어진충분히수화된두가지의이중막
(fully
hydrated bilayer)
을 사용하여 전산모사를 시작했다(Elmore, 2006).
일반세포막은전기적중성을띤128
개의POPC
단일 종류의지질분자로이루어진이중막을사용하였고, 128
개의POPG
지질분자로이루어진이중막을조금변형하여세포막을모사하기위해사용되었다
. Fig. 2
에전산모사에서사용된인 지질(phospholipid)
분자들의화학구조와그구성성분을나타 내었는데,
분자들사이차이는지질의머리부분뿐이므로무작 위로96
개POPG
분자들의글리세롤을POPE
지질의에탄올아민으로대체하여
3:1
비율의POPE-POPG
지질이중막을구성하여총
-32 e
의음전하를띤세균막을모사하였다(Hallock
et al., 2002; Hong et al., 2014; Mani et al., 2006).
막의구성이 Table 1. Sequences of three kinds of chitosan models according to their protonation statesChitosan Residue 0 1 2 3 4 5 6
fully protonated n1 p2 p p p p p
half protonated n d3 p d p d p
deprotonated n d d d d d d
1n, N-acetyl-D-glucosamine.2p, protonated D-glucosamine. 3d, de- protonated D-glucosamine.
Fig. 1. Molecular dynamics simulation system. Lipid bilayer is represented by cyan colored lines for fatty acyl chains, and by spheres for head groups of lipid molecules. Chitosan is represented by cyan (carbon), red (oxygen), blue (nitrogen), and white (hydro- gen) colored sticks. Water molecules are not displayed for clarity.
달라졌기때문에새지질이중막
(POPE-POPG)
을다시평형상 태에이르도록추가로73.06 nanoseconds (ns)
동안전산모사 를수행하였다.
분자동역학 전산모사(Molecular dynamics simula- tions)
첫번째전사모사에는먼저
1
개의키토산올리고당분자fully
와 앞서 준비한
POPE-POPG
지질 이중막(POPE 96
개와POPG 32
개로이루어진)
을포함한시스템으로전산모사로시작하였다
.
이들키토산올리고당과지질이중막은162
개의Na
+ 이온과136
개의Cl
-이온을포함한10,403
개의SPC
물분자 모델(Berweger et al., 1995)
의상자안에수용되었고,
이온들은 전체시스템을전기적으로중성으로만들면서,
최종적으로이 온농도를150 mM
로만들었다.
이어서52.06 ns
동안전산모 사를수행하여시스템을충분히평형상태에이르게하였다.
이 평형상태에이른시스템의키토산올리고당분자를수정하여half
와depro
로대체하고각각3
개및6
개의Na
+이온을제거 한후모두3
개의전산모사시스템이준비되었다.
비슷한방 법으로키토산올리고당과일반세포막과의상호작용을연구하기위해
fully
와POPC
지질이중막을포함한전산모사시스템을준비하였다
.
모든 분자동역학 전산모사는
GROMACS 5.0.4 (Van Der Spoel et al. 2005)
를이용하였으며, force field
로gromos53a6 (Oostenbrink et al., 2004)
를사용하였다.
전산모사는먼저시 스템을준비하면서발생할수있는bad contact
를제거하기위해
5000
단계의energy minimization
으로시작되었다.
이후60 picoseconds (ps)
동안점진적으로온도를10 K
에서310 K
까지 등적(constant volume)
조건에서가열후,
마지막으로등압등 온(constant pressure and temperature)
조건으로전환하여시스 템의밀도를평형상태에이르게하였다. Newton
운동방정식의 적분간격은2 femtosecond
로하였고
,
모든원자들의좌표는10 ps
마다trajectory
로저장되어결과분석에사용되었다.
시 스템의평균온도와압력은각각310 K
와1 bar
로유지되었고,
근거리정전기력과판데르발스상호작용은1.2 nm
의cutoff
를사용하였고,
원거리정전기력은주기적경계조건(periodic boundary condition)
과함께particle-mesh Ewald (PME)
방법 으로다루었다(Essmann et al., 1995; Toukmaji et al., 2000).
자유 에너지 계산(free energy calculations)
키토산올리고당의전하량에따른세포막과의상호작용의차 이를분석하기위해키토산올리고당의양성자화정도와지질이 중막사이의거리에따라자유에너지(Gibbs free energy)
가어 떻게변하는지계산해보았다.
반응좌표(reaction coordinate)
로지질이중막의 중심에서막과물의경계면에 수직으로키 토산의중심까지거리를선택하고, umbrella sampling (Torrie and Valleau, 1977)
을이용해전산모사를수행하였다.
지질막과키토산의거리를
0.2 nm
의간격으로각시스템마다총14
개의
window
를만들어힘상수로500 kJ/mol/nm
2을가하여전 산모사를 수행하였다.
총~7 µs
의전산모사를수행하였는데,
각각의
window
마다자유에너지가수렴할때까지최소120
ns
의전산모사를수행하여마지막60 ns
동안의trajectory
가weighted histogram analysis method (WHAM)
방법으로자유 에너지를계산하는데이용되었다(Hub et al., 2010).
결 과
Free energy profile
전산모사로부터얻어진자유에너지는또한
potential of mean
force (PMF)
로일컬어지는데,
양성자화정도에따른각키토산올리고당과지질이중막사이거리에따라
PMF
가어떻게변해가는지계산결과를
Fig. 3
에나타내었다.
또한각시스템마다지질이중막의위치를나타내기위해반응좌표를따라각지질 분자를구성하는그룹들의수밀도
(partial number density)
를 계산하여그래프에표시하였다.
수밀도계산을위해지질분자 그룹중에서특정원자들이선택되었는데Fig. 2
에붉은원으로 나타내었다.
이들원자들은각그룹의위치를대표하고또한전 기적으로전하를띠거나극성을띠고있어키토산올리고당과 의상호작용에서수소결합등을제공할수있기때문에선택 되었다.
그래프의수평축은반응좌표로지질이중막평면에대 해수직으로키토산혹은지질구성분자그룹들과의거리로, 0
값은지질이중막의중심을나타낸다.
그래프의좌측수직축에 Fig. 2. Lipid molecules and component groups of each lipid. Thecircles with red broken line indicate the selected atoms for number density calculations in Fig. 3.
는자유에너지를나타내었는데
,
자유에너지0
값은키토산올 리고당분자가지질이중막으로부터멀리떨어져더이상상호 작용이없고주위물분자들과의상호작용만있을때,
즉PMF
가거리에따라변하지않고평평하게될때를기준으로하였다.
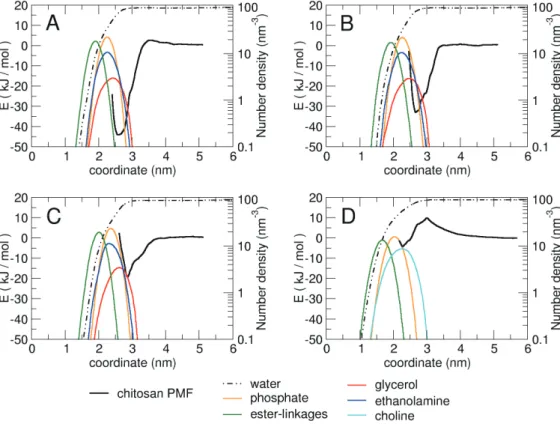
Fig. 3ABC
에서3
종류의양성자화정도에따라키토산올리고당과세균막과의따라자유에너지의변화를볼수있는데
,
모 든키토산올리고당분자들이공통적으로지질막의중심으로부 터약3.5 nm
이상멀어지면서자유에너지가0 kJ/mol
로접근 하기시작하면서약5.0 nm
에이르면완전히수렴하는것을알 수있다.
키토산올리고당이3.5 nm
보다가까워지면서부터세 균막과인력으로인해자유에너지가낮아지면서3
종류의양성 자화에따라서로다른모양을나타내는것을알수있는데,
공 통적으로는지질막중심으로부터약2.5-3.0 nm
떨어진세균막 의표면에서자유에너지가최저가되었다. fully
의경우+6 e
의전하를띠고있어서음전하를띤지질이중막
(POPE-POPG)
의한쪽단일막과
(-16 e)
의정전기적인력이가장강해자유에너지가약
3.5 nm
부터가파르게감소를시작해2.6 nm
에서최저-44.0 kJ/mol
까지낮아졌다(Fig. 3A).
수밀도(number density)
로부터지질구성그룹들의막내에서의분포를알수있는데,
두지질분자들의음전하를띤인산그룹(Phosphate)
과POPE
지질분자의양전하를띤에탄올아민그룹이같은위치에분포
되고
POPG
지질분자의전하를띠지않은글리세롤그룹은이들보다조금더세균막바깥쪽에분포하는것을알수있다
.
자 유에너지가가장낮은곳은지질분자들의글리세롤그룹이분 포하면서에탄올아민과인산그룹의분포가시작되는부분이 다.
하지만키토산올리고당이세균막좀더깊은곳으로접근하 면인산과에탄올아민의밀도가본격적으로커지고에스터결 합(ester-linkage)
의분포가시작되면서자유에너지가다시가 파르게증가하는것을알수있다.
이곳에서부터는정전기적인 력보다지질분자들의밀도가높아지면서키토산올리고당이막 내부로더깊이들어갈수록물리적인(physical)
저항이커지는 것으로해석된다.
half
의경우자유에너지를제외하고는물과지질분자의분포 등은fully
와거의동일한모양의그래프를보여준다(Fig. 3B).
+3 e
의전하를띤half
의경우2.7 nm
에서최저자유에너지-33.0 kJ/mol
로fully
의경우보다높아서정전기적인력이상대적으로약한것으로나타났다
. fully
의경우와마찬가지로세균막과가까워지면서표면의전하를띠거나전기적극성을띠 는지질분자의머리부분과정전기적상호작용을하면서자유
에너지가낮아지기시작한다
.
자유에너지최저점의위치는0.1
Fig. 3. Free energy profiles and partial number densities of water and component groups of lipid molecules along the reaction coordinate of (A) fully protonated chitosan, (B) half protonated chitosan, (C) deprotonated chitosan with bacterial membrane, (D) fully protonated chitosan with non-bacterial membrane.
nm
지질이중막의중심에서보다멀어졌고,
키토산올리고당이 지질막으로보다깊이들어갈수록자유에너지는급격하게증 가하는것을볼수있었다.
전하를띠지않는
depro
의경우또한자유에너지최저값을제외하고는
fully
및half
와거의동일한모양의그래프를보여준 다(Fig. 3C).
전기적으로중성인depro
의경우2.9 nm
에서최 저자유에너지는-19.2 kJ/mol
로half
의경우보다도더높아 서키토산올리고당과세균막과의인력이가장약한것으로나타났다
.
선호하는결합위치도지질이중막중심에서2.9 nm
로가장멀리떨어져있다
.
여기서전기적으로중성을띠지만키토 산올리고당과세균막사이여전히인력이작용함을볼수있는 데,
이것은총전하는0
이지만앞의두경우와마찬가지로키 토산올리고당은여전히전기적극성들띤친수성(hydrophilic)
분자이고,
이분자와전하를띠거나전기적극성인지질분자 의머리부분과는정전기적으로상호작용하면서인력이작용 하기때문이다.
전기적으로중성이지만극성을띠는지질분자 의글리세롤이분포한곳에서자유에너지최저점을지나고전 하를띠는인산과에탄올아민의수가증가하는곳부터자유에 너지가가파르게증가하면서인력에서척력으로전환되는것 을알수있다.
전산모사의결과에서키토산올리고당과세균막이강한인력 으로상호작용하는것을확인하였는데
,
마지막으로세균막이 아닌경우,
즉키토산올리고당(fully)
이일반적인세포막과어 떻게상호작용하는지자유에너지계산을통해비교해보았다(Fig. 3D). POPC
지질이중막의총전하는0
으로전기적으로 중성이지만지질분자의머리부분에콜린(choline)
및인산등전기적극성을띠는그룹을포함하는것은
POPE-POPG
지질이중막과비슷하다
.
그래프에서보듯이키토산올리고당과일 반세포막사이에서는오히려척력이작용하는것을확인할수 있었다.
자유에너지최저점은2.3 nm
에서-4.1 kJ/mol
로모든 경우에서가장약한인력이었고,
이최저점에도달하기위해서 키토산올리고당은3.0 nm
에존재하는9.9 kJ/mol
의에너지장 벽을넘어야한다.
따라서키토산올리고당은일반세포막표면 에결합하는것을선호하지않음을알수있다.
척력의원인은 정전기적상호작용때문인데,
전기적으로음성인인산보다양 성인콜린그룹이보다세포막표면의바깥쪽에분포하기때문 이다(Yi et al., 2008).
에너지장벽을통과하면서전기적으로음 성인인산그룹과인력이작용하기시작하면서자유에너지는 감소하기시작하고,
최저점을지나면서다시지질분자들의밀 도가높아지면서물리적인저항으로인해자유에너지가증가 하기시작한다.
수소 결합(Hydrogen bond interactions)
우리는자유에너지그래프를보다깊이이해하기위해반응 좌표에따라키토산올리고당과물분자그리고지질분자머 리부분의수소결합이어떻게달라지는가를조사하였다
(Fig.
4).
본계산에서는키토산올리고당,
물분자,
지질분자의질소 와산소원자들만수소결합의도너(donor)
와억셉터(acceptor)
로고려하였고,
도너와억셉터사이의거리가0.34 nm
이하이 고도너-
수소-
억셉터가이루는각도가150°
이상인경우만수소 결합으로간주하였다.
반응좌표인지질이중막과 키토산올리고당의거리에따라
0.02 nm
간격으로전산모사동안키토산올리고당과물분자그리고키토산올리고당과지질분자사이의평균수소결합의개 수를측정하였다
. Fig. 4A
로부터fully
의경우평균적으로약9
개의수소결합을하는것을알수있는데,
먼저세균막으로부 터멀리떨어져물속에있을경우지질분자와수소결합은없 고순수하게물분자들과만수소결합을한다.
점점지질막표면 으로다가가면서물분자의밀도는떨어지면서(Fig. 3)
수소결 합의수가감소한다.
반면에지질분자와의수소결합이증가하 면서총수소결합의개수는비슷하게유지된다.
이수소결합의 개수는키토산올리고당의양성자화정도와비례하는것을알 수있었는데, half
의경우약7
개였고(Fig. 4B), depro
의경우 약6
개로(Fig. 4C)
줄어들뿐모두비슷한경향의변화를보였 다.
자유에너지계산과마찬가지로양성자화정도와결합에너 지강도가비례하는것이수소결합의개수에서도나타났다.
키 토산올리고당과일반세포막과의전산모사에물분자와의수소결합은
fully
의경우와비슷한경향을보였는데,
지질분자와의수소결합의개수는
fully
의경우만큼증가하지않는것을볼수있었다
.
키토산올리고당과지질분자의수소결합중에서음전하를띤인산그룹
(Fig. 2)
과의비율이결합에너지및양성자화정도와비례하는또하나의수치로확인되었다
.
키토산올리고 당이지질분자의머리부분중에서인산그룹과의수소결합의 비율은전체수소결합에서74.6% (fully), 61.1% (half), 36.2%
(depro)
로각각나타났다.
고 찰
항균 작용의 기작(Mechanism of antibacterial ac- tivity)
전산모사
trajectory
중에서키토산올리고당과지질이중막의상호작용을분자수준에서자세하게보여주는대표적인
snap- shot
하나를Fig. 5
에나타내었다.
화학적항균물질의작용기 작은너무나다양하지만,
키토산올리고당의경우비슷한생체 중합체인항생펩티드(antimicrobial peptide, AMP)
의항생작용으로세포막에기공을형성하는
(pore forming)
모델및세포막의표면에결합하는카펫
(carpet)
모델등과비교할만하다(Wimley, 2010).
전산모사의결과에따르면키토산올리고당의경우특히카펫
(carpet)
모델(Gazit et al., 1996)
에가까운것으 로해석된다(Fig. 5).
키토산올리고당이AMP
기공형성모델처 럼소수성의지질이중막내부를관통하여채널형태를구성하 며세포막에구멍을뚫는것으로보기에는친수성이너무강하다
.
자유에너지계산결과로부터키토산올리고당이세균막의 표면에강하게결합하는것을알수있지만,
지질분자의친수성머리부분을지나면소수성이강한지방산사슬
(Fig. 2)
로이루어진이중막내부로들어가기위해탈수화비용
(desolvation
cost)
이너무크게된다.
아마도카펫모델처럼세균막의표면에결합하여막에직접적으로영향을주거나
,
막단백질의물질수 송이나내부에간접적으로영향을주어서세포를파괴하는것 으로예상된다(Pouny et al., 1992).
키토산은해양및수산산업의부산물로생체적합성이높고 항균기능이있어다양한식품및의약품분야에서연구중이지 만최근까지그항균작용의기작에대한원자수준의이해에는 이르지못하였다
.
우리는본연구에서처음으로분자동역학전 산모사를통해키토산의항균작용기작을원자수준에서규명 하였다.
양전하를띠는키토산은음전하를띠는세균막과강하 게결합하는반면,
전기적중성의일반세포막과는약하게상호 작용하는것을알수있었고,
이는기존의실험적연구들과잘 일치하고있다(Gottenbos et al., 2001; Matsuzaki, 2009).
본연 구를통한원자수준에서의항균작용기작의이해를바탕으 로성능이향상된항생혹은항균물질개발에도움이될것으 로기대된다.
사 사
이논문은부경대학교자율창의학술연구비
(2015
년)
에의하 여연구되었음References
Berweger CD, van Gunsteren WF and Müller-Plathe F. 1995.
Fig. 4. Number of hydrogen bonds between chitosan and water, chitosan and lipid. (A) fully protonated chitosan, (B) half protonated chito- san, (C) deprotonated chitosan with bacterial membrane, (D) fully protonated chitosan with non-bacterial membrane.
Fig. 5. Atomic detail of interactions between fully protonated chitosan and anionic bacterial membrane. POPE and POPG and lipid molecules are represented by lines and colored by orange and cyan, respectively. Chitosan molecule is represented the same way as shown in Fig. 1. Hydrogen bonds are represented by purple bro- ken lines.
Force field parametrization by weak coupling. Re-engineer- ing SPC water. Chem Phys Lett 232, 429-436. http://dx.doi.
org/10.1016/0009-2614(94)01391-8.
Choi B-K, Kim K-Y, Yoo Y-J, Oh S-J, Choi J-H and Kim C-Y.
2001. In vitro antimicrobial activity of a chitooligosaccha- ride mixture against Actinobacillus actinomycetemcomitans and Streptococcus mutans. Int J Antimicrob Agents 18, 553- 557. http://dx.doi.org/10.1016/S0924-8579(01)00434-4.
Elmore DE. 2006. Molecular dynamics simulation of a phos- phatidylglycerol membrane. FEBS letters 580, 144-148.
http://dx.doi.org/10.1016/j.febslet.2005.11.064.
Essmann U, Perera L, Berkowitz ML, Darden T, Lee H and Ped- ersen LG. 1995. A smooth particle mesh Ewald method. J Chem Phys 103, 8577. http://dx.doi.org/10.1063/1.470117.
Gazit E, Miller IR, Biggin PC, Sansom MSP and Shai Y.
1996. Structure and orientation of the mammalian anti- bacterial peptide cecropin P1 within phospholipid mem- branes. J Mol Biol 258, 860-870. http://dx.doi.org/10.1006/
jmbi.1996.0293.
Gottenbos B, Grijpma DW, van der Mei HC, Feijen J and Buss- cher HJ. 2001. Antimicrobial effects of positively charged surfaces on adhering Gram-positive and Gram-negative bacteria. J Antimicr Chemother 48, 7-13.
Hallock KJ, Lee D-K, Omnaas J, Mosberg HI and Ramamoor- thy A. 2002. Membrane composition determines Pardaxin's mechanism of lipid bilayer disruption. Biophys J 83, 1004- 1013. http://dx.doi.org/10.1016/S0006-3495(02)75226-0.
Hong C, Tieleman DP and Wang Y. 2014. Microsecond mo- lecular dynamics simulations of lipid mixing. Langmuir 30, 11993–12001. http://dx.doi.org/10.1021/la502363b.
Hub JS, de Groot BL and van der Spoel D. 2010. A free weight- ed histogram analysis implementation including robust er- ror and autocorrelation estimates. J Chem Theory Comput 6, 3713-3720. http://dx.doi.org/10.1021/ct100494z.
Kim I-Y, Seo S-J, Moon H-S, Yoo M-K, Park I-Y, Kim B-C and Cho C-S. 2008. Chitosan and its derivatives for tissue engi- neering applications. Biotechnol Adv 26, 1-21. http://dx.doi.
org/10.1016/j.biotechadv.2007.07.009.
Krajewska B, Wydro P and Jańczyk A. 2011. Probing the modes of antibacterial activity of chitosan. Effects of pH and mo- lecular weight on chitosan interactions with membrane lip- ids in Langmuir films. Biomacromolecules 12, 4144-4152.
http://dx.doi.org/10.1021/bm2012295.
Kumar RMNV, Muzzarelli RAA, Muzzarelli C, Sashiwa H and Domb AJ. 2005. Chitosan chemistry and pharmaceuti- cal perspectives. ChemInform 36, 6017-6084. http://dx.doi.
org/10.1002/chin.200511296.
Mani R, Cady SD, Tang M, Waring AJ, Lehrer RI and Hong M.
2006. Membrane-dependent oligomeric structure and pore formation of a β-hairpin antimicrobial peptide in lipid bi- layers from solid-state NMR. Proc Natl Acad Sci USA 103, 16242-16247. http://dx.doi.org/10.1073/pnas.0605079103.
Matsuzaki K. 2009. Control of cell selectivity of antimicrobial peptides. Biochimica et biophysica acta 1788, 1687-1692.
http://dx.doi.org/10.1016/j.bbamem.2008.09.013.
Niaounakis M. 2015. Biopolymers: Applications and trends. El- sevier, Amsterdam. http://dx.doi.org/10.1016/B978-0-323- 35399-1.09001-3.
Oostenbrink C, Villa A, Mark AE and Gunsteren WF. 2004. A biomolecular force field based on the free enthalpy of hy- dration and solvation: The GROMOS force-field parameter sets 53A5 and 53A6. J Comput Chem 25, 1656-1676. http://
dx.doi.org/10.1002/jcc.20090.
Parkin J, Chavent M and Khalid S. 2015. Molecular Simula- tions of Gram-Negative Bacterial Membranes: A Vignette of Some Recent Successes. Biophys J 109, 461–468. http://
dx.doi.org/10.1016/j.bpj.2015.06.050.
Picas L, Carretero-Genevrier A, Montero TM, Vázquez-Ibar JL, Seantier B, Milhiet P-E and Hernández-Borrell J. 2010a.
Preferential insertion of lactose permease in phospholipid do- mains: AFM observations. BBA-Biomembranes 1798, 1014- 1019. http://dx.doi.org/10.1016/j.bbamem.2010.01.008.
Picas L, Montero TM, Morros A, Vázquez-Ibar JL and Hernán- dez-Borrell J. 2010b. Evidence of phosphatidylethanol- amine and phosphatidylglycerol presence at the annular region of lactose permease of Escherichia coli. BBA-Bio- membranes 1798, 291-296. http://dx.doi.org/10.1016/j.
bbamem.2009.06.024
Pomastowski P et al. 2016. Silver-Lactoferrin Nanocomplexes as a potent antimicrobial agent. J Am Chem Soc 138, 7899–
7909. http://dx.doi.org/10.1021/jacs.6b02699.
Pouny Y, Rapaport D, Mor A, Nicolas P and Shai Y. 1992. In- teraction of antimicrobial dermaseptin and its fluorescently labeled analogues with phospholipid membranes. Biochem- istry 31, 12416-12423.
Schuttelkopf AW and van Aalten DM. 2004. PRODRG: a tool for high-throughput crystallography of protein-ligand com- plexes. Acta Crystallogr Sect D-Biol Crystallogr 60, 1355- 1363. http://dx.doi.org/10.1107/S0907444904011679.
Silhavy TJ, Kahne D and Walker S. 2010. The bacterial cell envelope. Cold Spring Harbor perspectives in biology 2, a000414. http://dx.doi.org/10.1101/cshperspect.a000414.
Taha SMA and Swailam HMH. 2002. Antibacterial activ- ity of chitosan against Aeromonas hydrophila. Food / Nahrung 46, 337-340. http://dx.doi.org/10.1002/1521-3803 (20020901)46:5<337::AID-FOOD337>3.0.CO;2-3.
Taveira SF, Nomizo A and Lopez R. 2009. Effect of the ionto- phoresis of a chitosan gel on doxorubicin skin penetration and cytotoxicity. J Control Release 134, 35-40. http://dx.doi.
org/10.1016/j.jconrel.2008.11.002.
Tham PTH and Yi M. 2015. Selectivity of the α6 nAChR sub- unit on α-conotoxin BuIA studied by molecular dynamics simulations. Korean J Fish Aquat Sci 48, 071-075. http://
dx.doi.org/10.5657/KFAS.2015.0071.
Torrie GM and Valleau JP. 1977. Nonphysical sampling dis- tributions in Monte Carlo free-energy estimation: Um- brella sampling. J Comput Phys 23, 187-199. http://dx.doi.
org/10.1016/0021-9991(77)90121-8.
Toukmaji A, Sagui C, Board J and Darden T. 2000. Efficient particle-mesh Ewald based approach to fixed and induced dipolar interactions. J Chem Phys 113, 10913. http://dx.doi.
org/10.1063/1.1324708.
Tsai GJ and Su WH. 1999. Antibacterial activity of shrimp chi- tosan against Escherichia coli. J Food Prot 62, 239-243.
Van Der Spoel D, Lindahl E, Hess B, Groenhof G, Mark AE and Berendsen HJ. 2005. GROMACS: fast, flexible, and free. J Computat Chem 26, 1701-1718. http://dx.doi.org/10.1002/
jcc.20291.
Vinsova J and Vavrikova E. 2008. Recent advances in drugs and prodrugs design of chitosan. Curr Pharm Des 14, 1311- 1326. http://dx.doi.org/10.2174/138161208799316410.
Wimley WC. 2010. Describing the mechanism of antimicrobial peptide action with the interfacial activity model. ACS Chem Biol 5, 905-917. http://dx.doi.org/10.1021/cb1001558.
Yi M, Nymeyer H and Zhou H-X. 2008. Test of the Gouy-Chap- man theory for a charged lipid membrane against explicit- solvent molecular dynamics simulations. Phys Rev Lett 101, 38103. http://dx.doi.org/10.1103/PhysRevLett.101.038103.