한국, 일본, 중국의 전통 미생물발효차(청태전, 아와반자, 보이차)의 미생물 군집 비교
김병혁1,2․장종옥2․이중복1․고상욱2․문두경2
1교촌 F&B(주) 교촌중앙연구소
2농촌진흥청 국립원예특작과학원 온난화대응농업연구소
Microbial Diversity Comparison between Korean, Japanese, and Chinese Post-Fermented Tea (Chungtaejeon, Awabancha, and Puerh tea)
Byung-Hyuk Kim1,2, Jong-Ok Jang2, Jung-Bok Lee1, Sangwook Koh2, and Doo-Gyung Moon2
1Kyochon Research & Innovation Center, Kyochon F&B Co., Ltd.
2Agricultural Research Institute for Climate Change, National Institute of Horticultural and Herbal Science, RDA
ABSTRACT Post-fermented tea was produced by a microbial fermentation process using sun-dried green tea leaves (Camellia sinensis (L.) O Kuntze) as the raw material. In this study, bacterial community analysis involved in the production of East-Asia traditional post-fermented tea (Korean Chungtaejeon, Japanese Awabancha, and Chinese Puerh tea) were investigated using double gradient-denaturing gradient gel electrophoresis (DG-DGGE), 16S rRNA gene clone library, and quantitative real-time PCR through 16S rRNA gene analysis. The dominant microorganisms of Chungtaejeon, Awabancha, and Puerh tea were identified as Pantoea sp., Lactobacillus sp., and Sphingomonas sp., respectively.
Interestingly, the dominant microorganism presents in the Korean and Japanese post fermented teas were a bacterium, while it was a fungus for the Chinese post-fermented tea.
Key words: Awabancha, Chungtaejeon, microbial community, post-fermented tea, Puerh tea
Received 18 December 2019; Accepted 13 February 2020 Corresponding author: Doo-Gyung Moon, Agricultural Research Institute for Climate Change, National Institute of Horticultural and Herbal Science, RDA, Jeju-si, Jeju-do 63240, Korea
E-mail: [email protected], Phone: +82-64-741-2590
서 론
차는 대부분의 나라에서 소비되고 있으며, 한국, 중국, 일 본 등의 동아시아 지역에서 오랜 역사 동안 널리 음용되어 왔다. 다년생 상록관목인 차나무(Camellia sinensis (L.) O Kuntze)는 한국, 중국, 일본, 인도, 스리랑카를 비롯한 아시 아, 아프리카, 러시아 등의 아열대 지역과 온대 지역에 걸쳐 넓게 재배되고 있으며, 차 추출물에는 caffeine, amino acid, polyphenol(catechins) 등의 다양한 성분을 함유하고 있다 (Ho 등, 2008). 최근 고도화된 산업화와 더불어 건강에 대한 관심이 증가하고 차 추출물의 다양한 건강증진 효과(암 예 방, 심혈관계, 신경계 등)가 보고되면서 차에 대한 관심과 소비가 증가하고 있다(Weinreb 등, 2009; Yang 등, 2009).
차의 제조방법에 따라 녹차(non-oxidation tea), 반발효 차(semi-oxidation tea), 발효차(oxidation tea), 후발효차 또는 미생물발효차(dark tea or post-fermented tea)로 나 뉘며, 제조 방법에 따라 추출물 내의 성분, 색, 향기, 차의
맛 등으로 차이를 구분할 수 있다(Shon 등, 2004). 미생물발 효차는 미생물에 의한 찻잎의 생물전환(bioconversion) 또 는 발효를 통해 제조된다. 일반적으로 미생물발효차는 발효 하지 않은 녹차에 비해 맛이 부드러우며, 생물전환을 통하여 gallic acid, methoxy phenolic compound, poly-flavo- noids 등의 증가로 인해 항균작용, 혈중 콜레스테롤 감소 효과, 항산화 활성 등이 보고되었으며, 미생물발효차에 대한 관심이 증가하고 있다(Lv 등, 2013; Zhang 등, 2013). 대표 적인 미생물발효차는 보이차(Puerh tea, Yunnan, China), 아와반차(Awabancha, Tokushima, Japan), 청태전(Chung- taejeon, Jangheung, Korea) 등이 생산되고 있으며, 미생물 발효에 의한 미생물발효차는 동아시아 3개국에서만 계승되 고 있다(Lv 등, 2013). 미생물발효차는 미생물 종류와 발효 정도에 따라 맛과 향의 차이가 결정되기에 미생물의 역할이 매우 중요하다. 그러나 한국에서 미생물발효차에 대한 연구 는 매우 부족하며 중국의 미생물발효차인 보이차의 발효과 정에 대한 연구가 주를 이루고 있다(Zhang 등, 2016; Zhu 등, 2016). 최근 들어 한국의 미생물발효차에 대한 연구가 시작되고 있는 실정이다(Kim 등, 2017a; Kim 등, 2017b).
자연환경에서 미생물들은 환경에 맞게 다른 미생물들과 상호작용을 통해 미생물 군집(microbial community)을 형 성하며, 미생물 군집의 특성은 한 종류의 미생물이 가지고
있는 특성과는 다른 특성을 가지며, 미생물 군집 내의 상호 작용을 통해 다른 특성을 가질 수 있다(Kim 등, 2010). 따라 서 미생물 군집을 이해하는 것이 매우 중요하며, 미생물 간의 상호작용을 통해 고유의 특성을 가지는 미생물발효차에 있 어 숙성 미생물 군집 구조를 규명하는 연구는 미생물발효차 의 특성과 품질을 파악하는 데 매우 중요하다고 판단된다.
미생물 군집 구조 조사 방법 중 전통적인 배양방법은 전체 미생물 중에 1% 미만만이 배양 가능하기 때문에 미생물 다 양성과 군집 구조를 조사하는 데 한계가 있다. 이에 16S rRNA gene을 이용한 분자생태학적 방법들이 생태학적인 관계를 규명하기 위해 널리 이용되고 있다(Kim 등, 2010;
Kim 등, 2014).
미생물발효차는 동아시아에서 발전되었으며, 미생물의 발효과정을 통해 각각의 고유한 특성이 결정되기 때문에 불 발효차(녹차), 완전발효차(홍차) 등과 구별되는 특유의 맛, 색, 향을 지니게 된다. 본 연구에서는 동아시아 3개국의 전 통 미생물발효차인 청태전, 아와반차, 보이차의 발효미생물 의 공통점과 차이점을 규명하고자 하였다. 한국의 청태전, 일본의 아와반차와 중국의 보이차의 발효미생물의 규명은 미생물발효차 특성과 품질 향상을 위해 필요한 연구로 분자 생태학적 기법을 통해 분석했으며, 미생물의 생태학적 기초 지식은 후발효 과정에서 미생물 간의 상호작용을 이해하는 데 기여할 것으로 판단된다.
재료 및 방법
시료
본 연구에 사용된 미생물발효차는 2017년도 생산된 청태 전(Dayewon, Jangheong, Korea), 아와반차(Yamaneen, Tokushima, Japan), 보이차(Taetea Group, Kunming, China)를 각각 분석하였다.
DNA extraction
Total DNA는 미생물발효차 0.5 g을 사용했고, Fast DNA Spin kit for Soil(MP Bio, Santa Ana, CA, USA)을 이용하 여 추출하였다.
PCR for microbial diversity (DG-DGGE)
미생물 군집의 다양성 분석을 위해 시료로부터 얻은 DNA 를 주형으로 1차 PCR과 nested-PCR 반응을 수행하였다 (Muyzer 등, 1993; Muyzer, 1999). 1차 PCR은 27F(5′- AGA GTT TGA TCM TGG CTC AG-3′)와 1541R(5′- AAG GAG GTG ATC CAN CCR CA-3′)을 이용하여 50 μL 안에 1× PCR buffer, 20 mM MgCl2, 40 mM dNTP mixture, 각 primer(1 μM), template DNA와 0.5 U Taq polymerase를 첨가하여 PCR을 수행하였다. PCR 반응조건 은 우선 94°C에서 5분 동안 DNA를 변성시킨 후 94°C에서 45초 denaturation, 56°C에서 45초 annealing, 72°C에서
45초 extension을 25 cycles 수행한 다음 72°C에서 7분 동안 final extension을 수행하였다. PCR products는 ethi- dium bromide 염색 후 1.2% agarose gel에서 확인하였다.
Double gradient-denaturing gradient gel electropho- resis(DG-DGGE) 분석을 위하여 1차 PCR의 증폭산물을 주형으로 2차 PCR을 수행하였으며, 이때 16S rDNA를 증 폭하기 위하여 사용된 primer는 40개의 GC-clamp가 붙은 341F-GC(5′-CGC CCG CCG CGC GCG GCG GGC GGG GCG GGG GCA CGG GGG GCC TAC GGG AGG CAG CAG-3′)와 786R(5′-CTA CCA GGG TAT CTA ATC-3′) 을 이용하였다(Forney 등, 2004; Kim 등, 2017a). 2차 PCR 을 위한 PCR mixture는 1차 PCR에 사용된 것과 동일하며, 사용된 PCR 반응조건은 94°C에서 5분 동안 DNA를 변성시 켜 94°C에서 45초 denaturation, 60°C에서 45초 anneal- ing, 72°C에서 45초 extension 하고 72°C에서 7분 동안 fi- nal extension을 수행하였다. Annealing 온도는 초기에는 60°C에서 시작해서 1 cycle당 0.5°C 감소하게 20 cycles 수행하였고, 그 후 50°C에서 15 cycles을 수행하여 touch- down PCR을 완료하였다. 얻어진 2차 PCR의 산물을 이용 하여 DG-DGGE를 수행하였다(Muyzer 등, 1993).
DG-DGGE 시료 운전조건
PCR 산물은 DcodeTM System(Bio-Rad, Hercules, CA, USA)을 이용하여 DG-DGGE를 수행하였다(Muyzer 등, 1993; Lee 등, 2005). DG-DGGE gel은 10~12% poly- acrylamide(37.5:1=acrylamide : bisacrylamide)에 urea 와 formamide 변성제를 40~70%의 농도구배가 형성되도 록 제작했으며, 제작된 DG-DGGE gel에 PCR 증폭산물을 30 µL씩 loading 하여 1× TAE buffer(40 mM Tris, 20 mM acetic acid, 1 mM EDTA, pH 8.0)에서 60°C, 120 V로 18시간 동안 전기영동하였다. 전기영동이 끝난 DG-DGGE gel은 ethidium bromide(1:10,000)에서 염색한 후 UV로 확인하였다(Kim 등, 2014).
16S rRNA gene clone library를 통한 박테리아 군집분석 16S rRNA gene clone library 분석을 위한 PCR은 27F (5′-AGA GTT TGA TCM TGG CTC AG-3′)와 1541R(5′
-AAG GAG GTG ATC CAN CCR CA-3′)을 사용하였다 (Forney 등, 2004). 각각의 primer를 이용하여 50 μL 안에 1× PCR buffer, 20 mM MgCl2, 40 mM dNTP mixture, 각 primer(1 μM), template DNA와 0.5 U Taq polymer- ase를 첨가하여 PCR을 수행하였다. 반응조건은 94°C에서 5분 동안 DNA를 pre-denaturation 시켜 94°C에서 1분 denaturation, 60°C에서 1분 annealing, 72°C에서 1분 30 초 extension 하고 72°C에서 5분 동안 final extension을 수행하였다. 16S rRNA gene clone library 구축을 위해 PCR 증폭산물을 1.2% agarose gel에 loading 한 후 약 1,500 bp 크기의 band를 잘라 HiGeneTM Gel & PCR Puri-
fication Solution Kit(Biofact, Daejeon, Korea)을 이용하 여 정제한 다음, All-in-one PCR cloning kit(Biofact)을 이용하여 ligation 하고, E. coli DH5α(RBC Bioscience, New Taipei, Taiwan)에 형질전환 후 LB+kanamycin 배지 에서 키워 생성된 colony들을 16S rRNA gene sequence 분석에 이용하였다.
염기서열 분석
DG-DGGE gel 상에서 다른 위치에 존재하는 각각의 band를 분리한 후 멸균증류수 50 μL를 첨가하고 4°C에서 하룻밤 동안 방치하여 상등액을 취하였다. 회수한 DNA를 주형으로 nested PCR에 사용한 primer를 이용하여 재증폭 하였으며, PCR 산물은 DNA recovery kit(Biofact)으로 정 제한 후 cloning을 수행하였다. Cloning은 All-in-one vec- tor(Biofact)를 이용하여 manufacturer’s protocol을 따라 수행한 후 16S rRNA gene sequence를 분석하였다. 분석 된 16S rRNA gene sequence는 NCBI(www.ncbi.nlm.nih.
gov)의 GenBank database를 이용하여 BLAST search를 통해 분석하였다.
미생물발효차 미생물 군집 비교를 위한 정량 real-time PCR
각 시료별 미생물의 군집의 크기를 정량적으로 평가하기 위해 qPCR을 수행하였다. 미생물 군집을 정량하기 위해서 27F와 518R(5′-ATT ACC GCG GCT GCT GG-3′) pri- mer를 이용하였고(Murray 등, 1996), Escherichia coli DH5α의 total DNA를 주형으로 PCR을 수행하여 491 bp의 PCR 산물을 얻었다. 곰팡이 군집의 정량을 위해 SR-4F(5′- AG CCG CGG TAA TTC CAG CT-3′)와 SR-7R(5′-TCC TTG GGC AAA TGC TTT CGC-3′)을 이용하였고, As- pergillus total DNA를 주형으로 하여 388 bp의 PCR 산물 을 얻었다(Nakayama 등, 1996). 또한 고세균 군집의 정량 을 위해서는 arch-21F(5′-TCC GGT TGA TCC YGC CGG-3′)와 arch-516R(5′-GGT DTT ACC GCG GCK GCT G-3′)을 이용하였고, 청태전 total DNA를 이용하여 496 bp의 PCR 산물을 확보하였다(Gantner 등, 2011). 각 각의 PCR 산물은 All-in-one vector(Biofact)에 cloning 하였으며, plasmid DNA는 HiGeneTM Plasmid Mini Prep Kit(Biofact)을 이용하여 추출하였다. 염기서열 분석은 M13- 20F와 M13-20R(All-in-one Vector Systems manual)을 이용하여 분석하였고, BLAST search를 통해 확인하였다.
염기서열이 확인된 plasmid DNA는 real-time PCR을 이용 하여 melting curve(Bio-Rad) 분석 후 정량분석을 위한 표 준 유전자(artificial standard clone)로 사용하였다.
정량 PCR을 위해 CFX96 TouchTM Real-Time PCR Detection System(Bio-Rad)과 iTaqTM SYBR® Green Supermix with ROX(Bio-Rad)를 이용하였다. 미생물 16S rRNA 유전자 정량 PCR은 각각의 primer를 이용하였으며
반응조건은 95°C에서 15분 동안 pre-denaturation 시켜 95°C에서 30초 denaturation, 67°C에서 30초 annealing, 72°C에서 30초 extension 후 fluorescence를 측정하고 45 cycles을 수행하였다. 그리고 final extension은 72°C에서 5분 동안 수행하였다. Melting curve 분석은 65°C부터 95°C까지 0.2°C씩 증가시키면서 fluorescence를 측정하였 다. 곰팡이 군집과 고세균 군집의 정량을 위해서 앞서 사용 된 PCR program을 이용했으며, annealing 온도는 69°C와 68°C에서 각각 수행하였다.
정량을 위해서 표준 유전자를 serial dilutions 하여 real- time PCR을 수행하였으며, DNA 농도는 NanoDrop ND- 2000(Thermo Scientific, Wilmington, DE, USA)을 이용 하여 1 ng/µL를 분석에 이용하였다(Kim 등, 2014; Kim 등, 2017a)
결과 및 고찰
DG-DGGE를 통한 미생물 군집분석
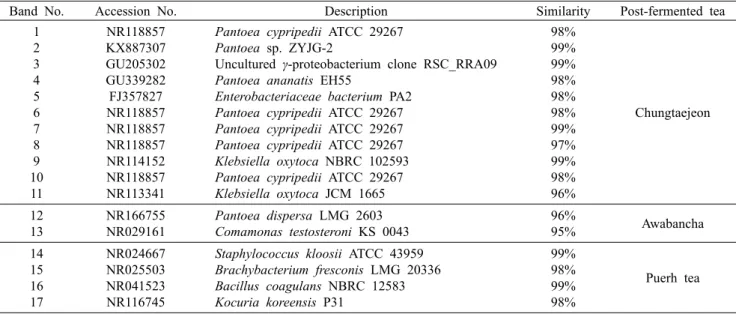
동아시아 3개국의 미생물발효차를 시료로 하여 미생물 군집 구조 분석 기법인 DG-DGGE를 이용하여 각 시료 내의 박테리아 군집 패턴을 확인하였다(Muyzer 등, 1993)(Fig.
1). 또한 박테리아 군집 구조 분석을 위해 청태전, 아와반차, 보이차의 DG-DGGE gel로부터 각각 11개, 2개, 4개의 밴 드를 확인하고 이를 회수하여 염기서열을 결정하였으며, NCBI(www.ncbi.nlm.nih.gov)의 GenBank database를 이 용하여 BLAST search(blastn)를 통해 상동성을 비교 검색 하였다(Table 1). DG-DGGE를 통한 청태전 우점 박테리아 는 총 11개의 밴드를 분리하여 분석한 결과, genus 수준에 서 4개의 종으로 Pantoea sp.(band 1, 2, 4, 6, 7, 8, 10), Enterobacteriaceae sp.(band 5), Klebsiella sp.(band 9, 11), Uncultured γ-proteobacterium(band 3)으로 확인되 었다(Table 1). 아와반차의 우점 박테리아는 2개 종으로 Pantoea sp.(band 12)와 Comamonas sp.(band 13)로 분 석되었다. 보이차의 우점 박테리아는 4개의 종으로 Staph- ylococcus sp.(band 14), Brachybacterium sp.(band 15), Bacillus sp.(band 16)와 Kocuria sp.(band 17)로 분석되 었다. 동아시아의 미생물발효차의 DG-DGGE를 통한 박테 리아 군집 구조분석을 수행한 결과 밴드 패턴이 전체적으로 상이한 것을 확인할 수 있었다. 또한 각국의 미생물발효차 간에 Pantoea sp. 외에 공통적인 미생물은 관찰되지 않았으 며, 다양한 미생물들이 분석되어 미생물발효차의 제다방법 에 따라 우점 미생물이 달라지는 것을 확인할 수 있었다.
16S rRNA gene clone library를 통한 미생물 군집 다양성 분석
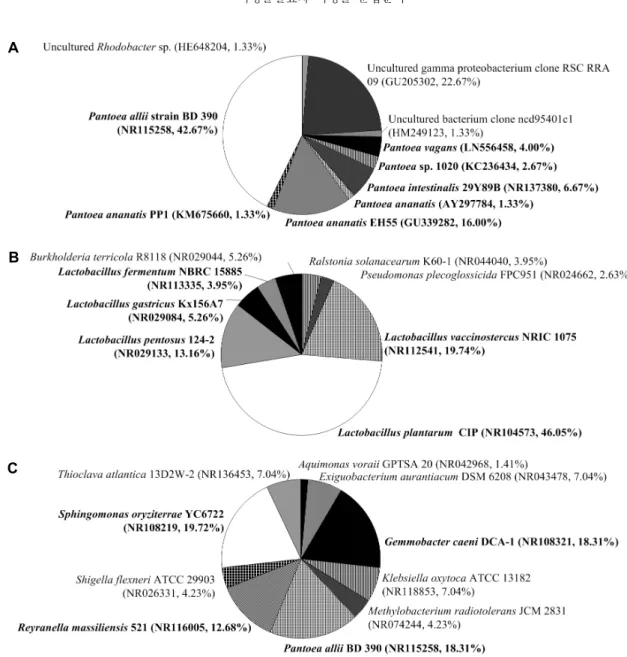
청태전의 16S rRNA gene의 박테리아 군집분석 결과 Pantoea sp.(74.67%)와 uncultured γ-proteobacterium (22.67%)이 우점하였다. 아와반차 박테리아 군집은 Lacto-
A B C U/F Acry/bis-acry conc. conc.
Fig. 1. DG-DGGE patterns of 16S rRNA gene fragments of East- Asia post-fermented teas bacteria. The DG-DGGE was electro- phoresed on a 8∼12% polyacrylamide gel with a 40∼70% urea- formamide gradient. The bright bands on the DGGE were ex- cised and the amplified DNA fragments partially sequenced and compared to known sequences using BLAST. A, Chungtaejeon (Korea); B, Awabancha (Japan); C, Pureh tea (China). U/F conc., urea/formamide concentration; Acry/bis-acry conc., acrylamide/
bis-acrylamide concentration.
Table 1. Identity of the bands obtained from 16S rRNA gene DG-DGGE bands and those of the closest relatives in the GenBank database, indicating the detected bacteria among East-Asia post-fermented teas
Band No. Accession No. Description Similarity Post-fermented tea 1
2 3 4 5 6 7 8 9 10 11
NR118857 KX887307 GU205302 GU339282 FJ357827 NR118857 NR118857 NR118857 NR114152 NR118857 NR113341
Pantoea cypripedii ATCC 29267 Pantoea sp. ZYJG-2
Uncultured γ-proteobacterium clone RSC_RRA09 Pantoea ananatis EH55
Enterobacteriaceae bacterium PA2 Pantoea cypripedii ATCC 29267 Pantoea cypripedii ATCC 29267 Pantoea cypripedii ATCC 29267 Klebsiella oxytoca NBRC 102593 Pantoea cypripedii ATCC 29267 Klebsiella oxytoca JCM 1665
98%
99%
99%
98%
98%
98%
99%
97%
99%
98%
96%
Chungtaejeon
12 13
NR166755 NR029161
Pantoea dispersa LMG 2603 Comamonas testosteroni KS 0043
96%
95% Awabancha 14
15 16 17
NR024667 NR025503 NR041523 NR116745
Staphylococcus kloosii ATCC 43959 Brachybacterium fresconis LMG 20336 Bacillus coagulans NBRC 12583 Kocuria koreensis P31
99%
98%
99%
98%
Puerh tea bacillus sp.(88.16%)가 우점하였고, Burkholderia sp.(5.26
%)가 다음으로 자리 잡고 있었다. 보이차는 Sphingomonas sp.(19.72%), Pantoea sp.(18.31%)와 Gemmobacter sp.
(18.31%), Reyranella sp.(12.68%)가 우점하는 것을 확인 하였다(Fig. 2, Table 2). 청태전에 우점하는 박테리아는 γ- proteobacteria(Pantoea sp., uncultured γ-proteobac- terium)가 97.34%로 우점하고 α-proteobacteria(Rhodo- bacter sp.)가 1.33%로 분석되었다. 아와반차에는 firmi- cutes(Lactobacillus sp.)가 88.16%, β-proteobacteria (Ralstonia sp., Burkholderia sp.)가 9.21% 우점하고 있으 며, 보이차에는 α-proteobacteria(Gemmobacter sp., Methylobacterium sp., Reyranella sp., Sphingomonas sp.)가 54.94%, γ-proteobacteria(Aquimonas sp., Kleb- siella sp., Pantoea sp., Shigella sp.)가 30.99%로 우점하 는 것으로 분석되었다. 청태전은 γ-proteobacteria, 아와반 차는 firmucutes, 보이차는 α-proteobacteria와 γ-pro- teobacteria가 우점하는 것으로 분석되어 동아시아 3개국 의 미생물발효차의 발효미생물의 차이를 뚜렷하게 구분되 는 것을 알 수 있다. Pantoea sp.는 주로 eucalyptus와 같은 식물의 잎에서 분리되며, Pantoea agglomerans는 저분자 항생제인 pantocin A와 pantocin B를 생성한다고 알려져 있다. 또한 meta-genome 분석을 통해 떡차에서 우점하며, 발효과정에서도 Sphingomonas sp., Bacillus sp.와 함께 우점하고 있다고 보고되었다(Jin 등, 2003; Wright 등, 2006; Baik 등, 2012; Shim 등, 2013). Pantoea agglom- erans T71의 gallic acid decarboxylase는 polyphenol tannic acid와 gallic acid의 분해과정에 관여하며, gallic acid를 tannic acid로 전환시키며, Pantoea dispersa Y08 은 carotenoids를 aroma compound로 전환한다고 보고되 었다(Zeida 등, 1998; Zhao 등, 2015). Sphingomonas sp.
A
B
C
Fig. 2. Dominant bacterial OTUs detected in the post-fermented teas 16S rRNA gene clone libraries. A, Chungtaejeon (Korea);
B, Awabancha (Japan); C, Puerh tea (China). Bold, dominant community.
는 aromatic compounds를 분해하며, 다양한 전통발효식품 의 제조 시 발효에 관여하는 중요한 박테리아로 알려져 있다 (Peng 등, 2015; Yi 등, 2017). 또한 Lactobacillus sp.는 발효를 통해 젖산 및 여러 가지 대사산물을 생성하는 미생물 로 각종 발효식품, 의약품, 사료 첨가제 등에 광범위하게 이 용되며, 최근 건강 증진 및 질병 예방의 특징을 가지는 pro- biotics 균주로 연구와 활용이 증가하고 있다(Saez-Lara 등, 2015). 또한 Lactobacillus fermentum OCS19로 녹차 추출물을 발효하여 급성 알코올성 지방간에 효과를 확인하 였다(Park 등, 2012). Pantoea sp., Sphingomonas sp., Lactobacillus sp.는 다양한 물질들의 생물전환능이 보고되 었으며, 동아시아 3개국의 전통 미생물발효차에서 각각 우 점하고 있는 것에 기반하여 미생물발효차의 발효과정에서 중요한 역할을 하는 것으로 판단된다. 미생물 군집분석을 통한 미생물발효차의 특징이 발현되며, 우점미생물의 생물
전환 대사과정 및 효능에 관한 활발한 연구를 통해 많은 지 식이 축적되기를 기대한다.
미생물발효차 우점 미생물의 군집 크기 비교
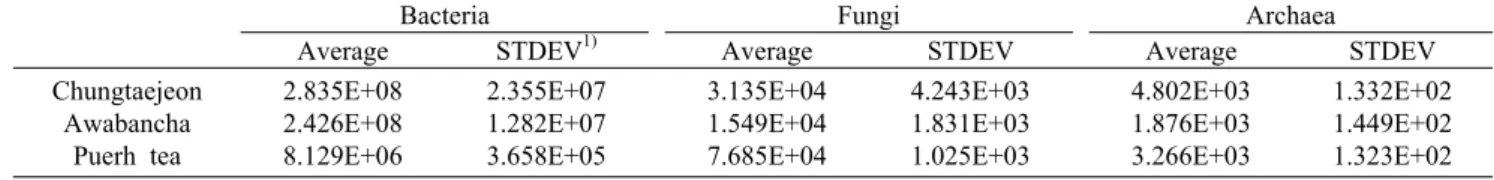
동아시아 3개국의 전통 미생물발효차의 군집분석을 수행 하여 우점 박테리아를 분석하고 박테리아, 고세균 및 곰팡이 의 군집 크기를 분석하였다. 총균수는 real-time PCR을 이 용하여 측정하고 군집의 크기를 비교하였다(Fig. 3, Table 3). 박테리아의 총균수는 청태전 2.835×108±2.355×107 molecule・copy/µL, 아와반차 2.426×108±1.282×107 mol- ecule・copy/µL, 보이차는 8.129×106±3.658×105 mole- cule・copy/µL로 분석되었다. 이 결과로부터 청태전과 아와 반차는 박테리아 군집의 크기는 2.42×108 molecule・copy/
µL 이상이나, 보이차는 8.12×106 molecule・copy/µL 정도 로 청태전과 아와반차 박테리아 군집이 보이차 박테리아 군
A
B
C
Fig. 3. Comparison of microbial community size using the real- time PCR from the East-Asia post-fermented teas. A, bacterial 16S rRNA gene; B, fungal 18S rRNA gene; C, archaeal 16S rRNA.
Table 3. Quantification using the bacterial and archaeal 16S rRNA gene and fungal 18S rRNA gene
Bacteria Fungi Archaea
Average STDEV1) Average STDEV Average STDEV
Chungtaejeon Awabancha
Puerh tea
2.835E+08 2.426E+08 8.129E+06
2.355E+07 1.282E+07 3.658E+05
3.135E+04 1.549E+04 7.685E+04
4.243E+03 1.831E+03 1.025E+03
4.802E+03 1.876E+03 3.266E+03
1.332E+02 1.449E+02 1.323E+02
1)STDEV: standard deviation.
집에 비해 약 34.8배와 29.8배 이상 큰 것으로 분석되었다.
곰팡이의 총균수는 청태전 3.135×104±4.243×103 mole- cule・copy/µL, 아와반차 1.549×104±1.831×103 mole- cule・copy/µL, 보이차 7.685×104±1.025×103 molecule・
copy/µL로 분석되었다. 보이차의 군집이 청태전과 비교해 2.45배, 아와반차와 비교해 4.96배 큰 것을 확인할 수 있었 다. 또한 고세균의 총균수는 청태전 4.802×103±1.332×
102 molecule・copy/µL, 아와반차 1.876×103±1.449×102 molecule・copy/µL, 보이차 3.266×103±1.323×102 mol- ecule・copy/µL로 분석되었다. 청태전 총균수는 아와반차 에 비해 2.56배, 보이차에 비해 1.47배 정도 큰 것을 확인할 수 있었다. 청태전의 고세균 총균수가 가장 크기는 하지만, 박테리아 총균수와 비교해 적은 차이가 나는 것을 확인할 수 있었다. 중국 전통 미생물발효차인 보이차는 분자생물학 적 기법을 통해 발효미생물을 분석한 결과 Aspergillus spp.
와 Penicillium spp.가 우점한다고 보고되었으며, 박테리아 보다 곰팡이류에 의한 발효 연구가 주를 이루고 있다(Abe 등, 2008; Zhao 등, 2010; Tian 등, 2013).
본 연구는 동아시아 3개국의 전통 미생물발효차의 미생 물 군집의 크기를 분석하고 미생물 다양성을 확인하였다.
미생물발효차의 미생물(박테리아, 곰팡이, 고세균)의 군집 의 크기는 박테리아 군집은 한국 청태전과 일본의 아와반차 가 중국의 보이차보다 크며, 곰팡이 군집의 크기는 중국의 보이차가 한국의 청태전과 일본의 아와반차보다 큰 것으로 확인되었다. 이를 통해 한국의 청태전과 일본의 아와반차는 박테리아에 의한 발효과정이 이루어질 가능성이 더 크며, 중국의 보이차는 곰팡이에 의한 발효과정이 이루어질 수 있 다고 판단할 수 있다. 또한 미생물 발효에 의해 숙성되는 청태전과 아와반차의 발효미생물은 청태전 Pantoea sp., 아 와반차 Lactobacillus sp.로 상이한 것을 확인할 수 있었다.
우점 미생물은 주어진 환경적 요소들을 가장 잘 이용하여 활발한 신진대사를 통해 우점한다고 알려져 있다. 일반적으 로 oligotrophic(청정하천지역) 환경에서는 미생물 다양성 은 높고 우점도는 낮지만, eutrophic(오염하천지역)에서는 미생물 다양성이 낮고 단일 미생물의 높은 우점도를 보인다 고 보고되었다(Kim 등, 2008). 이처럼 동아시아 3개국의 미생물발효차에 따른 각각의 미생물 우점도가 높은 것은 우 점된 미생물이 각기 다른 제다방법에 따른 찻잎을 가장 잘 이용하는 미생물이 판단할 수 있다. 또한 생물전환은 미생물 및 효소를 이용한 미생물 발효과정을 뜻하며, 전구물질에서 유용 대사산물을 생산 및 제조하는 기술이다(Kim 등, 2019).
생물전환율은 동일한 species 내에 genus가 다른 미생물을 이용할 경우 대사산물 및 생물전환능이 다르게 나타날 수 있어, 찻잎의 발효공정에 이용되는 미생물의 차이에 따라 다른 품질의 차가 제조될 수 있을 것으로 판단된다(Lee 등, 2018). 따라서 미생물발효차에서 발효균주는 차의 품질을 결정짓는 매우 중요한 요소로 판단된다. 본 연구를 통해 동 아시아 3개국의 미생물발효차 청태전, 아와반차, 보이차의
발효과정에 관여하는 미생물이 서로 다르며, 미생물의 차이 에 따라 맛과 향의 차이를 나타낼 것으로 사료되며, 나라별 로 제다방법의 차이로 인한 발효 미생물의 차이를 확인하였 다. 따라서 본 연구 결과를 활용하여 미생물발효차의 이해와 나라별 미생물발효차 제다법을 이해하는 데 기여할 수 있을 것으로 기대된다.
요 약
미생물발효차는 차나무(Camellia sinensis (L.) O Kuntze) 의 잎을 미생물 발효과정을 거쳐 생산된다. 본 연구에서 한 국의 청태전, 일본의 아와반차, 중국 보이차에 우점하는 박 테리아 분석을 위해 double gradient-denaturing gradient gel electrophoresis(DG-DGGE)와 16S rRNA gene clone library 기법으로 분석하였다. 각국의 미생물발효차에 우점 하는 박테리아가 청태전은 Pantoea sp., 아와반차는 Lac- tobacillus sp., 보이차는 Sphingomonas sp.인 것을 확인했 다. 미생물 군집 크기를 분석한 결과 청태전과 아와반차는 박테리아가 우점하고, 중국 미생물발효차는 곰팡이가 우점 하는 것으로 분석되었다.
REFERENCES
Abe M, Takaoka N, Idemoto Y, Takagi C, Imai T, Nakasaki K.
Characteristic fungi observed in the fermentation process for Puer tea. Int J Food Microbiol. 2008. 124:199-203.
Baik KS, Seong CN, Hwang YM, Kim GA, Lee NR, Kim D, et al. Microbial diversity of Ddek cha using DNA sequence analysis. J Korean Tea Soc. 2012. 18(3):86-91.
Forney LJ, Zhou X, Brown CJ. Molecular microbial ecology:
land of the one-eyed king. Curr Opin Microbiol. 2004. 7:210- 220.
Gantner S, Andersson AF, Alonso-Sáez L, Bertilsson S. Novel primers for 16S rRNA-based archaeal community analyses in environmental samples. J Microbiol Methods. 2011. 84:12-18.
Ho CT, Lin JK, Shahidi F. Tea and tea products: chemistry and health-promoting properties. CRC Press, Boca Raton, FL, USA. 2008. p 131-160.
Jin M, Liu L, Wright SAI, Beer SV, Clardy J. Structural and functional analysis of pantocin A: an antibiotic from Pantoea agglomerans discovered by heterologous expression of cloned genes. Angew Chem Int Ed Engl. 2003. 42:2898-2901.
Kim B, Lee SE, Song MY, Choi JH, Ahn SM, Lee KS, et al.
Implementation of artificial neural networks (ANNs) to analy- sis of inter-taxa communities of benthic microorganisms and macroinvertebrates in a polluted stream. Sci Total Environ.
2008. 390:262-274.
Kim BH, Baek KH, Cho DH, Sung Y, Koh SC, Ahn CY, et al. Complete reductive dechlorination of tetrachloroethene to ethene by anaerobic microbial enrichment culture developed from sediment. Biotechnol Lett. 2010. 32:1829-1835.
Kim BH, Jang JO, Joa JH, Kim JA, Song SY, Lim CK, et al.
A comparison of the microbial diversity in Korean and Chi- nese post-fermented teas. Microbiol Biotechnol Lett. 2017a.
45:71-80.
Kim BH, Jang JO, Kang Z, Joa JH, Moon DG. The microbial diversity analysis of the Korea traditional post-fermented tea
(Chungtaejeon). Kor J Microbiol. 2017b. 53:170-179.
Kim BH, Jang JO, Lee JH, Park YE, Kim JG, Yoon YC, et al.
Evaluation of the anti-oxidant activity of Pueraria extract fer- mented by Lactobacillus rhamnosus BHN-LAB 76. J Life Sci.
2019. 29:545-554.
Kim BH, Ramanan R, Cho DH, Oh HM, Kim HS. Role of Rhi- zobium, a plant growth promoting bacterium, in enhancing al- gal biomass through mutualistic interaction. Biomass Bio- energy. 2014. 69:95-105.
Lee JH, Yoon YC, Kim JK, Park YE, Hwang HS, Kwon GS, et al. Antioxidant and whitening effects of the fermentation of barley seeds (Hordeum vulgare L.) using lactic acid bacteria.
J Life Sci. 2018. 28:444-453.
Lee JW, Kim BH, Ahn CY, Kim HS, Yoon BD, Oh HM. Anal- ysis of microbial community during the anaerobic dechlorina- tion of perchloroethylene and trichloroethylene. Kor J Micro- biol. 2005. 41:281-286.
Lv HP, Zhang YJ, Lin Z, Liang YR. Processing and chemical constituents of Pu-erh tea: A review. Food Res Int. 2013. 53:
608-618.
Murray AE, Hollibaugh JT, Orrego C. Phylogenetic compositions of bacterioplankton from two California estuaries compared by denaturing gradient gel electrophoresis of 16S rDNA frag- ments. Appl Environ Microbiol. 1996. 62:2676-2680.
Muyzer G, de Waal EC, Uitterlinden AG. Profiling of complex microbial populations by denaturing gradient gel electropho- resis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl Environ Microbiol. 1993. 59:695- 700.
Muyzer G. DGGE/TGGE a method for identifying genes from natural ecosystems. Curr Opin Microbiol. 1999. 2:317-322.
Nakayama T, Watanabe S, Mitsui K, Uchida H, Inouye I. The phylogenetic relationship between the Chlamydomonadales and Chlorococcales inferred from 18SrDNA sequence data.
Phycol Res. 1996. 44:47-55.
Park JH, Kim Y, Kim SH. Green tea extract (Camellia sinensis) fermented by Lactobacillus fermentum attenuates alcohol-in- duced liver damage. Biosci Biotechnol Biochem. 2012. 76:
2294-2300.
Peng Q, Yang Y, Guo Y, Han Y. Analysis of bacterial diversity during acetic acid fermentation of Tianjin Duliu aged vinegar by 454 pyrosequencing. Curr Microbiol. 2015. 71:195-203.
Saez-Lara MJ, Gomez-Llorente C, Plaza-Diaz J, Gil A. The role of probiotic lactic acid bacteria and bifidobacteria in the pre- vention and treatment of inflammatory bowel disease and oth- er related diseases: A systematic review of randomized human clinical trials. Biomed Res Int. 2015. Article ID: 505878. doi:
10.1155/2015/505878.
Shim HJ, Cho JY, Moon JH, Kim SJ, Kim D, Shibn KH, et al.
Changes of bacterial communities in microbial-fermented tea during fermentation. J Korean Tea Soc. 2013. 19(3):91-98.
Shon MY, Kim SH, Nam SH, Park SK, Sung NJ. Antioxidant activity of Korean green and fermented tea extracts. J Life Sci. 2004. 14:920-924.
Tian J, Zhu Z, Wu B, Wang L, Liu X. Bacterial and fungal com- munities in Pu′er tea samples of different ages. J Food Sci.
2013. 78:M1249-M1256.
Weinreb O, Amit T, Mandel S, Youdim MBH. Neuroprotective molecular mechanisms of (-)-epigallocatechin-3-gallate: a re- flective outcome of its antioxidant, iron chelating and neurito- genic properties. Genes Nutr. 2009. 4:283-296.
Wright SAI, Jin M, Clardy J, Beer SV. The biosynthetic genes of pantocin A and pantocin B of Pantoea agglomerans Eh318.
Acta Hortic. 2006. 704:313-320.
Yang CS, Wang X, Lu G, Picinich SC. Cancer prevention by tea: animal studies, molecular mechanisms and human rele- vance. Nat Rev Cancer. 2009. 9:429-439.
Yi L, Su G, Hu G, Peng Q. Diversity study of microbial com- munity in bacon using metagenomic analysis. J Food Saf.
2017. 37:e12334. https://doi.org/10.1111/jfs.12334
Zeida M, Wieser M, Yoshida T, Sugio T, Nagasawa T. Purifica- tion and characterization of gallic acid decarboxylase from Pantoea agglomerans T71. Appl Environ Microbiol. 1998.
64:4743-4747.
Zhang L, Zhang ZZ, Zhou YB, Ling TJ, Wan XC. Chinese dark teas: Postfermentation, chemistry and biological activities.
Food Res Int. 2013. 53:600-607.
Zhang W, Yang R, Fang W, Yan L, Lu J, Sheng J, et al. Charac- terization of thermophilic fungal community associated with pile fermentation of Pu-erh tea. Int J Food Microbiol. 2016.
227:29-33.
Zhao Y, Zhong GF, Yang XP, Hu XM, Mao DB, Ma YP. Biocon- version of lutein to form aroma compounds by Pantoea dis- persa. Biotechnol Lett. 2015. 37:1687-1692.
Zhao ZJ, Tong HR, Zhou L, Wang EX, Liu QJ. Fungal colo- nization of Pu-erh tea in Yunnan. J Food Saf. 2010. 30:769- 784.
Zhu Y, Luo Y, Wang P, Zhao M, Li L, Hu X, et al. Simultaneous determination of free amino acids in Pu-erh tea and their changes during fermentation. Food Chem. 2016. 194:643-649.