비천연 아미노산이 도입된 재조합 요산분해효소의 생산을 위한 배양 최적화
조정행1,2, 박성주2, 최종일2*
Optimization of Cultivation Conditions for Production
of Recombinant Urate Oxidase with Unnatural Amino Acids
Jeong-Haeng Cho1,2, Seongju Park2, and Jong-il Choi2*
Received: 3 March 2020 / Revised: 23 March 2020 / Accepted: 24 March 2020
© 2020 The Korean Society for Biotechnology and Bioengineering
Abstract: In this study, the medium composition, carbon source, non-natural amino acid concentration, and induction condition were optimized for the production of recombinant urate oxidase with non-natural amino acids. Recombinant Escherichia coli strain was first cultured in various media including LB, 2 × YT and B, and cell growth and urate oxi- dase expression were compared. Based on the selected B medium, the concentrations of each component in medium were optimized through response surface methodology. At the optimal medium compositions, urate oxidase with non-natural amino acids was produced with 64.92 U/mg specific protein activity in flask culture. Among various carbon sources, the highest urate oxidase activity was obtained at the glucose con- centration of 5 g/L. The optimal inducer concentrations were 1 mM of IPTG. 1 mM of non-natural amino acids was efficient for urate oxidase production. This results will be useful for the production of therapeutic proteins with prolonged activity.
Keywords: recombinant urate oxidase, non-natural amino acids, recombinant Escherichia coli, response surface method, optimization
1. INTRODUCTION
미생물을 이용한 치료용 단백질의 생산에 관한 많은 기초 연 구들이 진행되었고, 이를 바탕으로 발효를 통한 치료용 단백 질의 대량생산이 이루어지고 있다. 최근에는 야생형 균주로 부터의 치료용 단백질의 생산뿐 만 아니라, 보다 효율적으로 생산하기 위해 대장균 및 효모에 클로닝 및 재조합 기술을 도입하여 재조합 치료용 단백질을 생산하고자 하는 많은 연 구들이 진행되고 있다 [1].
다양한 질병의 치료를 위한 치료용 단백질이 실제 사용되 기 위하여 여러 가지 고려사항들이 있지만, 그 중 하나는 반 복적인 복용 및 주입을 피하기 위하여 약물의 생체내 지속시 간을 연장해주어야 한다는 것이다. 이러한 생체내 치료용 단 백질의 지속성을 증가시키는 한 가지 방법은 인간 혈청 알부 민을 이용하는 것이다 [2]. 치료용 단백질이 환자에게 주입 될 때 치료용 단백질의 분해가 이루어지지만, 인간 혈청 알 부민을 치료용 단백질에 결합시킬 경우 반감기가 증대되었 다는 보고가 있다 [3]. 실제 많은 치료용 단백질의 생산에 인 간 혈청 단백질의 결합을 이용하고 있다 [4-6]. 링커를 사용 하여 화학적 결합에 의해 인간 혈청 알부민을 치료용 단백질 에 연결함으로써 치료용 단백질의 반감기를 증가시킬 수 있 는 기술을 알부미네이션이라고 하며, 이러한 알부미네이션 을 통하여 치료용 단백질의 반감기를 증가시키고 보다 더 높 은 약리학적 효과를 유도할 수 있다고 보고 되어있다 [7-11].
대표적인 대사질환의 하나인 통풍은 퓨린 대사의 최종 산 물인 요산의 혈액 내 축적으로 인하여 시작된다. 초기에는 고요산혈증이라는 증상이 나타나며 이후에 요산염을 생성
1주식회사 프로앱텍
1ProAbtech Co., Ltd., Gwangju 61005, Korea
2전남대학교 생물공학과
2Department of Biotechnology and Engineering, Interdisciplinary Program of Bioenergy and Biomaterials, Chonnam National University, Gwangju 61186, Korea
Tel: +82-62-530-1846, Fax: +82-62-530-1849 e-mail: [email protected]
Research Paper
하여 연골 및 관절 주위 조직과 피하조직에 침착되고, 이로 인해 염증과 통증을 동반하는 통풍으로 발전하게 된다. 혈액 내 축적된 요산을 분해하기 위하여 요산분해효소와 같은 산 화효소를 치료제로 사용하게 된다. 선천적 및 후천적으로 요 산분해효소 유전자의 변이가 생기거나 잘못된 식습관으로 인한 체내 요산이 축적된 환자의 경우 식이요법을 통해 체내 퓨린의 양을 줄이거나 직접적인 요산분해효소의 주입을 통 하여 치료하는 방법들이 사용되고 있다 [12-14].
최근 아스퍼질러스 플라버스(Aspergillus flavus)에서 유래 한 요산분해효소의 반감기를 늘리기 위하여 인간 혈청 알부 민과 결합할 수 있는 재조합 요산분해효소의 생산 연구가 보 고되었다 [15]. 재조합 요산분해효소는 인간 혈청 단백질의 결합을 위하여 160, 174번 아미노산이 엠버코돈(amber codon) 으로 치환되어 있고, 치환된 엠버코돈에 비천연 아미노산 4- azido-phenylalanine을 지정하여 DBCO-PEG4-MAL 링커를 통한 인간 혈청 알부민이 결합될 수 있는 비천연 아미노산을 함유한 단백질이다 [15].
이러한 비천연 아미노산이 첨가된 치료용 재조합 단백질 의 효과적인 생산을 위해서는 배양 배지, 단백질 발현, 그리 고 비천연 아미노산의 농도 등의 최적화가 필요하다. 이에 본 연구에서는 대장균을 이용하여 비천연 아미노산 4-azido- phenylalanine 가 도입된 재조합 요산분해효소의 생산을 위한 배양 최적화를 위하여, 반응표면분석법을 이용하여 배지 최 적화를 진행하였으며, 발현 조건 및 비천연 아미노산의 농도 등을 최적화하여 향 후 재조합 요산분해효소를 대량 생산할 수 있는 배양 조건을 확립하고자 하였다.
2. MATERIALS AND METHODS
2.1. 균주
재조합 요산분해효소를 생산하기 위한 모태 균주로 UAG 종 결 기능이 제거된 Escherichia coli C321.A(Addgene, Cambridge, MA, USA)를 숙주 대장균으로 사용하였다 [15]. Aspergillus flavus 유래 요산분해효소의 160번과 174번 아미노산의 염기 서열이 UAG로 치환된 재조합 요산분해효소 유전자를 포함 한 플라스미드 pQE80_UoxR과 mutant tRNA synthetase와 tRNA 유전자가 포함된 플라스미드 pEVOL_pAzF를 E. coli C321.DA에 형질전환하여 만든 E. coli C321.DA (pEVOL_pAzF pQE80_UoxR) 균주를 생산균주로 사용하였다 [15]. 재조합 요산분해효소에 비천연 아미노산 4-azido-phenylalanine을 도입하기 위해서는 비천연 아미노산을 아미노아실화할 수 있는 돌연변이 tRNA synthetase와 tRNA를 필요로 한다. 플 라스미드 pEVOL_pAzF는 아라비노스 (arabinose)의 유도에 의하여 돌연변이 tRNA synthetase를 발현한다. 플라스미드 pQE80_UoxR에 클로닝된 재조합 요산분해효소 유전자는 IPTG 에 의하여 발현이 유도된다 [15].
2.2. 배지 조성
재조합 요산분해효소의 발현 및 세포성장을 증대하기 위한 기초 배지 선정을 위하여 3가지 배지 (2 × YT, LB 및 B 배지) 를 사용하여 실험을 수행하였다. LB 배지의 조성은 10 g/L yeast extract, 5 g/L tryptone, 10 g/L NaCl, 2 × YT 배지의 조 성은 16 g/L tryptone, 10 g/L yeast extract, 5 g/L NaCl, 그리고 B 배지의 조성은 12 g/L tryptone, 24 g/L yeast extract, 5 g/L glycerol, 2.3 g/L KH
2PO
4, 12.53 g/L K
2HPO
4를 각각 사용하 였다. 플라스미드 항생제 마커로는 엠피실린(ampicillin)과 클 로람페니콜 (chloramphenicol)을 각각 100 mg/mL과 35 mg/mL 로 첨가하였다. 배양은 50 mL 배지를 포함한 250 mL 플라스 크를 이용하여 37
oC, 250 rpm에서 진행하였다. 비천연 아민 노산이 첨가된 재조합 요산분해효소의 발현을 위하여 배지 에 1 mM 농도의 4-azido-phenylalanine (Sigma-Aldrich, St. Louis, MO, USA )와 1 mM IPTG (Sigma-Aldrich, St. Louis, MO, USA ) 그리고 0.2% (w/v) 아라비노스를 첨가하였다.
세포의 성장은 UV-Vis spectrophotometer (VERSAmax, Molecular Devices, Sunnyvale, CA, USA) 를 이용하여 600 nm에서의 흡광값(OD
600)을 이용하여 측정하였다. 단백질의 발현 양상은 SDS-PAGE를 사용하여 확인하였다 [17].
2.3. 요산분해효소 활성 측정
Urate oxidase 의 활성을 측정하기 위하여 세포 배양액의 OD
600값을 10으로 맞춘 배양액 10 mL을 50 mL tube에 옮기 고 원심분리 (4000 rpm, 4
oC, 15 min)를 통하여 균주를 회수 한 다음, lysis buffer (50 mM sodium phosphate, 0.3 M NaCl) 5 mL을 첨가하여 균체를 현탁 하였다. 이후 초음파파쇄기를 이용하여 15 sec on, 5 sec off, amplification 10%에서 5분간 세포 파쇄를 진행하였다. 세포 파쇄 이후 원심분리 (4000 rpm, 4
oC, 15 min)를 통하여 상등액을 회수하였다. 얻어진 상등액 중에서 10 mL를 reaction buffer (90 mL lysis buffer, 100 mL uric acid solution, 800 mL urate oxidase assay buffer [50 mM sodium borate, 0.2M NaCl pH7.5]) 와 혼합하여 총량 1 mL로 30
oC에서 10분 간격으로 반응시킨 후 293 nm에서 흡광값을 측정하여 소모된 요산의 양을 standard curve를 통하여 계산 하였다. [15] 상등액 속의 단백질의 농도를 정량하기 위하여 Pierce BCA Protein Assay kit (Thermo Scientific, Waltham, MA, USA) 을 사용하였다.
2.4. 반응표면분석법
본 실험에서 진행된 3개의 배지에서 가장 높은 성장을 나타 낸 B 배지를 기본 배지로 선택하고, 배지의 조성을 최적화하 기 위하여 배지 성분 중 tryptone, yeast extract, KH
2PO
4와 K
2HPO
2의 농도를 각기 달리하여 23개의 배지를 제조하였다.
Glycerol의 농도는 탄소원의 영향에 관한 실험에서 최적화를
진행하였고, 나머지 배지 조성의 최적화를 RSM으로 수행하
였다. 각 배지의 조성은 Table 1에 나타내었다. 각 성분별 농
도 범위는 6-18 g/L trypeone, 12-36 g/L yeast extract, 7.41-
22.24 g/L KH
2PO
4와 K
2HPO
2에서 진행하였다. 균주는 B 배
지를 이용하여 37
oC, 250 rpm에서 16시간 전배양을 진행하
였고 이후 1% (v/v)의 비율로 접종하여 23개의 배지가 50 mL 들어있는 250 mL flask에서 각각 본배양을 진행하였다. 이후 세포성장이 0.5 OD
600부근에서 재조합 단백질 발현을 위하여 최종 농도 1 mM IPTG, 1 mM 4-azido-phenylalanine과 0.2%
(w/v) 아라비노스 를 첨가하였다. 37
oC에서 250 rpm에서 7시 간동안 단백질 발현을 진행한 뒤 세포성장과 재조합 요산분 해효소 활성을 측정하였고, 얻어진 데이터를 response surface methodology 방법 (Minitab Inc, State College, PA, USA)을 이 용하여 최적 배지 조성을 찾는 실험을 진행하였다 [18, 19].
3. RESULTS AND DISCUSSION
3.1. 기본배지 선정과 단백질 발현 확인
재조합 요산분해효소 생산을 위한 재조합 대장균의 배양은 37
oC에서 각각 2 × YT, LB 및 B 배지를 이용하여 24시간 진 행하였다. 2 × YT와 LB 배지에는 각각 5g/L glycerol을 첨가 하여 B 배지와 탄소원 농도를 일정하게 하였다. 재조합 대장 균 배양 결과 B 배지에서 가장 높은 세포 성장을 얻을 수 있 었다. SDS-PAGE를 이용한 재조합 요산분해효소 발현을 확 인한 결과, B 배지에서 2 × YT와 LB 배지 보다 높은 단백질 발현 양상을 확인할 수 있었으며, 시간에 따른 발현양은 큰
차이를 보이지는 않았으나 시간이 증가함에 따라 높은 재조 합 요산분해효소 발현을 확인할 수 있었다 (Fig. 1). 이러한 결과를 바탕으로 B 배지를 기본 배지로 하여 탄소원에 따른 성장과 재조합 단백질 발현에 관한 실험을 진행하였다.
B 배지에 탄소원으로 글리세롤과 포도당을 사용한 각각의 경우 세포 성장을 비교한 결과, 포도당을 첨가한 경우 세포 성장이 우수한 것으로 확인이 되었다 (data not shown). 이 후, 포도당 농도를 5, 10, 15, 20 g/L로 설정하고 B 배지를 이용하 여 세포 성장 실험을 진행한 결과 높은 포도당 농도의 배지 조건에서 세포의 성장이 저해됨을 확인하였다 (Fig. 2). 5 g/L 포도당 첨가군에서의 세포 성장은 15 OD
600을 확인할 수 있 었으나, 10 g/L 이상의 포도당이 첨가된 실험군에서 세포의 성장이 8 OD
600수준으로 저해되었다 (Fig. 2(a)). 이 결과는 37
oC와 같은 대장균에서의 최적 성장 온도에서 높은 농도의 포도당을 이용한 배양은 빠른 대사로 인한 많은 유기산 및 세포성장 저해물들의 생산으로 인해 세포의 성장이 저해되 었을 것으로 사료되었다. 좀 더 낮은 배양 온도인 28
oC에서 포도당 농도에 따른 세포 성장 실험을 진행하였다. 그 결과, 28
oC에서 배양 시 10 g/L 포도당 농도에서도 5 g/L 포도당 농 도에서 배양한 결과와 유사하게 19시간 배양 후 16 OD
600수 준의 세포 성장을 확인할 수 있었지만, 포도당이 15, 20 g/L 로 첨가된 실험군에서는 여전히 세포 성장이 저해됨을 확인 Table 1. The medium compositions for RSM experiments
Tryptone Yeast extract K2HPO4 and KH2PO4 Tryptone (g/L) Yeast extract (g/L) K2HPO4 and KH2PO4 (g/L)
1
−
1−
1−
1 6 12 7.412
−
1−
1 0 6 12 14.833
−
1−
1 1 6 12 22.244
−
1 0−
1 6 24 7.415
−
1 0 1 6 24 14.836
−
1 1−
1 6 36 7.417
−
1 1 0 6 36 14.838
−
1 1 1 6 36 22.249 0
−
1−
1 12 12 7.4110 0
−
1 0 12 12 14.8311 0
−
1 1 12 12 22.2412 0 0 0 12 24 14.83
13 0 0 1 12 24 22.24
14 0 1 0 12 36 14.83
15 0 1 1 12 36 22.24
16 1
−
1−
1 18 12 7.4117 1
−
1 0 18 12 14.8318 1
−
1 1 18 12 22.2419 1 0
−
1 18 24 7.4120 1 0 1 18 24 14.83
21 1 1
−
1 18 36 7.4122 1 1 0 18 36 14.83
23 1 1 1 18 36 22.24
할 수 있었다 (Fig. 2(b)).
SDS-PAGE를 이용한 단백질 발현 양상을 확인한 결과 5 g/L 포도당이 첨가된 플라스크 배양군에서 가장 높은 재조합 단 백질의 발현이 확인되었으며, 포도당이 15, 20 g/L 첨가된 플 라스크 배양에서는 상대적으로 적은 발현양이 확인되었다 (Fig. 3). 실제 발현된 단백질의 활성을 특정한 결과 5 g/L 포
도당이 첨가된 배지에서 가장 높은 재조합 요산분해효소 활 성이 얻어졌다 (Fig. 4). 5, 10, 15 g/L 포도당이 첨가된 배지에 서 45.03, 31.1, 25.85 U/mg protein의 재조합 요산분해효소 활성이 각각 얻어졌다.
3.2. 재조합 요산분해효소의 생산 최적화를 위한 배지 성 분의 반응표면분석법
2 × YT, LB 및 B 배지 중에서 B 배지에서 세포 성장과 재조 합 요산분해효소의 발현이 가장 높은 것을 확인하였으며, 탄 소원으로 5 g/L의 포도당이 첨가될 경우 재조합 단백질의 발 현과 활성이 높다는 것을 본 실험에서 확인하였다. 이러한 결과로부터, 재조합 요산분해효소의 생산 최적화를 위해서 실험계획법에 의한 배지 성분 최적화가 필요하였다. 이를 위 Fig. 1. Urate oxidase overexpression in 2 × YT, LB and B media, respectively. (a) total protein, (b) supernatant, (c) pellet (Lane1:
Marker, Lane 2 : 3 h after induction in LB, Lane 3 : 5 h after induction in LB, Lane 4 : 7 h after induction in LB, Lane 5 : 3 h after induction in 2 × YT, Lane 6 : 5 h after induction in 2 × YT, Lane 7 : 7 h after induction in 2 × YT, Lane 8 : 3 h after induction in B, Lane 9 : 5 h after induction in B, Lane 10 : 7 h after induction in B). ← : Urate oxidase protein(34 kDa).
Fig. 2. Growth of E. coli C321 ΔA (pEVOL_pAzF pQE80_UoxR) at different glucose concentrations (a) at 37
oC. (b) at 28
oC (■: 5 g/L Glucose, ▲: 10 g/L Glucose, ●: 15 g/L Glucose, ◆: 20 g/L Glucose).
Fig. 3. Urate oxidase expression in E. coli C321 ΔA (pEVOL_pAzF
pQE80_UoxR) at different glucose concentrations at 28
oC culture
conditions. (Lane1, 6 : 5 g glucose/L without induction, Lane 2, 7
: 5 g glucose/L with induction, Lane 3,8 : 10 g glucose/L with
induction, Lane 4, 9 : 15 g glucose/L with induction, Lane 5, 10 :
20 g glucose/L with induction. ← : Urate oxidase protein(34 kDa).
하여 반응표면분석법을 통한 배지 성분 최적화를 진행하였 다. B 배지 성분에서 tryptone, yeast extract, 그리고 KH
2PO
4와 K
2HPO
4의 농도를 변화시켜 23개의 실험군 배지를 제작하였 으며 (Table 1), 재조합 요산분해효소 활성값을 측정하였다 (Fig. 5). 그 결과 최대 요산분해효소의 활성을 위한 배지성분 들의 가중치 값은 tryptone은 0.0303, yeast extract는 −1, K
2HPO
4와 KH
2PO
4는 0.7778으로 계산이 되었으며 (Table 2), 이 값을 기본 B 배지에 조정하여 12 g/L tryptone, 12 g/L yeast extract, 17.4 g/L KH
2PO
4, 3.195 g/L K
2HPO
4농도에서 최적 의 재조합 요산분해효소의 활성이 얻어지는 것으로 예측이 되었으며, 활성 값은 52.35 U/mg protein을 얻을 수 있을 것으 로 예상하였다. 이를 B2배지로 명명하고 추후 실험에 사용 하였다. 반응표면 분석법으로 얻어진 B2 배지를 제조하여 활성을 특정한 결과 평균 64.92 U/mg protein의 값이 확인되 었다.
3.3. 발현 조건 최적화
5 g/L 포도당이 포함된 B2 배지를 이용하여 비천연 아미노산 과 inducer의 농도에 따른 요산분해효소의 활성을 측정하였 다. 비천연 아미노산으로 사용되는 4-azido-phenylalanine는 0.5, 1, 2, 5 mM 로 첨가하였으며, IPTG 농도는 0.5, 1, 2, 5 mM Fig. 4. Urate oxidase activity of E. coli C321 ΔA (pEVOL_pAzF
pQE80_UoxR) in B medium at different glucose concentrations.
Table 2. Optimal medium composition for B2 media after RSM
Tryptone (g/L) Yeast Extract (g/L) K2HPO4 (g/L) KH2PO4 (g/L)
Value(1) 0.0303 -1 0.7778 0.7778
g/L 12.1818 12 17.4 3.195
Fig. 5. Urate oxidase activity of recombinant of E. coli C321 ΔA (pEVOL_pAzF pQE80_UoxR) in 23 medium compositions.
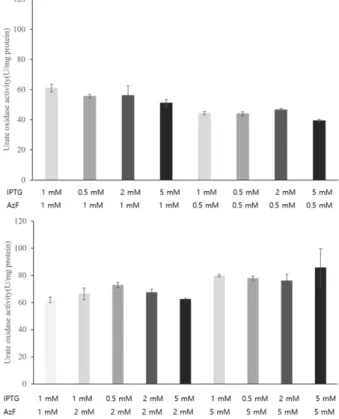
Fig. 6. Urate oxidase activity at different IPTG (0.5, 1, 2 and 5 mM)
and 4-azido-phenylalanine (0.5, 1, 2, and 5 mM) concentrations.
로 첨가하여 진행하였다. Synthetase 발현에 inducer로 작용 하는 아라비노스는 0.2%로 일정하게 유지하였다. 실험결과 IPTG 농도에 따른 요산분해효소의 활성값은 큰 차이를 나타 내지 않았지만, 비천연아미노산인 4-azido-phenylalanine의 첨가 농도가 증가하면 할수록 요산분해효소의 활성이 증가 됨을 확인할 수 있었다. 5 mM의 4-azido-phenylalanine이 첨 가되었을 경우 1 mM의 첨가대비 1.33배 높은 요산 분해효소 의 활성 (85.6 U/mg protein)을 확인할 수 있었다 (Fig. 6). 하 지만 비천연 아미노산의 가격이 비싸고 첨가농도에 비례하 여 요산분해효소의 활성이 급격하게 증가되는 경향성은 나 타나지 않았기 때문에, 경제적으로 요산분해효소의 생산을 위해서는 1 mM 농도의 비천연 아미노산의 첨가와 1 mM 농 도의 IPTG 첨가가 효율적으로 판단되었다.
4. CONCLUSION
본 연구에서는 체내 반감기를 증대시키기 위하여 비천연 아 미노산이 도입된 재조합 요산분해효소의 생산을 위한 배지 및 발현 조전 최적화를 진행하였다. 초기 3가지 기본배지를 비교한 결과 B 배지에서 가장 높은 세포 성장과 재조합 단백 질의 발현이 확인되었으며, 이 후 반응포면분석법을 이용하 여 재조합 요산분해효소의 활성을 최대로 하는 배지 조성을 확인하였다.
최적화된 배지를 이용하여 첨가되는 비천연 아미노산의 농도와 inducer로 사용되는 IPTG 농도에 따른 재조합 단백질 의 활성을 측정하여 가장 효율적인 발현조건을 확인하였다.
본 연구는 향 후 비천연 아미노산이 첨가된 재조합 단백질의 경제적인 대량 생산에 활용될 수 있을 것이다.
Acknowledgements
이 논문은 2018년도 정부(미래창조과학부)의 재원으로 한국 연구재단의 지원 (NRF-2018R1D1A1B07049359), 해양수산 부 GSP사업 (213008-05-4-SB910), 전남대학교 연구년 지원 사업(2018-3367)을 받아 수행된 연구입니다.
REFERENCES
1. Kontermann, R. E. (2011) Strategies for extended serum half-life of protein therapeutics. Curr. Opin. Biotechnol. 22: 868-876.
2. Lund, U., A. Rippe, D. Venturoli, O. Tenstad, A. Grubb, and B.
Rippe (2003) Glomerular filtration rate dependence of sieving of albumin and some neutral proteins in rat kidneys. Am. J. Physiol.
Renal. Physiol. 284: 1226-1234.
3. Kontos, S., and J. A. Hubbel (2012) Drug development: longer- lived proteins. Chem. Soc. Rev. 41: 2686-2695.
4. Elsadek, B., and F. Kratz (2012) Impact of albumin on drug deliv-
ery-new applications on the horizon. J. Control. Release 157: 4-28.
5. Kratz, F., and B. Elsadek (2012) Clinical impact of serum proteins on drug delivery. J. Control. Release 161: 429-445.
6. Sleep, D., J. Cameron, and L. R. Evans (2013) Albumin as a versa- tile platform for drug half-life extension. Biochim. Biophys. Acta 1830: 5526-5534.
7. Duttaroy. A., P. Kanakaraj, B. L. Osborn, H. Schneider, O. K. Pic- keral, C. Chen, G. Zhang, S. Kaithamana, M. Singh, R. Schuling- kamp, D. Crossan, J. Bock, T. E. Kaufman, P. Reavey, M. Carey- Barber, S. R. Krishnan, A. Garcia, K. Murphy, J. K. Siskind, M. A.
McLean, S. Cheng, S. Ruben, C. E. Birse, and O. Blondel (2005) Development of a long-acting insulin analog using albumin fusion technology. Diabetes 54: 251-258.
8. Ikuta. S, V. T. Chuang, Y. Ishima, K. Nakajou, M. Furukawa, H.
Watanabe, T. Maruyama, and M. Otagiri (2010) Abumin fusion of thioredoxin-the production and evaluation of its biological activity for potential therapeutic applications. J. Control. Release 147: 17-23.
9. Osborn, B. L., L. Sekut, M. Corcoran, C. Poortman, B. Sturm, G.
Chen, D. Mather, H. L. Lin, and T. J. Parry (2002) Albutropin: a growth hormone-albumin fusion with improved pharmacokinetics and pharmacodynamics in rats and monkeys. Eur. J. Pharmacol.
456: 149-158.
10. Rosenstock, J., J. Reusch, M. Bush, F. Yang, and M. Stewart, (2009) Potential of albiglutide, a longacting GLP-1 receptor ago- nist, in type 2 diabetes: a randomized controlled trial exploring weekly, biweekly, and monthly dosing. Diabetes Care 32: 1880-1886.
11. Subramanian, G. M., M. Fiscella, A. Lamouse-Smith, S. Zeuzem, and J. G. McHutchison, (2007) Albinterferon alpha-2b: a genetic fusion protein for the treatment of chronic hepatitis C. Nat. Bio- technol. 25: 1411-1419.
12. Zhang R., S. H. Gao, C. Y. Zhu, Y. Sun, X. L. Liu, M. M. Zhao, and C. H. Wang (2019) Characterization of a novel alkaline Arx- ula adeninivorans urate oxidase expressed in Escherichia coli and its application in reducing uric acid content of food. Food Chem.
293: 254-262.
13. Vogt B. (2005) Urate oxidase (rasburicase) for treatment of severe tophaceous gout. Nephrol Dial Transplant 20: 431-433.
14. Jung S, and I. Kwon (2017) Synergistic degradation of a hyperuri- cemia-causing metabolite using one-pot enzyme-nanozyme cas- cade reactions. Scientific Reports 7: 44330.
15. Lim S. I., Y. S. Hahn., and I. Kwon (2015) Site-specific albumina- tion of a therapeutic protein with multi-subunit to prolong activity in vivo. J. Control. Release 207: 93-100.
16. Chin J. W., W. S. Stephen, B. M. Andrew, S. K. Dabid, W. Lei, and G. S. Peter (2002) Addition of p-azido-L-phenylalanine to the genetic code of Escherichia coli. J. Am. Chem. Soc. 124: 9026-9027.
17. Richard, J. S. (2004) Purifying proteins for proteomics: a labora- tory manual. 1st ed., Cold Spring Harbor Laboratory Press. NY.
USA
18. Kim H., C. W. Yun, J. Choi, and S. J. Han (2015) Optimization of medium for protease production by Enterobacteriaceae sp. PAMC 25617 by response surface methodology. Kor. Chem. Eng. Res. 53:
1-6.
19. Kim H. D., Y. K. Im, J. Choi., S. J. Han (2016) Optimization of physical factor for amylase production by Arthrobacter sp. by response surface methodology. Kor. Chem. Eng. Res. 54: 140-144.