ORIGINAL ARTICLE
한반도에서 종 분포 모델을 이용한 두 침입외래식물, 돼지풀과 물참새피의 잠재적 분포 예측
Prediction of Potential Distributions of Two Invasive Alien Plants, Paspalum distichum and Ambrosia artemisiifolia , Using Species Distribution Model in Korean Peninsula
이승현1*ㆍ조강현2ㆍ이우주3
1한국환경공단, 2인하대학교 생명과학과, 3인하대학교 통계학과
SeungHyun Lee1*, Kang-Hyun Cho2 and Woojoo Lee3
1Korea Environment Corporation, Incheon 22689, Korea
2Department of Biological Sciences, Inha University, Incheon 22212, Korea
3Department of Statistics, Inha University, Incheon 22212, Korea
Received 25 August 2016, revised 6 September 2016, accepted 10 September 2016, published online 30 September 2016
ABSTRACT: The species distribution model would be a useful tool for understanding how invasive alien species spread over the country and what environmental variables contribute to their distributions. This study is focused on the potential distribution of two invasive alien species, the common ragweed (Ambrosia artemisiifolia) and knotgrass (Paspalum distichum) in the Korean Peninsula. The maximum entropy (Maxent) model was used for the prediction of their distribution by inferring their climatic environmental requirements from localities where they are currently known to occur. We obtained their presence data from the Global Biodiversity Information Facility and the Korean plant species databases and bioclimatic data from the WorldClim dataset. As a results of the modelling, the potential distribution predicted by global occurrence data was more accurate than that by native occurrence data. The variables determining the common ragweed distribution were precipitation of the driest month and annual mean temperature. Both annual and the coldest quarter mean temperatures were critical factors in determining the knotgrass distribution. The Maxent model could be a useful tool for the prediction of alien species invasion and the management of their expansion.
KEYWORDS: Alien invasive plant, Ambrosia artemisiifolia, Climate, Paspalum distichum, Species distribution model
요 약: 종분포 모델은 어떤 지역에서 침입외래종이 어떻게 확장되고 어떤 환경 요인이 이들의 분포에 영향을 미치는지를 이해하는데 매우 유용한 도구이다. 본 연구에서는 한반도에서 두 침입외래종인 돼지풀 (Ambrosia artemisiifolia)과 물참새피 (Paspalum distichum)의 분포에 대하여 연구하였다. 이 두 종의 현재의 분포지에서 기후환경 요인을 분석하고 이 두 종의 분포를 예측하기 위하여 Maxent (the maximum entropy) 모델을 이용하였다. 이 두 종의 출현 자료는 Global Biodiversity Information Facility와 우리나라의 식물종 데이터베이스에서, 생물기후 자료는 WorldClim 자료로부터 얻었다. 모델을 수행한 결과, 자생지 위치자료를 이용한 예측 결과보다 전지구 위치자료를 이용한 예측이 연구 대상종의 잠재적 분포지를 잘 설명하였다. 이들 종의 분포에 기여한 기후환경 요인으로서 돼지풀에서는 최건월의 강수량과 연평균온도가, 물참새피에서는 연평균온도와 최한사분기의 평균온도가 선정되었다. Maxent 종분포 모델은 외래종의 침입을 예측하고 이들의 확산을 관리하는데 유용한 도구가 될 것으로 생각된다.
핵심어: 외래침입식물, 단풍잎돼지풀, 기후, 물참새피, 종분포 모델
*Corresponding author: [email protected], ORCID 0000-0001-9687-464X
ⓒ Korean Society of Ecology and Infrastructure Engineering. All rights reserved.
This is an open-access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0/), which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
189
1. 서 론
침입외래종 (invasive alien species, IAS)은 정상적으 로는 발견되지 않는 자연 환경에 우연히 또는 의도적으로 도입된 생물로서, 새로운 환경에서 확산 능력이 켜서 생물 다양성을 위협하는 심각한 악영향을 초래한다 (Richardson et al. 2000). 최근 우리나라에서 외래식물은 40과 321종이 분포한다고 보고되었다 (Lee et al. 2011). 특히 이중에서 환경부는 ‘생물다양성 보전 및 이용에 관한 법률’에 의거 하여 생태계를 교란하거나 교란할 우려가 있는 식물 14종 을 ‘생태계교란 생물’로서 지정하여 관리하고 있다. 국내 에서 꾸준히 증가하고 있는 침입외래식물은 고유식물에 대한 경쟁배타에 의한 고유종의 감소와 멸종, 생태계 다양 성 파괴 및 고유 식생 파괴와 교란에 의한 생육 기반 훼손, 교잡에 의한 유전적 혼돈 등의 부정적인 영향을 주는 것으 로 알려져 있다 (Chisholm 2009).
침입외래종을 체계적으로 관리하기 위하여 이미 도입 되었거나 도입될 것을 예상하는 지역에서 이들의 잠재적 인 서식지를 예측하는 것이 중요하다 (Beaumont et al.
2014, Qin et al. 2014). 따라서 최근 외래침입식물의 잠 재적인 확산과 분포를 예측하기 위하여 종 분포 모델을 적용 한 다양한 연구가 활발히 진행되고 있다 (Wang and Wang 2006, Dullinger et al. 2009, Smolik et al. 2010, Tererai and Wood 2014). 종 분포 모델 (species distribution model, SDM)은 종과 환경과의 관계를 파악하기 위한 경 험적 모델로서 생물종의 생태적 지위를 파악하여 새로운 지 역에서 이들의 분포를 파악할 수 있는 기법이다 (Guisan and Thuiller 2005, Elith and Leathwick 2009). 종 분포 모델은 종의 출현 또는 비출현 자료를 이용하는 GLM (generalized linear model), GAM (generalized additive model) 등과 출현자료만을 이용하는 GARP (generic algorithm for rule-set predictions), Maxent (maximum entropy model) 등으로 구분된다 (Elith et al. 2006, Austin, 2007, Zimmermann et al. 2010, Padalia et al.
2014).
종 분포 모델을 이용하여 침입식물의 분포를 예측하는 연구가 활발하게 진해되고 있다. 외래침입식물인 crofton weed (Ageratina adenophora)에 대하여 중국에서 GARP 를 이용하여 잠재적 분포지역을 예측하여 이 종에 대한 위험 지역을 제시하여 외래종 관리에 활용하였으며 (Wang and Wang 2006), 남아프리카에서 Maxent를 이용하여 이 종
의 확산 가능성과 기후적으로 적합한 지역을 예측하고, 실 제 출현자료와 비교하였다 (Tererai and Wood 2014). 한 편 세계적으로 문제가 되고 있는 대표적인 침입외래종인 돼 지풀 (Ambrosia artemisiifolia)에 대한 종 분포 모델을 적 용한 다양한 연구가 보고되고 있다. 오스트리아에서 돼지 풀의 분포를 GLM, GAM, GBMs (generalized boosted regression models)을 이용하여 예측한 결과에서 도입 초기 에 유출된 번식체의 수인 번식체 압력이 이 종의 확산에 중요 한 요인으로 나타났다 (Dullinger et al. 2009). 또한 오스트 리아에서 돼지풀의 분포를 예측하는데 있어서, 종 분포 모델 과 확산 모델을 결합한 통합 모델이 분포지를 보다 정확히 예 측할 수 있었다 (Smolik et al. 2010). 중국에서는 기후변화 시나리오에 따라서 돼지풀과 단풍잎돼지풀 (A. trifida)의 분 포 적합지의 변화를 Maxent를 이용하여 예측하여 이들 종의 관리에 활용하고 있다 (Qin et al. 2014).
한편 국내에서 외래침입식물 연구는 현지조사를 통한 분포연구 (Cho and Shin 2001, Lim and Hwang 2006, Kim et al. 2013)에 중점을 두고 있고, 잠재적 분포를 예측 하기 위하여 종 분포 모델을 적용한 연구는 매우 부족한 실 정이다 (Cho and Lee 2015). 따라서 본 연구의 목적은 환 경부가 지정한 생태계교란 생물 중에서 우리나라에서 분 포지 유형이 상이한 외래침입식물인 돼지풀 (Ambrosia artemisiifolia)과 물참새피 (Paspalum distichum)를 대 상으로 자생지, 전지구, 한반도의 공간규모에서 분포를 파 악하고, 기후 환경요인을 변수로 하는 종 분포 모델을 적용 하여 한반도에서 이들의 잠재적인 분포지역을 예측하고, 이들 식물의 분포에 영향을 주는 기후 환경요인을 식별하 는데 있다.
2. 연구 방법
2.1 연구 대상종 선정
본 연구에서는 생물다양성 보전 및 이용에 관한 법률에 따 라서 생태계교란 생물로 지정된 외래식물 중에서 한반도에서 분포지의 유형이 상이한 것으로 알려진 돼지풀 (Ambrosia artemisiifolia)과 물참새피 (Paspalum distichum)를 조 사대상 식물로 선정하였다. 돼지풀은 한반도 중남부에 넓 게 분포하나 물참새피는 주로 남부지역에서 발견되고 있 다 (NIER 2012).
돼지풀은 국화과로서 키가 약 1 m에 달하는 일년생 초 본식물이다. 이 식물의 자생지는 북미로서 돼지풀 속
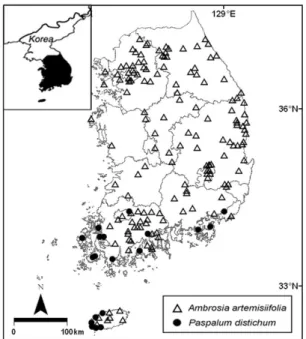
Fig. 1. Map showing the distribution of the study species in
South Korea. Provinces are separated by a solid line. The triangles and circles indicate the locations of Ambrosiaartemisiifolia and Paspalum distichum, respectively.
Table 1. The number of occurrence data for two species
studied in the global and the native area and the Korean Peninsula.Study species
Locations Global
area Native area Korean Peninsula
Ambrosia
artemisiifolia
6,940 1,026 178Paspalum
distichum
6,155 151 32(Ambrosia) 중에서 북미에서 분포지가 가장 넓다 (USDA 2014). 특히 이 식물은 바람에 비산된 꽃가루가 알레르기 를 유발하여 보건 문제를 야기하고 있으며, 농경지에서는 경쟁력이 높은 잡초로서 농업 문제를 유발하고 있다 (EU COST 2014). 한편 유럽, 아시아, 호주, 아프리카, 남미에 서 돼지풀이 외래종으로 도입되어 침입식물로 생태적 문제 를 야기하고 있다 (ISSG 2014). 우리나라에서는 미국으로 부터 도입되어 중부지방으로부터 남부지방으로 확산되어 전국적으로 분포하고 있다 (Fig. 1) (NIER 2012). 이 식물 은 주로 도로변, 하천변, 목초지, 밭, 공터 등에서 대규모 군 락을 이루고 있다.
물참새피는 벼과에 속하며, 지하경과 포복경으로 퍼져 나가 드러눕거나 키가 최대 약 60 cm로 곧게 서는 다년생 초본식물이다. 자생지는 아메리카 열대 지역을 포함한 지 역이다 (Weber 2003, NIER 2012). 자생지에서는 습지 로부터 배수가 양호한 초지까지 생육지 범위가 넓지만 흔 히 목초지, 습초지, 도랑에서 발견된다 (USDA 2014).
현재 물참새피는 세계 각국으로 유입되어 침입종으로 생 태적 문제를 야기하고 있다. 인도의 Keoladeo 국립공원 에서 물참새피는 고유식물을 훼손하는 심각한 종으로 보 고되었고 (Kumar and Mittal 1993, Middleton 1998), 포르투갈과 그리스에서는 자연습지와 수로로 침입하여
다른 고유식물과 물고기에게 피해를 주는 것으로 알려져 있다 (Bernez et al. 2005, Stroh 2006). 우리나라에서는 남부지방에 도입되어 제주도, 전라남도, 전라북도, 경상 남도 등의 주로 남부지방에 분포하고 있다 (Fig. 1) (NIER 2012). 우리나라에서 이 식물은 하천, 습지, 저수 지, 논 등의 물가나 습기가 많은 토양에 군락을 이루어 지 면과 수면을 덮고 있다.
2.2 식물 위치 자료의 수집
연구 대상종인 돼지풀과 물참새피가 국내에 도입된 외 래식물인 점을 착안하여 위치자료를 자생지와 전지구 (한 반도 위치자료 제외)로 구분하여 세계생물다양성정보기 구 (GBIF 2014)에서 제공하는 표본자료의 위치정보를 수집하였다. 두 종의 자생지 위치자료 중 돼지풀은 플로리 다, 캘리포니아, 루이지애나, 미주리, 아칸소, 코네티컷, 콜 로라도, 아이오와, 펜실베니아, 미시시피, 조지아, 퀘벡, 온 타리오, 뉴저지, 텍사스, 캔사스, 버지니아, 미시간, 뉴욕, 미네소타, 워싱턴 주 등 북미 일대, 물참새피는 에콰도르, 아르헨티나, 볼리비아, 브라질, 칠레, 콜롬비아, 페루, 우루 과이, 가이아나, 베네수엘라 국가 등 중남미 일대에서 수집 된 자료를 사용하였다.
또한 자생지와 전지구 위치 자료를 이용하여 만들어진 두 종의 종 분포 모델을 검증하기 위하여, 한반도에서 이 들의 위치 자료는 국립수목원의 국가생물종지식정보시 스템 (KNA 2014)에서 제공하는 표본자료를 참고하였 고, 국립환경과학원에서 발행한 생태계교란생물 자료집 (NIER 2012)의 분포지 도면에서 출현위치를 육안으로 확인 후 QGis DT (2014)를 이용하여 경위도 좌표를 수집 하였다 (Table 1).
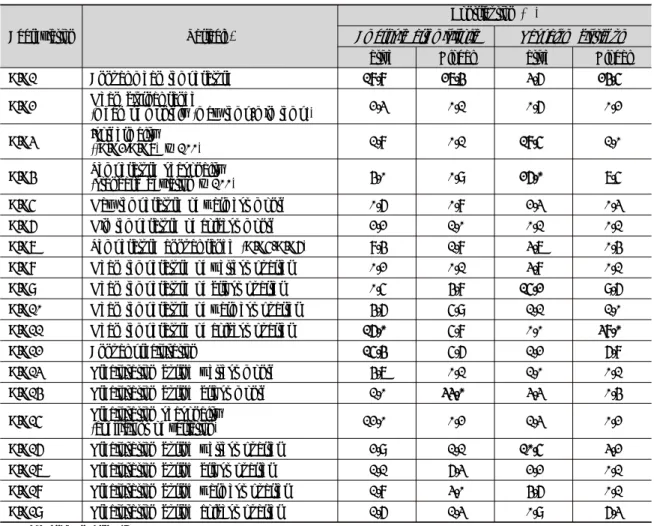
Table 2. The percentage contribution of the bioclimatic variables used in the Maxent model of the two alien species
based on their native and global occurrence data. The values are averages with 100 replicate runs. The bold values indicate the variables that had over 10% in contribution for each species.Abbreviation Variable*
Contribution (%)
Ambrosia artemisiifolia Paspalum distichum
Native Global Native Global
BIO1 Annual mean temperature 18.8 27.4 3.6 24.5
BIO2 Mean diurnal range
(mean of monthly (max temp-min temp)) 2.3 0.1 0.6 0.2
BIO3 Isothermality
((BIO2/BIO7) × 100) 1.8 0.1 18.5 1.0
BIO4 Temperature seasonality
(standard deviation × 100) 4.0 0.9 26.0 7.5
BIO5 Max temperature of warmest month 0.6 0.8 2.3 0.3
BIO6 Min temperature of coldest month 2.2 1.0 0.1 0.1
BIO7 Temperature annual range (BIO5-BIO6) 8.4 1.8 3.7 0.4
BIO8 Mean temperature of wettest quarter 0.2 0.1 3.8 0.1
BIO9 Mean temperature of driest quarter 0.5 4.8 15.2 9.6
BIO10 Mean temperature of warmest quarter 4.6 5.9 1.1 1.0
BIO11 Mean temperature of coldest quarter 16.0 5.8 0.0 38.0
BIO12 Annual precipitation 15.4 5.6 1.2 6.8
BIO13 Precipitation during wettest month 4.7 0.1 1.0 0.1
BIO14 Precipitation during driest month 1.0 33.0 3.3 0.4
BIO15 Precipitation seasonality
(coefficient of variation) 12.0 0.2 1.3 0.2
BIO16 Precipitation during wettest quarter 2.9 1.1 10.5 3.2
BIO17 Precipitation during driest quarter 1.1 6.3 2.2 0.1
BIO18 Precipitation during warmest quarter 1.8 3.0 4.6 0.1
BIO19 Precipitation during coldest quarter 1.6 1.3 0.9 6.3
* Data from WorldClim (2014)
2.3 환경 자료의 수집
종 분포 모델을 구축하기 위한 환경변수는 기후 환경요 인만을 사용하였다. 이 기후 자료는 WorldClim (2014)에 서 제공하는 공간 해상도 5 km × 5 km의 19개 생물기후 변수를 이용하였다 (Table 2). 이들 생물기후 변수는 생물 학적으로 의미 있는 기후 요인으로서 크게 온도와 강수량 으로 구분된다. 온도 변수로서는 연평균온도 (BIO1), 평 균일교차 (BIO2), 등온성 (BIO3) (BIO2/BIO7 × 100), 온도계절성 (BIO4) (표준편차 × 100), 최난월의 최고온 도 (BIO5), 최한월의 최저온도 (BIO6), 연온도범위 (BIO7) (BIO5 – BIO6), 최습사분기의 평균온도 (BIO8), 최건사분기의 평균온도 (BIO9), 최난사분기의 평균온도 (BIO10) 및 최한분기의 평균온도 (BIO11)이었다. 강수
량 변수로서는 연강수량 (BIO12), 최습월의 강수량 (BIO13), 최건월의 강수량 (BIO14), 강수량계절성 (BIO15) (변동계수), 최습사분기의 강수량 (BIO16), 최 건사분기의 강수량 (BIO17), 최난사분기의 강수량 (BIO18) 및 최한분기의 평균온도 (BIO19)이었다. 여기 에서 생물기후변수의 사분기 (quarter)는 일년을 4등분한 3개월씩의 기간을 의미한다.
2.4 종 분포 모델링
본 연구에서는 연구대상종의 위치자료가 출현지점만 을 나타내고 있기 때문에 출현자료만을 적용할 때 다른 모 델에 비교하여 비교적 정확도가 높은 Maxent를 적용하였 다 (Phillips et al. 2006, Yang et al. 2013). Maxent 수행
을 위한 프로그램은 Maxent 3.3.3k (Phillips 2013)를 이용 하였다. Maxent 모델의 성능을 검증하기 위하여 연구 대상종 의 모든 출현자료를 이용하여 10차 교차검증 (10-fold cross validation)을 실시하였다. 모델의 과적합 (over-fitting)을 피 하기 위하여 정규화 승수 값 (regularization multiplier value) 을 정하는 것이 필요한데, 이를 위하여 Maxent의 기본값인 1을 이용하였고, 환경변수의 최대 지점수는 10,000개로 설정하였 다. 기후환경 변수의 중요도를 평가하기 위하여 잭나이프 (jackknife) 기법을 사용하였으며, 반응곡선 (response curves)을 토대로 각 기후환경변수 값과 분포확률과의 관계 를 파악하였다. Maxent 모델의 출력형식 (output format) 은 분포확률의 의미로 해석이 가능한 로지스틱 형식 (logistic format, 최저값 0 및 최고값 1로 설정)을 선택하였 으며, 모델에 의하여 자동으로 생성된 연구 대상종의 잠재적 분포도를 QGis DT (2014)와 ArcGIS (ESRI, Redlands, CA, 미국)을 병행하여 재구성하였다.
모델의 정확도는 교차 검증으로부터 구해지는 수신자 조작 특성 곡선 (ROC, receiver operating characteristic curve)의 아랫부분 면적 (AUC, area under the curve)으 로서 판단하였다. AUC는 0.5를 기준으로 하여 1.0이면 모델이 완벽하며, 0.8 이상이면 모델 예측이 비교적 잘 이 루어졌다고 판단한다 (Swets 1988). 분석 결과의 안정성 을 살펴보기 위해 Maxent의 추가적인 옵션인 ‘random test percentage’를 통하여 검증 자료 (test data)를 25%, 훈련 자료 (training data)를 75%로 설정하였다. 사용할 수 있는 모든 자료를 이용하여 모델의 성능을 검증하기 위 하여 10차 교차검증 (10-fold cross validation), 부트스 트랩 (bootstrap)과 부표본추출 (subsampling)을 각 100 회 반복(replicates)하여 AUC를 구하였다. 또한, 정확한 예측결과를 도출하기 위해서는 100회 반복된 예측결과의 수렴 (convergence)을 위한 적절한 시간을 가지는 것이 매우 중요하며, Maxent의 추가적인 옵션인 ‘maximum iterations’을 통하여 최적화된 알고리즘을 찾기 위한 반 복수 (number of iterations)의 설정이 가능하다. 본 연 구에서는 반복수를 Maxent의 기본값인 500이 아닌 5,000으로 증가시켰다. 마지막으로 한반도 위치자료를 이용하여 자생지 및 전지구 위치자료를 이용하여 도출된 모델을 검증하였다.
3. 결과
3.1 돼지풀의 분포
기후환경요인을 변수로 하여 Maxent를 적용한 돼지풀 의 종 분포 모델에서 10차 교차검증을 이용한 수신자 조작 특성 곡선의 아랫부분 면적인 AUC는 자생지 위치자료를 이용하면 0.948, 전지구 위치자료를 이용하면 0.841로서 적합도가 높았다 (Fig. 2). 또한 부트스트랩과 부표본추출 을 이용한 AUC는 자생지 위치자료에서는 각각 0.963 및 0.951이었고, 전지구 위치자료에서는 각각 0.864 및 0.858으로서 본 모델 결과가 매우 안정한 것을 확인할 수 있었다 (Fig. 2).
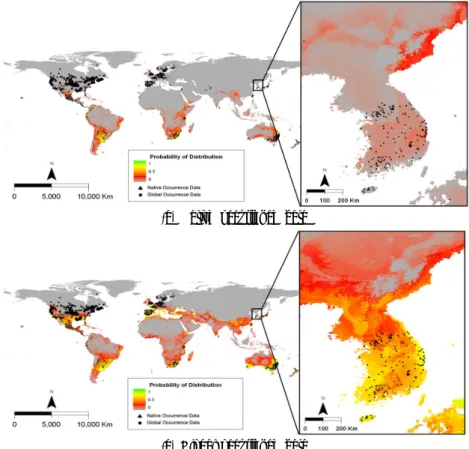
돼지풀의 자생지인 북미의 위치자료를 이용한 종 분포 모델에서는 자생지를 제외한 다른 지역에서는 이들이 도 입된 실제 분포지와 모델에 의한 생육가능지역의 차이가 크게 나타났다 (Fig. 3). 특히 돼지풀이 도입되어 분포하는 대부분의 중남미에서는 서식하지 못하는 것으로 예측되었 다. 그러나 한반도를 제외한 전지구 위치자료를 이용한 종 분포 모델에서는 모델에 의한 예측이 실제 분포를 잘 설명 하고 있었다. 돼지풀은 고위도의 북반구와 건조지역을 제 외한 나머지 지역에서 광범위하게 생육이 가능할 것으로 예측되었다.
자생지 위치자료를 이용하여 한반도에서 돼지풀의 생 육가능지역을 예측한 결과, 돼지풀은 한반도의 북부 동해 안과 제주도와 남부 해안을 제외한 중남부 지역에 낮은 확 률로 분포할 것으로 예측되었다 (Fig. 3). 반면에 전지구 위치자료를 이용한 경우에는 우리나라 북부 산악지역을 제외하고 전체 지역에 분포하는 것으로 나타났고 특히 제 주도와 한반도의 남부 해안을 따라서 분포 확률이 높았다. 한반도에서 실제 보고된 위치자료와 중첩 분석하면, 자생 지 위치자료보다 전지구 위치자료를 이용한 모델 결과가 돼지풀의 생육지를 잘 설명하였다.
돼지풀의 종 분포 모델에 적용된 생물기후 변수 중에서 기여도가 높은 변수는 자생지 위치자료를 이용한 모델에서 는 연평균온도 (BIO1), 최한 사분기의 평균온도 (BIO11), 연강수량 (BIO12) 및 강수량의 계절성 (BIO15)이었고, 전지구 위치자료에서는 최건월의 강수량 (BIO14)과 연평 균온도 (BIO1)가 총 60% 이상의 변이를 설명하였다 (Table 2). 실제 한반도 분포지 자료와 보다 적합한 전지구 위치자료를 이용한 변수반응곡선을 살펴보면 최건월의 강 수량이 0 mm 이상인 지역에서 분포확률이 증가하고 약
(a) Cross-validation with native occurrence data (b) Cross-validation with global occurrence data
(c) Bootstrap with native occurrence data (d) Bootstrap with global occurrence data
(e) Subsampling with native occurrence data (f) Subsampling with global occurrence data
Fig. 2. The area under the curve (AUC) of the receiver operating characteristic (ROC) of the species distribution model in
Ambrosia artemisiifolia using data set from the native and global ranges. Cross-validation, bootstrap, and subsampling
techniques are used in the model for the replicates. The lines indicate mean values for the 100 replicates of the models and grey shading indicates their standard deviations.50 mm 이상에서 최상에 도달하였고, 연평균온도가 0 °C 이상인 지역에서 분포확률이 증가하는 경향을 보였고 25
°C 이상인 지역에서 분포확률이 가장 높았다 (Fig. 4).
3.2 물참새피의 분포
물참새피에서 10차 교차검증을 이용한 자생지 위치자 료에 대한 AUC는 0.966, 한반도를 제외한 전지구 위치자 료에 대한 AUC는 0.891이었고, 부트스트랩과 부표본추 출을 이용한 AUC는 자생지 위치자료를 이용하면 각각
0.987 및 0.965, 전지구 위치자료를 이용하면 각각 0.908 및 0.900으로서 본 모델 결과가 매우 안정한 것을 확인할 수 있었다 (Fig. 5).
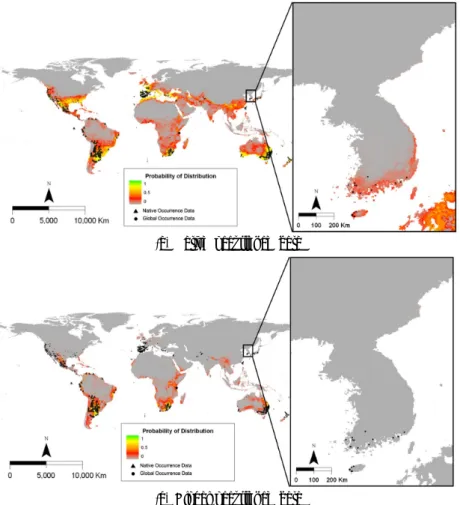
물참새피의 자생지인 중남미의 위치자료를 이용한 종 분포 모델에 의하여 예측된 전지구에서 생육가능지는 실 제 분포지와 큰 차이를 나타내었다 (Fig. 6). 특히 물참새 피가 도입되어 실제 분포하는 북미, 서유럽, 동아시아에서 는 생육하지 못하는 것으로 예측되었다. 한편 한반도를 제 외한 전지구 위치자료를 이용한 종 분포 모델에서는 모델 에 의한 생육가능지가 실제 분포지를 포함하여 보다 넓은
(a) Native occurrence data
(b) Global occurrence data
Fig. 3. Map showing the potential distribution of Ambrosia artemisiifolia predicted by the Maxent model using the data
set from native and global ranges.(a)
Ambrosia artemisiifolia
(b)
Paspalum distichum
Fig. 4. Environmental response curve for bioclimatic variables contributing greater than 10% to the species distribution
models of two alien plants using the global occurrence data. The lines indicate mean values for the 100 replicates of the models and grey shading indicates their standard deviations.(a) Cross-validation with native occurrence data (b) Cross-validation with global occurrence data
(c) Bootstrap with native occurrence data (d) Bootstrap with global occurrence data
(e) Subsampling with native occurrence data (f) Subsampling with global occurrence data
Fig. 5. The area under the curve (AUC) of the receiver operating characteristic (ROC) of the species distribution model
in Paspalum distichum using data set from the native and global ranges. Cross-validation, bootstrap, and subsampling techniques are used in the model for the replicates. The lines indicate mean values for the 100 replicates of the models and grey shading indicates their standard deviations.
범위를 차지하였다. 물참새피는 고위도의 북반구와 건조 지역을 제외한 나머지 지역에서 광범위하게 생육이 가능 할 것으로 예측되었다.
자생지 위치자료를 이용한 종 분포 모델에서 물참새피 는 한반도에서 생육이 불가능할 것으로 예측되었다 (Fig.
6). 전지구 위치자료를 이용한 경우에는 이 식물의 생육가 능지는 제주도, 전라남도와 경상남도의 남해안 일대로 한 정되었으며, 경상북도의 동해안 일대에서도 1% 미만의 낮은 분포확률이지만 서식이 가능할 것으로 예측되었다.
이러한 예측 결과는 이 식물의 실제 위치자료와 대부분 일 치하는 경향으로 보였다 (Fig. 6).
물참새피의 자생지 위치자료를 이용한 모델이 온도계 절성 (BIO4), 등온성 (BIO3), 최건 사분기의 평균 온도 (BIO9) 및 최습 사분기의 강수량 (BIO8)이 전체 변이의 70% 이상을 기여하였고, 전지구 위치자료를 이용한 경우 에 최한 사분기 평균온도 (BIO11)와 연평균온도 (BIO1) 가 60% 이상의 변이를 설명하였다 (Table 2). 그러므로 물참새피 종 분포 모델에서는 온도에 관련된 변수가 주요
(a) Native occurrence data
(b) Global occurrence data
Fig. 6. Map showing the potential distribution of Paspalum distichum predicted by the Maxent model using the data
set from native and global ranges.환경요인이었다. 한반도 위치자료와 가장 적합한 전지구 위치자료를 이용한 모델의 변수반응곡선을 분석한 결과, 최한 사분기 평균온도가 0 °C이상 30 °C 미만인 지역에서 물참새피의 분포확률이 높았고, 연평균온도가 20 °C 이상 인 지역에서는 분포확률이 감소하는 경향이었다 (Fig. 4).
4. 논의
본 연구에서는 표본 위치정보와 기후 환경요인을 변수로 하는 Maxent 방법을 적용하여, 우리나라에서 분포 유형이 상이한 외래침입식물인 돼지풀과 물참새피의 잠재적인 분 포지역을 예측하고자 하였다. 두 종에 적용한 Maxent 방법 에 의하여 도출된 AUC (수신자 조작 특성 곡선의 아랫부 분 면적) 값이 모두 0.8 이상이어서 예측 모델이 적합하다 고 판단되었다 (Fig. 2 and 5) (Thuiller et al. 2005). 특히 자생지 위치자료만을 이용한 모델이 전 지구 위치자료를
이용한 모델보다 높은 예측도를 보였는데, 일반적으로 국 소적으로 분포하는 종에 대한 종 분포 모델의 AUC가 광 범위하게 분포하는 종보다 큰 것으로 알려져 있다 (Yang et al. 2013).
연구 대상종이 외래식물인 점에 착안하여 이들의 자생 지에서 분포 자료를 이용하여 한반도의 생육가능지역을 파악하고자 하였다. 그러나 자생지 위치 정보를 이용하여 한반도에서 돼지풀과 물참새피의 잠재적 분포를 예측한 것이 이들 식물이 도입된 지역까지 포함한 전세계에서 위 치 정보를 이용한 것보다 실제 한반도에서 이들의 분포를 잘 설명하지 못하였다. 종 분포 모델에서 생물 출현자료가 지리적으로 편향되면 예측된 잠재적 서식지와 실제 분포 지에 큰 차이를 유발할 수 있다 (Barnes et al. 2014). 한편 이러한 결과는 외래식물인 돼지풀과 물참새피의 기본지위 (fundamental niche) 폭이 이들의 자생지에서 추정한 실 현지위 (realized niche) 폭보다 넓다는 것과 이들 식물이
도입된 환경에서 이들의 표현형 적응성과 적합도가 증가 한다는 것을 생각할 수 있다 (Chisholm 2009). 즉 외래식 물은 이들의 원산지 환경요인만을 고려하여 한반도에서의 분포지를 예측하면 그 분포가능지가 실제 분포지와 적합 하지 않을 수가 있다는 것을 의미한다. 그러므로 이들 외래 종의 실현지위가 아닌 기본지위를 폭넓게 파악하여야 분 포가능지역을 올바르게 예측할 수 있을 것으로 생각된다.
기후 요인을 고려한 두 종 외래식물의 한반도에서 잠재 분포지는 상이하였는데, 물참새피는 주로 한반도 남쪽 해 안에 제한적으로 분포하고 돼지풀은 북부 산악지역을 제 외한 한반도 전역에 분포하는 것으로 예측되었다. 이러한 잠재분포지 차이는 이 두 종의 기후 환경 요인에 대한 반응 이 다르기 때문이다. 물참새피는 겨울의 온도가 낮지 않은 난대성 기후에 분포하지만 연평균 기온이 너무 높은 기후 대에서는 분포 확률이 낮아지는 것으로 판단된다 (Fig.4).
물참새피 종자가 겨울에 생존하기 위한 가장 중요한 환경 요인이 온도인 것으로 보고되어 있다 (Shibayama 1988).
또한 종자발아의 최적온도가 20-40 °C (Okuma and Chikura 1984) 또는 28-35 °C (Huang and Hsiao 1987) 이며, 이 온도 범위에서 휴면 종자의 생존확률이 높은 것으 로 알려져 있다 (Okuma and Chikura 1984). 따라서 물참 새피 종자의 생존에는 겨울 온도가 중요하고 종자 발아율도 온난하여야 높다는 것을 알 수 있다. 한편 물참새피의 분포 확률이 연평균기온이 20 °C 이상인 곳에서 낮아지는 것은 연평균온도가 높은 곳에 사막 등의 건조지가 분포하며 이곳 에서 물가에 생육하는 물참새피가 분포하기 어렵기 때문인 것으로 생각된다.
물참새피와는 대조적으로 돼지풀은 온도에 대한 반응 은 둔감하지만 가장 건조한 달의 강수량이 50 mm 이상인 지역에서 분포확률이 높았다 (Fig. 6). 돼지풀 종자의 발아 온도에 대한 연구를 살펴보면, 발아 적정온도가 12-32℃
(Cha et al. 2002)와 5-25 °C (Leiblein-Wild et al. 2014) 로서 물참새피에 비하여 낮아서 저온에서도 발아가 용이 할 것으로 생각된다. 또한 발아에 미치는 수분 조건을 살펴 보면, 토양함수량이 7% 미만이면 발아가 전혀 이루어지 지 않았고 토양함수량이 15%에서도 종자 중 4%만이 발 아에 성공하여 (Cha et al. 2002), 돼지풀의 분포에는 발아 시기에 토양의 수분조건이 매우 중요할 것으로 생각된다.
우리나라의 경우 가장 건조한 달은 4월이고 (KMA 2012), 돼지풀의 발아와 초기 생장이 이루어지는 시기인 4-5월에는 토양함수량이 5-15%이므로 (Lee et al.
1997), 이 시기에 강수에 의한 토양 수분 공급이 돼지풀의 분포에 중요한 요인으로 작용할 것으로 생각된다. 본 연구 에서도 최건월의 강수량이 돼지풀의 분포를 결정하는 중 요한 요인으로 선정되었으며, 중국에서 돼지풀의 잠재분 포지 예측에서도 이 종의 분포에 기여한 환경변수가 고도 와 더불어 4월 평균 강수량, 10월 평균 강수량 및 연 강수량 으로 나타났다 (Qin et al. 2014).
본 연구에서는 표본 위치정보와 기후 환경의 자료를 이 용하여 종 본포 모델로 한반도에서 돼지풀과 물참새피의 잠재 분포지를 예측하였는데, 이 두 종의 현재 분포지가 서 로 상이한 것을 잘 설명하고 있었다. 그러나 기후 환경요인 만을 환경변수로 사용한 한계를 가지고 있었다. 그러므로 외래종의 잠재 생육지를 보다 잘 추정하기 위해서는 기후 환경과 더불어 지형, 지질, 토양, 토지이용, 인위적 교란 등 환경 요인을 고려하여야 할 것으로 생각된다. 또한 새로운 환경에 도입된 외래침입식물의 잠재적 분포지역을 예측하 기 위해서 이들의 자생지에서 파악한 생육 환경 자료만으 로는 예측된 분포 가능지가 제한되는 것으로 나타났다. 이 러한 한계를 극복하기 위해서는 대상 식물에 대한 생리생 태 및 지리분포에 대한 역사적인 연구가 병행되어야 할 것 으로 생각된다. 결론적으로 다양한 환경 요인을 반영하고 정확한 생태적 지위를 파악한 종 분포 모델을 이용하면, 도 입되었거나 도입될 것이 예상되는 지역에서 침입외래식물 의 잠재적 분포지를 보다 정확하게 예측할 수 있을 것으로 판단된다. 또한 한반도에서 외래식물의 분포 가능지를 도 입 전후에 미리 예측한다면 이들 식물을 관리하는데 중요 한 정보를 제공할 수 있을 것으로 생각된다.
References
Austin, M. 2007. Species distribution models and ecological theory: A critical assessment and some possible new approaches. Ecological Modelling 200: 1-19
Barnes, M.A., Jerde, C.L., Wittmann, M.E., Chadderton, W.L., Ding, J., Zhang, J., Purcell, M., Budhathoki, M.
and Lodge, D.M. 2014. Geographic selection bias of occurrence data influences transferability of invasive Hydrilla verticillata distribution models. Ecology and Evolution 4: 2584-2593.
Beaumont, L.J., Gallagher, R.V., Leishman, M.R., Hughes, L. and Downey, P.O. 2014. How can knowledge of the climate niche inform the weed risk assessment process? A case study of Chrysanthemoides
monilifera in Australia. Diversity and Distributions 20:
613-625.
Bernez, I., Ferreira, M.T., Albuquerque, A. and Aguiar, F.
2005. Relations between river plant richness in the Portuguese floodplains and the widespread water knotgrass (Paspalum paspalodes). Hydrobiologia 551:
121-130.
Cha, S.H., Kim, W.H. and Kim, J.H. 2002. Effects of some environmental factors on the germination of seeds in Ambrosia arfemisiifolia var. elatior. Journal of Ecology and Environment 25: 163-170. (in Korean) Chisholm, R. 2009. The ecology, economics, and
management of alien invasive species. In, Levin, S.A.
(ed.), The Princeton Guide to Ecology. Princeton University Press. New Jersey, USA. pp. 731-739.
Cho, K.-H. and Lee, S.H. 2015. Prediction of changes in the potential distribution of a waterfront alien plant, Paspalum distichum var. indutum, under climate change in the Korean Peninsula. Ecology and Resilient Infrastructure 2: 206-215. (in Korean)
Cho, K.H. and Shin, D.H. 2001. Vegetation structure and distribution of exotic plants with geomorphology and disturbance in the riparian zone of Seunggi Stream, Incheon. Journal of Ecology and Environment 24:
273-280. (in Korean)
Dullinger, S., Kleinbauer, I., Peterseil, J., Smolik, M. and Essl, F. 2009. Niche based distribution modelling of an invasive alien plant: Effects of population status, propagule pressure and invasion history. Biological Invasions 11: 2401-2414.
Elith, J. and Leathwick, J.R. 2009. Species distribution models: Ecological explanation and prediction across space and time. Annual Review of Ecology, Evolution, and Systematics 40: 677-697.
Elith, J., Graham, C.H., Anderson, R.P., Dudík, M., Ferrier, S., Guisan, A., Hijmans, R.J., Huettmann, F., Leathwick, J.R., Lehmann, A., Li, J., Lohmann, L.G., Loiselle, B.A., Manion, G., Moritz, C., Nakamura, M., Nakazawa, Y., Overton, J.M., Peterson, A.T., Phillips, S.J., Richardson, K., Scachetti-Pereira, R., Schapire, R.E., Soberón, J., Williams, S., Wisz, M.S. and Zimmermann, N.E. 2006. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 29: 129-151.
EU COST. 2014. Sustainable Management of Ambrosia artemisiifolia in Europe (SMARTER). European Cooperation in Science and Technology. http://www.rag weed.eu. Assessed 26 October 2014.
GBIF. 2014. The global biodiversity information facility.
http://www.gbif.org. Assessed 16 January 2014.
Guisan, A. and Thuiller, W. 2005. Predicting species distribution: offering more than simple habitat models.
Ecology letters 8: 993-1009.
Huang, W.Z and Hsiao, A.I. 1987. Factors affecting seed dormancy and germination of Paspalum distichum.
Weed Research, UK 27: 405-415.
ISSG. 2014. The Global Invasive Species Database. The Invasive Species Specialist Group, the Species Survival Commission (SSC) of the International Union for Conservation of Nature (IUCN). http://www.issg.org/
database/welcome/. Assessed 13 October 2014.
Kim, Y.H., Kil, J.H., Hwang, S.M. and Lee, C.W. 2013.
Spreading and distribution of Lactuca scariola, invasive alien plant, by habitat types in Korea. Weed & Turfgrass Science 2: 138-151. (in Korean)
KMA. 2012. Climatological Normals of Korea (1981- 2010). Korea Meteorological Administration, Seoul, Korea. (in Korean)
KNA. 2014. Korea Biodiversity Information System.
Korea National Arboretum. http://www.nature.go.kr.
Assessed 22 January 2014. (in Korean)
Kumar, C.R.A and Mittal, D.D. 1993. Habitat preference of fishes in wetlands in relation to aquatic vegetation and water chemistry. Journal of the Bombay Natural History Society 90: 181-192.
Lee, H.J., Kim, Y.O. and Chang, N.K. 1997. Allelopathic effects on seed germination and fungus growth from the secreting substances of some plants. Journal of Ecology and Environment 20: 181-189.
Lee, Y.M., Park, S.H., Jung, S.Y., Oh, S.H. and Yang, J.C. 2011. Study on the current status of naturalized plants in South Korea. Korean Journal of Plant Taxonomy 41: 87-101. (in Korean)
Leiblein-Wild, M.C., Kaviani, R. and Tackenberg, O.
2014. Germination and seedling frost tolerance differ between the native and invasive range in common ragweed. Oecologia 174: 739-750.
Lim, D.O and Hwang, I.C. 2006. Exotic plants and conservation in Gayasan National Park. Korean Journal of Environment and Ecology 20: 281-288. (in Korean) Middleton, B.A. 1998. The water buffalo controversy in
Keoladeo National Park, India. Ecological Modelling 106: 93-95.
NIER. 2012. Invasive Alien Species. National Institute of Environmental Research, Incheon, Korea. (in Korean) Okuma, M and Chikura, S. 1984. Ecology and control of
subspecies of Paspalum distichum L., Chikugo suzumenohie, growing in creeks in the paddy area on the lower reaches of the Chikugo River in Kyushu. 4.
Possibility of reproduction by seeds. Weed Research, Japan 29: 45-50.
Padalia, H., Srivastava, V. and Kushwaha, S. 2014. Modeling potential invasion range of alien invasive species, Hyptis suaveolens (L.) Poit. in India: Comparison of MaxEnt and GARP. Ecological Informatics 22: 36-43.
Phillips, S.J. 2013. Maxent Software for Species Habitat Modeling, version 3.3.3k. http://www.cs.princeton.
edu/~schapire/maxent/. Assessed 13 October 2014.
Phillips, S.J., Anderson, R.P. and Schapire, R.E. 2006.
Maximum entropy modeling of species geographic distributions. Ecological Modelling 190: 231-259.
QGis DT. 2014. Quantum gis geographic information system. http://www.qgis.org. Assessed 26 October 2014.
Qin, Z., DiTommaso, A., Wu, R.S. and Huang, H.Y.
2014. Potential distribution of two Ambrosia species in China under projected climate change. Weed Research, UK 54: 520-531.
Richardson, D.M., Pyšek, P., Rejmánek, M., Barbour, M.G., Panetta, F.D. and West, C.J. 2000. Naturalization and invasion of alien plants: concepts and definitions.
Diversity and Distributions 6: 93-107.
Shibayama, H. 1988. Ecology of aquatic weeds in creeks of the paddy growing area in the lower reaches of the Chikugo river. Bulletin of the Kyushu National Agricultural Experiment Station 25: 1-75.
Smolik, M.G., Dullinger, S., Essl, F., Kleinbauer, I., Leitner, M., Peterseil, J., Stadler, L.-M. and Vog, G.
2010. Integrating species distribution models and interacting particle systems to predict the spread of an invasive alien plant. Journal of Biogeography 37:
411-422.
Stroh, H.G. 2006. Contribution to the ephemeral wetland
vegetation along riverbanks and lakeshores of Western Thrace (NE Greece). Tuexenia 26: 353-388.
Swets, J.A. 1988. Measuring the accuracy of diagnostic systems. Science 240: 1285-1293.
Tererai, F and Wood, A.R. 2014. On the present and potential distribution of Ageratina adenophora (Asteraceae) in South Africa. South African Journal of Botany 95 (5):152-158.
Thuiller, W., Richardson, D.M., Pyšek, P., Midgley, G.F., Hughes, G.O. and Rouget, M. 2005. Niche‐based modelling as a tool for predicting the risk of alien plant invasions at a global scale. Global Change Biology 11:
2234-2250.
USDA. 2014. The PLANTS Database. National Plant Data Team, United States Department of Agriculture, Greensboro, NC 27401-4901, USA. http://plants.usda.
gov.Assessed 26 October 2014.
Wang, R and Wang, Y.Z. 2006. Invasion dynamics and potential spread of the invasive alien plant species Ageratina adenophora (Asteraceae) in China. Diversity and Distributions 12: 397-408.
Weber, E. 2003. Invasive Plant Species of the World: a Reference Guide to Environmental Weeds. CABI Publishing, Wallingford, UK.
WorldClim. 2014. WorldClim-Global Climate Data.
http://www.worldclim.org. Assessed 13 October 2014.
Yang, X.Q., Kushwaha, S.P.S., Saran, S., Xu, J.C. and Roy, P.S. 2013. Maxent modeling for predicting the potential distribution of medicinal plant, Justicia adhatoda L. in Lesser Himalayan foothills. Ecological Engineering 51:
83-87.
Zimmermann, N.E., Edwards Jr., T.C., Graham, C.H., Pearman, P.B. and Svenning, J.-C. 2010. New trends in species distribution modelling. Ecography 33:
985-989.