Introduction
In the past decade, the prevalence of overweight and obesity has increased dramatically.
1,2)Currently, 65% of United States adults are overweight and 31% of adults are obese.
1)Obesity has known to cause insulin resistance, Type 2 diabetes, cardiovascular disease, high blood pressure and even certain types of cancer.
3-5)Recent data from the Centers for Disease Control reported that Type 2 diabetes, cardiovascular disease, various forms of cancer combine to make up to 70% of all deaths in the United States.
5)Recently, Olshansky et al. predicted that steady increase in life expectancy seen last centuries will soon decline due to prevalence of obesity.
6)Therefore, it is important to understand the cause and proper treatment or prevention of obesity.
The past ten years have been the golden age of obesity research.
7)While long understood that the amount of body fat is determined by an interaction of genes and environment, we are for the first time to identify some of the genes responsible for defining the physiological pathways for the regulation of energy balance. Among the most consequential discoveries were ob gene, leptin and its receptor
8,9)lacking either leptin or its receptors develop morbid obesity weighing about three times of the normal littermates. Although sciences had advanced the understanding of the genetic role in developing obesity, obesity promoting changes in our diet and lack of physical activity are outpacing scientific progress. As it is recommended by Dr. JS Flier in Cell in 2004, a key goal of future research must be to identify mechanisms by which environmental factors interact with specific genes, either to promote, or facilitate resistance to obesity.
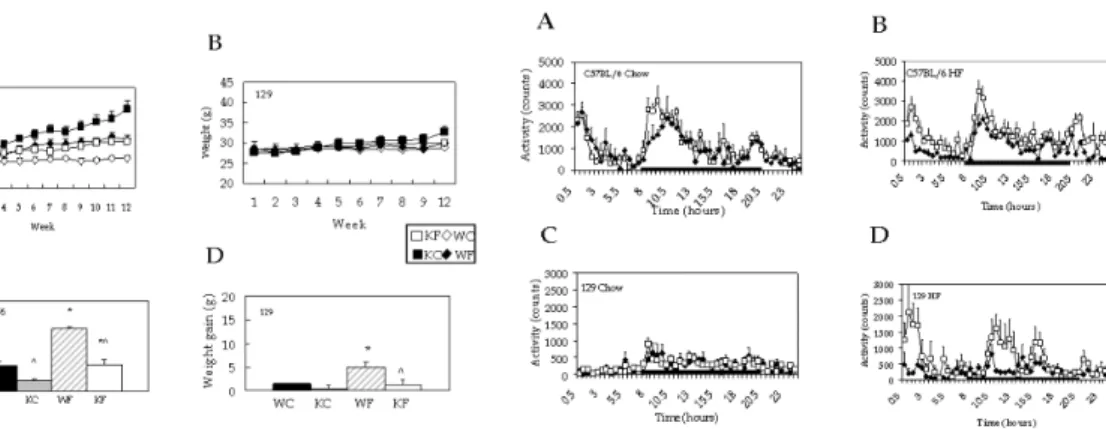
7)In this paper, I will present three different series of studies investigating 1) interaction between genetic background (C57BL/6, obesity prone and 129, obesity resistant) and single gene deletion of melanin concentrating (MCH) hormone in the developing high fat diet induced obesity,
10)2) interaction between high fat diet and exercise in the development of obesity and insulin resistance,
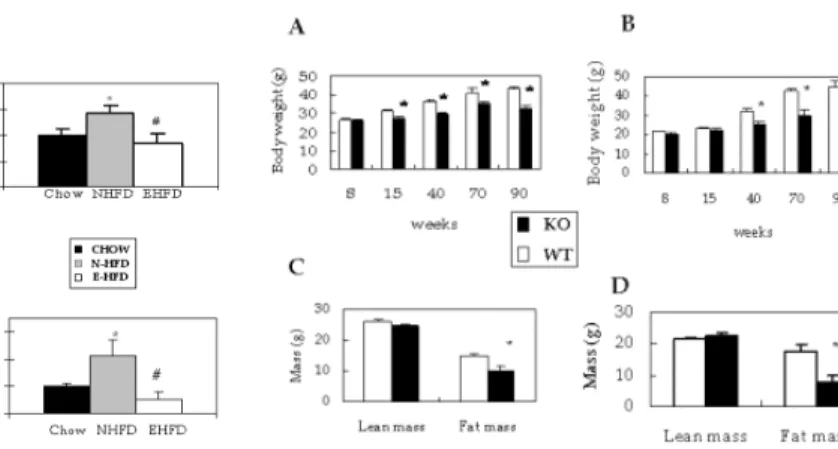
12)3) role of leanness in MCH ablated mice on aging associated physiological variables such as insulin resistance, insulin secretion, locomotor activity, and tumor suppressor, p53.
13)연세대학교 의과대학 사회체육학과