47(5), 503~511(2018)
https://doi.org/10.3746/jkfn.2018.47.5.503쿠마린 유도체 보충급여가 난소절제 마우스의 내장지방 및 지질대사에 미치는 영향
함주리․이미경
순천대학교 식품과학부 식품영양학전공
Effects of Coumarin Derivatives on Visceral Fat Mass and Lipid Metabolism in Ovariectomized Mice
Ju Ri Ham and Mi-Kyung Lee
Department of Food and Nutrition, Sunchon National University
ABSTRACT This study investigated the effects of coumarin derivatives (herniarin, osthole and bergapten) on body fat mass and lipid metabolism in ovariectomized (OVX) mice. Female C3H/HeN mice were divided into five groups:
negative control group (Sham), OVX and OVX mice were fed with a diet containing 0.02% (w/w) herniarin, osthole or bergapten for 6 weeks. Visceral fat (cervical, abdominal, and mesenteric white adipose tissue) mass increased in the OVX group compared with the Sham group; however, herniarin supplementation significantly reduced visceral fat mass. The size of cervical adipocytes decreased in all coumarin supplemented groups relative to the OVX group.
Furthermore, three coumarin derivatives significantly down-regulated expression of lipogenic genes such as fatty acid synthase, stearoyl-coenzyme A desaturase-1 and diacylglycerol O-acyltransferase 2 in adipose tissue. Osthole and ber- gapten supplementation significantly decreased hepatic lipid (triglyceride and free fatty acid) contents compared with the OVX group. Osthole supplementation significantly decreased fatty acid synthase and phosphatidate phosphohydrolase activity compared with the OVX group, whereas bergapten significantly increased hepatic fatty acid oxidation activity.
Overall, these results suggest that herniarin, osthole or bergapten may be useful natural agents for the treatment or prevention of postmenopausal adiposity and lipid dysmetabolism.
Key words: adiposity, coumarin derivatives, hepatic lipid, ovariectomy, visceral fat mass
Received 21 February 2018; Accepted 19 April 2018
Corresponding author: Mi-Kyung Lee, Department of Food and Nutrition, Sunchon National University, Suncheon, Jeonnam 57922, Korea
E-mail: [email protected], Phone: +82-61-750-3656
서 론
폐경(menopausal)은 일반적으로 50세 전후의 여성에서 에스트로겐 호르몬 생산 및 분비 저하로 정상적인 월경이 중지되는 현상을 의미하며, 폐경기 여성 비만은 동일 연령 남성에 비해 발병률이 높다(1). 폐경기 여성의 비만 요인은 여러 가지가 관여하지만 에스트로겐 호르몬 감소가 가장 중 요한 요소로 인지되고 있다(2). 폐경기의 에스트로겐 결핍은 지질대사, 당 대사, 인슐린 작용 및 체지방 분포 등에 영향을 미쳐 체중 및 체지방량의 증가를 유도하며(3), 이는 제2형 당뇨병, 고혈압, 고중성지방혈증 등의 질병으로 발전될 수 있어 이를 통틀어 ‘폐경기 대사증후군(postmenopausal metabolic syndrome)’이라고 한다(4). 폐경기 대사증후군 발병을 감소시키기 위해 호르몬, 약물, 운동 및 식사요법이 주로 처방되고 있는데, 특히 호르몬 보충요법(hormone re-
placement therapy, HRT)은 폐경 여성의 심혈관계 질환이 나 골다공증 개선에 효과적인 것으로 알려져 있다(5). 그러나 장기간의 HRT는 유방암, 자궁내막암, 담낭질환, 고혈압 등 의 발병률을 높이고 있어(6) 보다 안전하고 효과적인 치료제 개발이 요구되고 있다. 이와 같이 에스트로겐을 대체할 수 있는 새로운 기능성 천연물질로 식물성 에스트로겐(phy- toestrogen)에 대한 연구가 관심을 모으고 있다.
식물성 에스트로겐은 에스트로겐과 구조가 비슷하여 에 스트로겐 수용체에 결합하여 유사한 효능을 나타내며(7), 대 표적으로 isoflavone, lignan, coumarin 등이 알려져 있다 (8). 쿠마린계 유도체 중 herniarin(7-methoxycoumarin), osthole(7-methoxy-8-isopentenyl coumarin)과 ber- gapten(5-methoxypsoralen)(Fig. 1)은 주로 라벤더(9), 쑥(10), 마할렙버찌(11), 벌사상자(12) 등에 함유되어 있다.
Herniarin, osthole 및 bergapten은 조골세포의 분화 및 활 성을 증가시켜 골다공증을 개선하는 효과가 보고되었으며 (8), 고지방을 급여한 흰쥐의 고지질혈증과 지방간을 개선하 는 것으로 보고되어 있다(13). 이외에도 이들 쿠마린 유도체 들의 항염증(14) 및 항암(15) 작용이 알려져 있으나, 폐경기 동물모델의 내장지방 감소 및 관련 기전 연구는 이루어져
Herniarin Osthole Bergapten Estradiol
Fig. 1. Structures of coumarin derivatives and estradiol.
Table 1. Composition of experimental diets
Ingredients (g/kg) Sham OVX OVX+H OVX +O OVX
+B Casein
D,L-Methionine Corn starch Sucrose Cellulose Corn oil Mineral mix1) Vitamin mix2) Choline bitartrate Herniarin Osthole Bergapten
200 3 500 150 50 50 35 10 2
-
-
- 200
3 500 150 50 50 35 10 2
-
-
- 200
3 499.8
150 50 50 35 10 2 0.2
-
- 200
3 499.8
150 50 50 35 10 2
- 0.2
- 200
3 499.8
150 50 50 35 10 2
-
- 0.2
1)AIN-76 mineral mixture. 2)AIN-76 vitamin mixture.
있지 않다. 폐경기 동물모델은 주로 난소를 절제하여 에스트 로겐을 결핍시켜 유도하므로(16), 본 연구에서도 난소를 절 제한 마우스에게 herniarin, osthole과 bergapten을 각각 6주간 급여한 후 체중변화, 내장지방량과 지방세포 크기 및 지질대사 관련 유전자 발현, 혈청과 간조직 내의 지질 함량 및 지질대사 효소활성도를 비교・분석하여 폐경기 대사증후 군 개선 기능성 식품소재로 쿠마린계 유도체의 활용 가능성 을 평가하고자 하였다.
재료 및 방법
실험동물 사육
실험동물은 6주령의 암컷 C3H/HeN 마우스 40마리를 오 리엔트바이오(Seongnam, Korea)로부터 구입하였다. 마우 스는 1주간 고형식이로 적응시킨 후 난괴법에 의하여 군당 8마리씩 비난소절제군(Sham), 난소절제군(OVX), 난소절제 +herniarin 급여군(OVX+H), 난소절제+osthole 급여군 (OVX+O)과 난소절제+bergapten 급여군(OVX+B)으로 나누었다. 모든 마우스는 졸레틸-럼푼으로 마취하여 비난 소절제군은 개복 후 난소를 절제하지 않고 봉합하였으며, 난소절제군은 난소를 절제하고 1주일간의 회복기를 거친 후 6주간 사육하였다. 본 연구의 사육기간은 난소절제 후 지질 함량변화를 제시한 선행연구(17,18)를 참고하여 결정되었 다. 동물사육실의 환경은 항온(22±2°C), 항습(55±5%), 12 시간 간격(08:00~20:00)의 광주기로 일정한 조건을 유지하 고 동물들은 폴리카보네이트 케이지에 한 마리씩 분리하여 사육하였다. 동물실험은 순천대학교 동물실험윤리위원회의 승인을 받아 수행하였다(승인번호: SCNU IACUC-2015-01).
본 실험에 사용한 기본식이는 AIN-76의 식이조성에 준 하였으며(Table 1), herniarin(>98%, Sigma-Aldrich, Darm- stadt, Germany), osthole 및 bergapten(>98%, TCI Co., Ltd., Tokyo, Japan) 급여량은 선행연구(19,20,21)를 바탕 으로 식이에 0.02%(w/w) 수준으로 혼합하여 급여하였다.
식이와 식수는 자유롭게 섭취(
ad libitum
)하도록 하였다.혈청 및 장기 채취
사육이 끝난 실험동물은 희생 전 12시간 동안 절식시킨 후 에테르를 흡입시켜 마취시킨 다음 복부 하대정맥(inferior
vena cava)으로부터 공복혈액을 채취해 900×
g
에서 15분 간 원심분리 하여 혈청을 분리하였다. 실험동물의 자궁, 간, 내장지방조직(자궁주변, 후복막, 신장주변, 장간막)은 혈액 채취 후 즉시 적출하여 생리식염수로 수차례 헹군 다음 표면 의 수분을 제거한 후 액체질소로 급냉시켜 -70°C에 보관하 였다.혈청 중의 에스트라디올 함량 측정
혈청 중의 에스트라디올 농도는 효소면역측정법(enzyme- linked immunosorbent assay, ELISA)으로 마우스 ELISA kit(Cusabio Biotech Co., Ltd., Wuhan, China)을 사용하여 측정하였다.
혈청과 간조직의 지질 함량 측정
혈청 중의 중성지질과 총 콜레스테롤 함량은 조제된 kit (Asan, Gyeonggi, Korea)을 사용하여 측정하였다. 간조직 의 지질 함량은 Folch 등(22)의 방법에 준하여 클로로포름 : 메탄올(2:1, v/v) 혼합액으로 지질을 추출한 후 중성지질과 콜레스테롤 함량은 혈청과 동일한 kit으로 측정하였으며, 유 리지방산은 측정용 kit(Shinyang Diagnostics, Seoul, Ko- rea)으로 측정하였다.
지방조직의 형태학적 분석
자궁주변 지방조직 일부를 10% 포름알데히드 용액에 24
Table 2. Primer sequences for RT-qPCR
Gene Full name Forward/Reverse (5'-3')
Actb Cpt1b
Cpt2 Dgat2
Fasn Lipe Scd1
β-actin
carnitine palmitoyltransferase 1b carnitine palmitoyltransferase 2 diacylglycerol O-acyltransferase 2 fatty acid synthase
lipase, hormone sensitive stearoyl-CoA desaturase-1
GAGGTATCCTGACCCTGAAGTA/GCTCGAAGTCTAGAGCAACATAG TGGCTACGGGGTCTCTTACA/AAGTTCGGCGATGTCCAACA GCCTGCTGTTGCGTGACTG/TGGTGGGTACGATGCTGTGC
CTGGCTGATAGCTGCTCTCTACTTC/TGTGATCTCCTGCCACCTTTC CGCTCCTCGCTTGTCGTCTG/AGCCTTCCATCTCCTGTCATCATC GTGAATGAGATGGCGAGGGTC/TGAGGAGTCGCGTTAGAGTC TTCTTCATCGACTGCATGGC/ACTCAGAAGCCCAAAGCTCAG
시간 고정한 다음 수세하고 60% 에탄올에서부터 상승 농도 로 탈수하여 파라핀에 포맷하고, 이것을 4 μm 두께로 박절 하여 hematoxylin-eosin(H-E) 염색한 다음 광학현미경에 서 200배 배율로 관찰하였다.
지방조직의 RNA 분리 및 real-time qPCR 분석
Total RNA 추출은 Trizol 시약(Invitrogen Life Tech- nologies, Grand Island, NY, USA)으로 추출하여 정제하였 으며, 분리한 RNA는 Nano drop 1000(Thermo Fisher Scientific, Waltham, MA, USA)을 이용하여 260 nm와 280 nm에서 흡광도를 측정하여 정량하였다. 그 후 mRNA 발현 은 SYBR green PCR kit(Qiagen, Hilden, Germany)을 사 용하여 real-time PCR analysis를 수행하였다. Real-time PCR은 CFX96TM real-time PCR system(Bio-Rad, Her- cules, CA, USA)을 이용하여 수행하였으며, threshold cy- cle(Ct) 값은 형광커브와 역치선이 만나는 cycle 값으로 CFX3.1(Bio-Rad)로 분석하였다. 각각의 유전자 발현은 동 일한 시료의Actb
발현을 측정하여 보정하였으며, 2-ΔΔCt 방 법을 사용하여 계산되었다(23). Primer 염기서열은 Table 2와 같다.간조직의 효소원 분리 및 지질대사 효소 활성도 측정
간조직의 효소원 분리는 Hulcher와 Oleson(24)의 방법 을 수정・보완하여 사용하였다. 간조직 0.5 g을 취해 잘게 자른 후, 0.1 M triethanolamine, 0.02 M EDTA(ethyl- enediamine tetracetate, pH 7.4), 그리고 2 mM DTT(di- thiothreitol, pH 7.0)를 포함한 완충용액을 이용하여 균질 화한 다음 600×g
(4°C)에서 10분간 원심분리 하여 상층액 을 취한 후 다시 10,000×g
(4°C)에서 20분간 재원심분리 하여 얻은 침전물을 완충용액에 녹여 미토콘드리아를 얻었 다. 분리된 상층액을 100,000×g
(4°C)에서 1시간 동안 초원 심분리 하여 상층액으로 시토졸 분획을 얻었고 침전물은 완 충용액에 녹여 마이크로좀으로 사용하였다. 각 효소원의 단 백질 함량은 bovine serum albumin을 표준으로 하는 Brad- ford(25)의 방법을 사용하여 측정하였다.지질대사 효소 활성도는 Ham 등(26)의 연구에 준하여 측정하였다. Fatty acid synthase(FAS)는 세포질 분획에서 산화되는 NADPH 정도를 340 nm에서 측정하였고, phos- phatidate phosphohydrolase(PAP)는 마이크로좀 분획에
서 생성되는 phosphomolybdate를 820 nm에서 측정하였 다. Fatty acid β-oxidation(β-oxidation)은 NAD+가 NADH 로 환원되는 정도를 340 nm에서 측정하였으며, carnitine palmitoyl-CoA transferase(CPT)는 carnitine이 palmi- toyl-CoA를 기질로 생성된 총 CoASH 생성률을 412 nm에 서 측정하였다.
통계처리
실험결과는 SPSS package(version 22, SPSS Inc., Chi- cago, IL, USA) 프로그램을 이용하여 실험군당 평균±표준 오차로 표시하였고 각 군 간의 평균치의 통계적 유의성 검정 은 one-way ANOVA를 실시하고 실험군 간의 차이는
P
<0.05 수준에서 Duncan’s multiple range test로 사후 검정 하였다.
결과 및 고찰
체중, 자궁무게 및 혈청 에스트라디올 함량에 미치는 영향
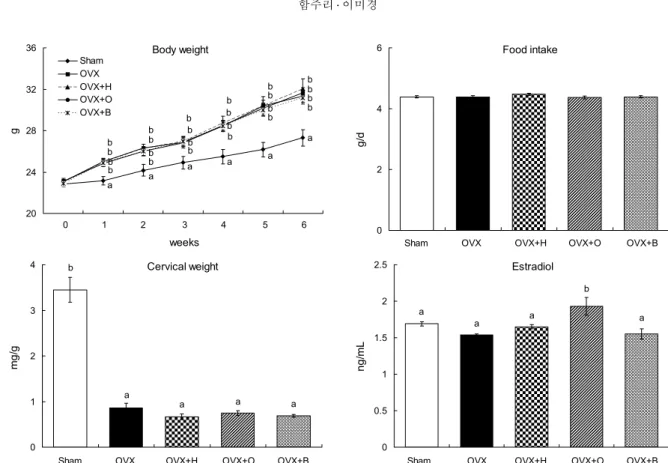
여성의 경우 폐경으로 인한 에스트로겐 호르몬 저하는 에 너지 저장을 증가시켜 비만의 원인이라고 알려져 있는데 (27), 본 연구에서도 난소를 절제한 폐경기 마우스모델에서 유의적인 체중증가(26.25%)를 확인하였다(Fig. 2). 이는 난 소절제를 시행한 1주 후부터 Sham군에 비해 체중이 증가한 선행연구 결과와 일치하였다(28). 난소절제에 의한 체중증 가는 식이섭취량 증가가 주요한 원인으로 작용하지만(29), 본 연구에서는 난소절제로 인한 유의적인 식이섭취량 증가 는 관찰되지 않았다. 또한, 쿠마린 유도체 보충 급여는 난소 절제 마우스의 체중과 식이섭취량에는 유의적인 영향을 미 치지 않았다(Fig. 2). 난소절제는 자궁무게 감소를 유발한다 고 보고되어 있는데(30), 본 실험에서도 OVX군의 단위 체 중당 자궁무게가 Sham군에 비해 크게 감소하였다. 그러나 herniarin, osthole 및 bergapten 급여에 따른 자궁무게 변 화는 없었다(Fig. 2).여성 호르몬의 생성에 중요한 역할을 하는 난소를 적출하 게 되면 에스트라디올의 함량이 감소하여 폐경기 증상이 유 발된다(31). 본 연구에서는 OVX군의 혈청 에스트라디올 함 량이 Sham군에 비해 약 9% 정도 감소하였으나 통계적 유의 성은 없었다. Das 등(32)의 보고에 의하면 홍차에 함유된 폴리페놀 및 식물성 에스트로겐 성분이 난소절제 흰쥐의 에
Body weight
a a a
a a a
b b b b b b
b b b b b b
b
b b b b b
b b b b b b
20 24 28 32 36
0
weeks
g .
Sham OVX OVX+H OVX+O OVX+B
0 1 2 3 4 5 6
Food intake
0 2 4 6
0
g/d .
Sham OVX OVX+H OVX+O OVX+B Cervical weight
a a a a
b
0 1 2 3 4
0
mg/g .
Sham OVX OVX+H OVX+O OVX+B
Estradiol
a b
a a a
0 0.5 1 1.5 2 2.5
0
ng/mL .
Sham OVX OVX+H OVX+O OVX+B
Fig. 2. Effects of herniarin, osthole, and bergapten on body weight, food intake, cervical weight, and serum estradiol levels in
ovariectomized mice. Values are expressed as the mean±SE (n=8). The means not sharing a common letter (a,b) are significantly different among groups (P<0.05).스트로겐 수용체와 결합해 혈청의 에스트라디올 농도를 증 가하였으나, 본 연구에서는 osthole 보충군만 OVX군에 비 해 유의적으로 에스트라디올 함량이 증가하였을 뿐 her- niarin과 bergapten 급여군은 변화가 없었다(Fig. 2). 이는 쿠마린 유도체 보충에 따른 에스트라디올 함량변화가 체중 감소에 의존적이지 않음을 시사한다.
내장지방량, 지방세포 크기 및 지질대사 관련 유전자 발현에 미치는 영향
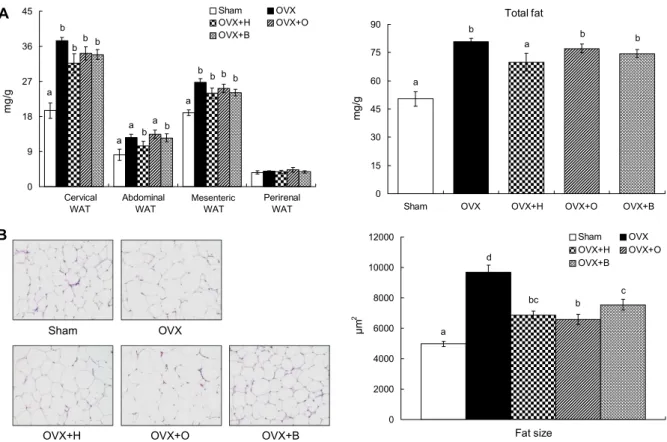
체중 단위당 내장지방량을 측정한 결과 OVX군의 자궁주 변지방, 후복막지방 및 장간막지방 무게는 Sham군에 비해 각각 47.85%, 34.97%, 29.08%의 유의적 증가를 보여 총 내장지방 무게는 난소절제로 인해 37.63% 증가하였다(Fig.
3A). 그러나 OVX+H군의 자궁주변지방과 복부지방량이 OVX군에 비해 15.39%와 17.68%의 유의적인 감소를 보인 한편, OVX+O군과 OVX+B군의 자궁주변 지방량은 감소하 는 경향이었다. 지방조직량의 증가는 지방세포의 크기가 비 대해지는 과정과 지방세포의 수가 증가되는 과정을 통해 발 생한다(33). 따라서 자궁주변 지방조직의 형태학적 변화를 살펴보면 OVX군의 지방세포 크기는 Sham군에 비해 48.67
% 비대해졌으나, 쿠마린 유도체 급여군 모두 지방세포 크기 가 OVX군에 비해 일정하고 작았다(Fig. 3B). 이와 같이 세 가지 쿠마린 유도체가 체지방 감소 효과와 달리 모두 유사하 게 난소절제로 인해 비대해진 지방세포 크기가 감소하였다.
대표적인 식물성 에스트로겐이라고 알려져 있는 제니스테 인을 난소절제한 마우스에게 급여한 연구(34)와 유사하게 지방세포 크기가 감소하였다. 특히 herniarin의 경우 내장지 방 무게 및 지방세포 크기가 동시에 감소함에 따라 폐경기 후 체지방 축적 감소에 가장 효과적일 것으로 사료된다.
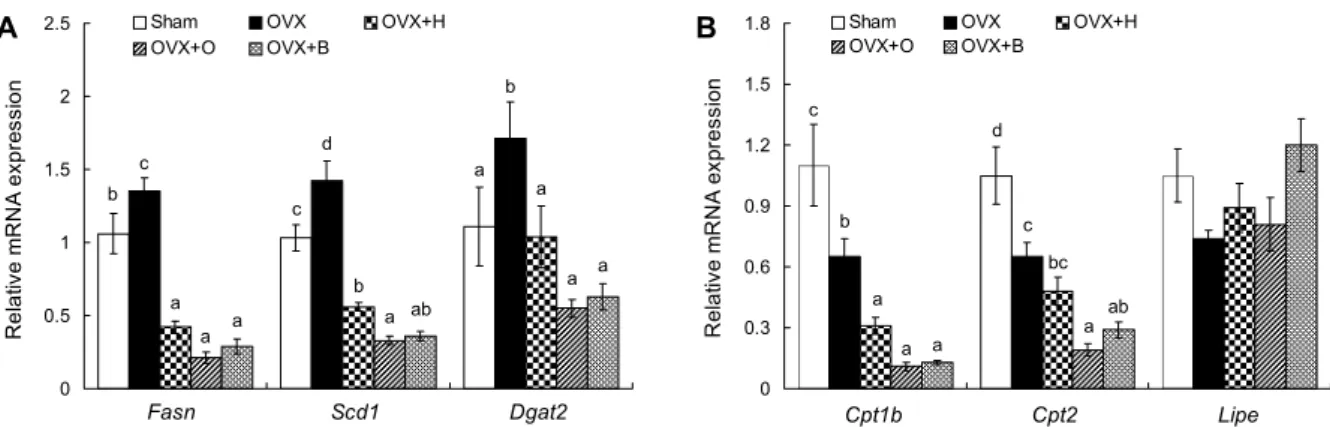
본 실험에서 난소절제 마우스의 체지방량 감소 효과가 에 스트라디올 함량에 의존적이지 않은 것으로 보여, 쿠마린계 유도체의 내장지방 또는 지방세포 크기 감소 효과에 대한 메커니즘을 밝히기 위해 지방조직에서 지질합성 및 산화 관 련 유전자 변화를 측정하였다. 본 연구결과 OVX군의 지방 합성 유전자
Fasn
,Scd1
과Dgat2
가 Sham군보다 유의적으 로 증가하였으며(Fig. 4A), 이들 유전자 간에는 양의 상관관 계가 관찰되었다(Fasn
과Scd1
, r=0.891,P
<0.001,Fasn
과Dgat2
, r=0.669,P
<0.001,Scd1
과Dgat2
는 r=0.569,P
<0.001)(data not shown).
Fasn
은 지방산 합성 유전자인 반 면, 주로 지방조직과 간조직에서 발현되는Scd1
은 단일 불 포화지방산을 생합성 하여 인지질, 중성지질 등과 같이 여러 형태의 지방합성에 기질을 제공한다(35). 따라서Scd1
의 발 현 억제는 단일 불포화지방산의 생성을 감소시켜 지질감소 에 효과적인 것으로 알려져 있다(35). 또한,Dgat
는 중성지 질 합성의 마지막 단계를 촉진하는 인자로(36)Fasn
과Scd1
의 발현이 감소하게 되면Dgat
또한 감소하게 되어 중 성지질 감소에 효과적이다(37). Bryzgalova 등(38)은 고지 방식이를 급여한 난소절제 마우스에게 에스트라디올을 처a a
b
a b
b
b b
b
a b
b
b b
0 9 18 27 36 45
0
mg/g .
Sham OVX
OVX+H OVX+O
OVX+B
a
A
Cervical Abdominal Mesentric Perirenal WAT WAT WAT WAT
Mesenteric
Total fat b b a
b
a
0 15 30 45 60 75 90
0
mg/g .
Sham OVX OVX+H OVX+O OVX+B
B
Sham OVX
OVX+H OVX+O OVX+B
a d
bc b
c
0 2000 4000 6000 8000 10000 12000
0 μm2 .
Sham OVX
OVX+H OVX+O
OVX+B
Fat size
Fig. 3. Effects of herniarin, osthole and bergapten on visceral adipose tissue weight (A) and size of cervical white adipose tissue
(B) in ovariectomized mice. Values are expressed as the mean±SE (n=8). The means not sharing a common letter (a-d) are significantly different among groups (P<0.05). WAT: white adipose tissue, Total fat: sum of cervical WAT, abdominal WAT, mesenteric WAT, and perirenal WAT.리한 결과 지방세포의
Fasn
과Scd1
감소를 통하여 지방축 적 및 지방세포 크기가 감소한다고 보고하였다. 본 연구에서 herniarin 보충군만 내장지방량이 유의적으로 감소한 양상 과 달리 모든 쿠마린계 유도체는 지방조직의 지질합성 관련 유전자 발현을 억제하였다. 이처럼 osthole과 bergapten 급 여 시 지방조직의 지질합성 관련 유전자들의 발현은 감소하 였으나 내장지방량이 유의적으로 감소하지 않은 것은 유전 자 수준에서의 변화는 진행되지만 표현형의 변화를 끌어내 는 데 물질 간의 차이가 있는 것으로 사료되며, 그 이유는 향후 더 연구되어야 할 것이다.한편,
Lipe
유전자는 지방조직에서 저장된 중성지질의 수 화를 촉진하고 자궁 등 여러 조직에서 콜레스테롤 에스터화 를 촉진하는 다양한 기능을 한다(39).Lipe
활성화는 지질의 산화를 조절하는 반면에 결핍되면 지방세포의 수나 크기를 증가시키는데(40), 본 연구결과 난소절제 또는 쿠마린계 유 도체 보충에 따른 유의적인 변화는 없었다(Fig. 4B). 또한, long-chain acyl-carnitine의 생성을 증가시켜 미토콘드리 아에서 β-oxidation을 촉진하는(41)Cpt1b
와Cpt2
유전자 발현은 OVX군이 Sham군보다 유의적으로 억제되었으며, 이들 유전자는 세 가지 쿠마린 유도체 보충으로 발현이 더욱 억제되었다(Fig. 4B). 이는 쿠마린계 유도체 급여 시 지방산 합성이 Sham군보다도 크게 감소한 결과라고 생각한다. 이처럼 herniarin, osthole 및 bergapten은 난소절제 동물의 지방조직 내 지방산 및 중성지질 합성을 억제함으로써 내장 지방량 또는 지방세포 크기 축소에 기여할 것으로 생각한다.
간조직의 지질 함량 및 지질대사 관련 효소 활성도에 미치는 영향
Graffagnino 등(42)은 난소절제로 에스트로겐 분비가 저 하되면 에너지와 지질대사 간의 불균형이 초래되고 지단백 질과 결합한 중성지질이 가수분해 되면서 폐경기에 혈중지 질 농도가 증가한다고 하였다. 따라서 본 실험에서 혈청의 지질 농도를 측정한 결과, 중성지질 함량은 실험군 간에 유 의적인 차이를 보이지 않았으나 OVX군의 총콜레스테롤 함 량은 Sham군보다 유의적으로 증가하였으며, 쿠마린 유도체 에 의한 영향은 관찰되지 않았다(Table 3).
체내 지질 농도가 높아지게 되면 지방산 대사 관련 효소에 의해 영향을 받고 조직에 지방이 축적되게 된다(43). 따라서 간조직의 지질 함량 및 지질대사 관련 효소 활성도를 측정하 였다. 간조직의 중성지질은 OVX군이 Sham군보다 유의적 으로 증가하였으나(Fig. 5A), OVX+O군과 OVX+B군에서 OVX군에 비해 각각 22.98%와 55.39% 감소하였다. 유리지 방산 또한 OVX+O군과 OVX+B군이 OVX군에 비해 유의적 으로 감소하였다(Fig. 5A). Herniarin은 내장지방량 감소에
a b c
b
d c
a
a b
a a a
a a ab
0 0.5 1 1.5 2 2.5
0
Relative mRNA expression .
Sham OVX OVX+H
OVX+O OVX+B
Fasn Scd1 Dgat2
A
d c
b c
bc a
a a
ab a
0 0.3 0.6 0.9 1.2 1.5 1.8
0
Relative mRNA expression .
Sham OVX OVX+H
OVX+O OVX+B
Cpt1b Cpt2 Lipe
B
Fig. 4. Effects of herniarin, osthole, and bergapten on cervical white adipose tissue lipid synthesis (A) and oxidation (B) gene
expression in ovariectomized mice. Values are expressed as the mean±SE (n=8). The means not sharing a common letter (a-d) are significantly different among groups (P<0.05). Fasn: fatty acid synthesis, Scd1: stearoyl-CoA desaturase-1, Dgat2: diacylglycerol O-acyltransferase 2, Cpt1b: carnitine palmitoyltransferase 1b, Cpt2: carnitine palmitoyltransferase 2, Lipe: lipase, hormone sensitive.Table 3. Effects of herniarin, osthole, and bergapten on serum lipid contents in ovariectomized mice
Sham OVX OVX+H OVX+O OVX+B
TG3) (mg/dL) TC (mg/dL)
126.11±11.161) 128.07±7.37a2)
107.90±11.52 184.45±2.74b
118.78±12.83 177.74±13.32b
99.19±12.94 188.72±4.54b
120.29±10.27 183.74±5.33b
1)Values are expressed as the mean±SE (n=8).
2)The means in the row not sharing a common letter are significantly different among groups (P<0.05).
3)TG: triglyceride, TC: total cholesterol.
Triglyceride
b
c c
b
a
0 4 8 12 16 20 24
0
mg/g .
Sham OVX OVX+H OVX+O OVX+B
A
Cholesterola a b ab
b
0 0.5 1 1.5 2 2.5
0
mg/g .
Sham OVX OVX+H OVX+O OVX+B
Free fatty acid
ab a c bc
d
0 7 14 21 28 35
Sham OVX OVX+H OVX+O OVX+B
mg/g .
FAS c
ab a b ab
0 2 4 6 8 10
Sham OVX OVX+H OVX+O OVX+B
nmol/min/mg protein .
B
PAPab a ab bc c
0 25 50 75 100
Sham OVX OVX+H OVX+O OVX+B
μmol/min/mg protein .
Fig. 5. Effects of herniarin, osthole, and
bergapten on hepatic lipid levels (A) and hepatic lipid synthesis (B) and oxidation (C) enzyme activities in ovariectomized mice. Values are expressed as the mean±SE (n=8). The means not sharing a com- mon letter (a-d) are significantly different among groups (P<0.05). FAS: fatty acid synthase, PAP: phosphatidate phosphohy- drolase, β-oxidation: fatty acid β-oxida- tion, CPT: carnitine palmitoyl-CoA trans- ferase.
β-oxidation
b ab a
a c
0 1 2 3 4 5 6 7
Sham OVX OVX+H OVX+O OVX+B
nmol/min/mg protein .
C
β-Oxidation CPT0 2 4 6 8 10
Sham OVX OVX+H OVX+O OVX+B
pmol/min/mg protein .
는 효과적이었으나 간조직의 지질 함량 개선에는 유의적인 영향을 보이지 않았다(Fig. 5A). 난소절제 마우스를 대상으 로 간조직의 지질저하 효과를 보고한 선행연구 중 쿠마린계 물질에 대한 결과가 없어서 직접적인 비교는 할 수 없으나, 난소절제 쥐에게 녹차분말 첨가식이(1%)를 6주간 먹인 결 과 간조직의 콜레스테롤 및 중성지방 함량이 유의적으로 감 소하였다는(44) 결과와 유사하였다.
에스트로겐 호르몬의 결핍은 대사적인 이상을 초래하는 데(45), 특히 지방산 합성 및 중성지질 합성 관련 효소인 FAS와 PAP의 활성이 증가하여 지방산의 길이가 연장되고 결과적으로 간조직 내 지질 함량이 증가하게 된다(21). 본 연구에서 FAS와 PAP를 측정한 결과 Sham군과 OVX군 간 의 차이는 없었으나, OVX군에 비해 OVX+O군의 FAS는 23.07%, PAP는 43.82%의 활성이 감소하였다. 그러나 중 성지방 함량이 가장 낮은 OVX+B군의 FAS 효소의 활성은 OVX군에 비해 유의적으로 높았다(Fig. 5B). 한편, Stark와 Holub(46)에 의하면 난소절제 흰쥐에서 에스트로겐 대체 물질 급여로 β-oxidation 활성이 증가하였다. 따라서 지질 산화에 관련되는 효소인 β-oxidation과 CPT를 측정한 결 과, 난소절제군의 지방산 β-oxidation 활성이 Sham군보다 유의적으로 낮았다. 그러나 OVX+B군의 β-oxidation 활성 이 OVX군에 비해 38.28% 유의적으로 증가하였다(Fig. 5C).
이처럼 osthole의 경우 지방산 산화보다 지방산 및 중성지 질 합성 효소를 유의적으로 억제하였으며, bergapten은 지 방산 합성보다는 지방산 산화를 촉진함으로써 간조직의 지 질 함량을 감소하는 것으로 나타났다. 이와 같이 쿠마린계 유도체들의 난소절제 마우스의 간조직 지질 함량 감소는 물 질에 따라 다른 기전으로 이루어지는 것으로 사료된다.
요 약
본 연구는 폐경 이후 여성의 내장지방 및 지질대사에 미치는 쿠마린계 유도체(herniarin, osthole, bergapten)의 영향을 살펴보기 위하여, 난소를 절제한 마우스에게 herniarin, osthole 및 bergapten을 각각 식이에 0.02% 수준으로 조제 하여 6주간 급여하였다. Herniarin, osthole 및 bergapten 보충으로 인한 난소절제 마우스의 체중과 식이섭취량에는 영향을 미치지 않았다. 내장지방량은 혈청의 에스트라디올 함량에 비의존적으로 herniarin 급여 시 자궁주변 및 복부지 방 무게가 유의적으로 감소하여 총 내장지방량이 OVX군에 비해 OVX+H군에서 13.48% 감소하였다. 한편 자궁주변 지 방조직의 지방세포 크기는 세 가지 쿠마린 유도체 급여군 모두에서 뚜렷이 감소하였다. 자궁주변 지방조직의 지방 합 성에 관여하는
Fasn
,Scd1
과Dgat2
의 발현이 Sham군에 비해 OVX군에서 증가하였으나 herniarin, osthole 및 ber- gapten 모두 지방 합성 관련 유전자 및 지방산 산화에 관련 되는Cpt1b
와Cpt2
는 유전자 발현이 낮았다. 혈청의 중성지 질과 총 콜레스테롤은 실험군 간의 유의적인 차이를 보이지않았으나, 난소절제로 인하여 간조직의 중성지질 함량은 증 가하였고 중성지질, 콜레스테롤 및 유리지방산 함량은 OVX 군에 비해 OVX+O와 OVX+B군에서 유의적으로 감소하였 다. Herniarin은 내장지방량 감소에는 효과적이었으나 간조 직의 지질 함량 개선에는 유의적이지 않았다. 지질 합성에 관련되는 효소인 FAS와 PAP의 경우 OVX군에 비해 OVX+
O군에서 유의적으로 감소하였으며, 지방산 산화 효소인 β -oxidation의 경우는 OVX+B군이 OVX군에 비해 활성이 증가하였다. 이처럼 쿠마린 유도체의 급여는 난소절제로 인 한 내장지방 축적 또는 지질대사를 개선하여 폐경기 이후 비만 예방에 유용한 천연자원 물질로 활용 가능성이 있는 것으로 평가된다.
감사의 글
이 성과는 2015년도 정부(과학기술정보통신부)의 재원으로 한국연구재단의 지원을 받아 수행된 연구임(No. NRF-2015 R1C1A2A01052146).
REFERENCES
1. BaHammam AS, Pandi-Perumal SR, Piper A, Bahammam SA, Almeneessier AS, Olaish AH, Javaheri S. 2016. Gender differences in patients with obesity hypoventilation syn- drome. J Sleep Res 25: 445-453.
2. D’Eon TM, Souza SC, Aronovitz M, Obin MS, Fried SK, Greenberg AS. 2005. Estrogen regulation of adiposity and fuel partitioning. Evidence of genomic and non-genomic reg- ulation of lipogenic and oxidative pathways. J Biol Chem 280: 35983-35991.
3. Pasquali R, Casimirri F, Labate AM, Tortelli O, Pascal G, Anconetani B, Gatto MR, Flamia R, Capelli M, Barbara L.
1994. Body weight, fat distribution and the menopausal sta- tus in women. The VMH Collaborative Group. Int J Obes
Relat Metab Disord 18: 614-621.
4. Spencer CP, Godsland IF, Stevenson JC. 1997. Is there a menopausal metabolic syndrome?. Gynecol Endocrinol 11:
341-355.
5. Belchetz PE. 1994. Hormonal treatment of postmenopausal women. N Engl J Med 330: 1062-1071.
6. Grady D, Herrington D, Bittner V, Blumenthal R, Davidson M, Hlatky M, Hsia J, Hulley S, Herd A, Khan S, Newby LK, Waters D, Vittinghoff E, Wenger N, HERS Research Group. 2002. Cardiovascular disease outcomes during 6.8 years of hormone therapy: Heart and Estrogen/progestin Replacement Study follow-up (HERS Ⅱ). JAMA 288: 49- 57.
7. Kuiper GGJM, Lemmen JG, Carlsson B, Corton JC, Safe SH, Van Der Saag PT, Van Der Burg B, Gustafsson JÅ.
1998. Interaction of estrogenic chemicals and phytoestrogens with estrogen receptor β. Endocrinology 139: 4252-4263.
8. Woo JT, Yonezawa T, Nagai K. 2010. Phytochemicals that stimulate osteoblastic differentiation and bone formation. J
Oral Biosci 52: 15-21.
9. Brown SA. 1962. Biosynthesis of coumarin and herniarin in lavender. Science 137: 977-978.
10. Hofer O, Szabó G, Greger H. 1986. 2-Hydroxy-4-methoxy-
trans-cinnamic acid as a precursor of herniarin in Artemisia
dracunculus. Monatsh Chem 117: 1219-1222.
11. Santamour Jr FS, Riedel LGH. 1994. Distribution and in- heritance of scopolin and herniarin in some Prunus species.
Biochem Syst Ecol 22: 197-201.
12. Zhang L, Yan R, Su R, Yang C, Liu S, Yu X, Chang X, Zhang S, Liu C, Xu M, Zheng W, Chem Y, Wang Q. 2014.
Bioavailability enhancement of osthole after oral admin- istration of Bushen Yizhi prescription extract to rats fol- lowed by Cnidium monnieri (L.) Cusson fruits extract in comparison to pure osthole at different doses. J Ethnophar-
macol 152: 266-271.
13. Zhang Y, Xie ML, Zhu LJ, Gu ZL. 2007. Therapeutic effect of osthole on hyperlipidemic fatty liver in rats. Acta Phar-
macol Sin 28: 398-403.
14. Rezaee R, Behravan E, Behravan J, Soltani F, Naderi Y, Emami B, Iranshahi M. 2014. Antigenotoxic activities of the natural dietary coumarins umbelliferone, herniarin and 7- isopentenyloxy coumarin on human lymphocytes exposed to oxidative stress. Drug Chem Toxicol 37: 144-148.
15. You L, An R, Wang X, Li Y. 2010. Discovery of novel osthole derivatives as potential anti-breast cancer treatment.
Bioorg Med Chem Lett 20: 7426-7428.
16. Preuss HG. 1993. Nutrition and diseases of women: car- diovascular disorders. J Am Coll Nutr 12: 417-425.
17. Paquette A, Shinoda M, Rabasa Lhoret R, Prud’homme D, Lavoie JM. 2007. Time course of liver lipid infiltration in ovariectomized rats: impact of a high-fat diet. Maturitas 58:
182-190.
18. Côté I, Chapados NA, Lavoie JM. 2014. Impaired VLDL assembly: a novel mechanism contributing to hepatic lipid accumulation following ovariectomy and high-fat/high-cho- lesterol diets?. Br J Nutr 112: 1592-1600.
19. Liu WX, Jia FL, He YY, Zhang BX. 2012. Protective effects of 5-methoxypsoralen against acetaminophen-induced hep- atotoxicity in mice. World J Gastroenterol 18: 2197-2202.
20. Zhang J, Xue J, Wang H, Zhang Y, Xie M. 2011. Osthole improves alcohol-induced fatty liver in mice by reduction of hepatic oxidative stress. Phytother Res 25: 638-643.
21. Sim MO, Ham JR, Lee HI, Seo KI, Lee MK. 2014. Long- term supplementation of umbelliferone and 4-methylum- belliferone alleviates high-fat diet induced hypertriglycer- idemia and hyperglycemia in mice. Chem Biol Interact 216:
9-16.
22. Folch J, Lees M, Sloane Stanley GH. 1957. A simple method for the isolation and purification of total lipids from animal tissues. J Biol Chem 226: 497-509.
23. Livak KJ, Schmittgen TD. 2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 25: 402-408.
24. Hulcher FH, Oleson WH. 1973. Simplified spectrophotomet- ric assay for microsomal 3-hydroxy-3-methylglutaryl CoA reductase by measurement of coenzyme A. J Lipid Res 14:
625-631.
25. Bradford MM. 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72: 248-254.
26. Ham JR, Lee HI, Choi RY, Sim MO, Seo KI, Lee MK. 2016.
Anti-steatotic and anti-inflammatory roles of syringic acid in high-fat diet-induced obese mice. Food Funct 7: 689-697.
27. Picard F, Deshaies Y, Lalonde J, Samson P, Labrie C, Bélanger A, Labrie F, Richard D. 2000. Effects of the estro- gen antagonist EM-652.HCl on energy balance and lipid metabolism in ovariectomized rats. Int J Obes Relat Metab
Disord 24: 830-840.
28. Lemieux C, Picard F, Labrie F, Richard D, Deshaies Y. 2003.
The estrogen antagonist EM-652 and dehydroepiandroster- one prevent diet- and ovariectomy-induced obesity. Obes Res 11: 477-490.
29. McElroy JF, Wade GN. 1987. Short- and long-term effects of ovariectomy on food intake, body weight, carcass composition, and brown adipose tissue in rats. Physiol Behav 39: 361-365.
30. Lee YH, Hyun SH, Choung SY. 2006. Effect of singled and mixed pomegranate on postmenopausal symptoms in ovar- iectomized rats. Yakhak Hoeji 50: 177-183.
31. Kim HC, Kum EJ, Kwon DH, Lee H. 2009. The effect of pomegranate extracts on the menopausal syndromes. J Exp
Biomed Sci 15: 217-227.
32. Das AS, Das D, Mukherjee M, Mukherjee S, Mitra C. 2005.
Phytoestrogenic effects of black tea extract (Camellia si-
nensis) in an oophorectomized rat (Rattus norvegicus) mod-
el of osteoporosis. Life Sci 77: 3049-3057.33. Rosen ED, Spiegelman BM. 2000. Molecular regulation of adipogenesis. Annu Rev Cell Dev Biol 16: 145-171.
34. Jeong SH. 2017. Anti-obesity effects of genistein in female ovariectomy-induced obese mice. J Oil Appl Sci 34: 427-435.
35. Miyazaki M, Ntambi JM. 2003. Role of stearoyl-coenzyme A desaturase in lipid metabolism. Prostaglandins Leukot
Essent Fatty Acids 68: 113-121.
36. Postic C, Girard J. 2008. Contribution of de novo fatty acid synthesis to hepatic steatosis and insulin resistance: lessons from genetically engineered mice. J Clin Invest 118: 829-838.
37. Li JX, Ke DZ, Yao L, Wang S, Ma P, Liu L, Zuo GW, Jiang LR, Wang JW. 2017. Response of genes involved in lipid metabolism in rat epididymal white adipose tissue to different fasting conditions after long-term fructose consump- tion. Biochem Biophys Res Commun 484: 336-341.
38. Bryzgalova G, Lundholm L, Portwood N, Gustafsson JÅ, Khan A, Efendic S, Dahlman-Wright K. 2008. Mechanisms of antidiabetogenic and body weight-lowering effects of es- trogen in high-fat diet-fed mice. Am J Physiol Endocrinol
Metab 295: E904-E912.
39. Osuga JI, Ishibashi S, Oka T, Yagyu H, Tozawa R, Fujimoto A, Shionoiri F, Yahagi N, Kraemer FB, Tsutsumi O, Yamada N. 2000. Targeted disruption of hormone-sensitive lipase results in male sterility and adipocyte hypertrophy, but not in obesity. Proc Natl Acad Sci USA 97: 787-792.
40. Wang SP, Laurin N, Himms-Hagen J, Rudnicki MA, Levy E, Robert MF, Pan L, Oligny L, Mitchell GA. 2001. The adipose tissue phenotype of hormone-sensitive lipase defi- ciency in mice. Obes Res 9: 119-128.
41. Zang Y, Wang T, Xie W, Wang-Fischer YL, Getty L, Han J, Corkey BE, Guo W. 2005. Regulation of acetyl CoA car- boxylase and carnitine palmitoyl transferase-1 in rat adipo- cytes. Obes Res 13: 1530-1539.
42. Graffagnino CL, Falko JM, La Londe M, Schaumburg J, Hyek MF, Shaffer LET, Snow R, Caulin-Glaser T. 2006.
Effect of a community-based weight management program on weight loss and cardiovascular disease risk factors. Obesity 14: 280-288.
43. Kang MY, Chung SI, Lee SC. 2016. Effects of Drynariae Rhizoma hot water extract on metabolic improvement in the ovariectomized rat model of menopause. Korean J Med
Crop Sci 24: 370-374.
44. Choi MJ, Jung YJ. 2009. Effects of green tea powder on plasma and liver lipids in ovariectomized rats. J Kor Tea
Soc 15: 93-100.
45. Carr MC. 2003. The emergence of the metabolic syndrome
with menopause. J Clin Endocrinol Metab 88: 2404-2411.
46. Stark KD, Holub BJ. 2004. Differential eicosapentaenoic acid elevations and altered cardiovascular disease risk factor re-
sponses after supplementation with docosahexaenoic acid in postmenopausal women receiving and not receiving hor- mone replacement therapy. Am J Clin Nutr 79: 765-773.