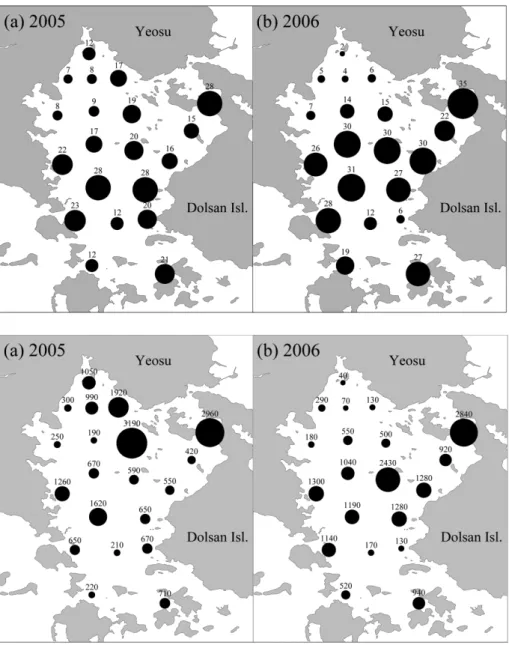
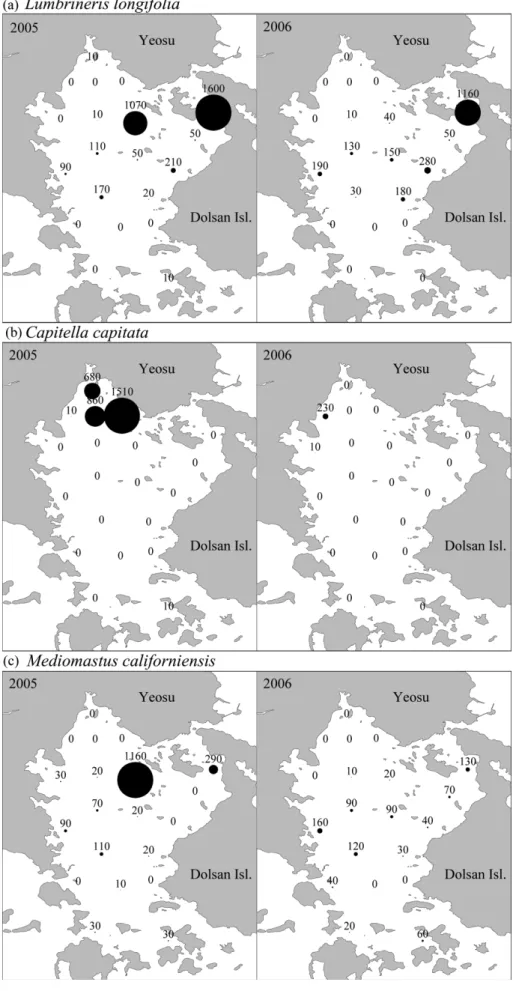
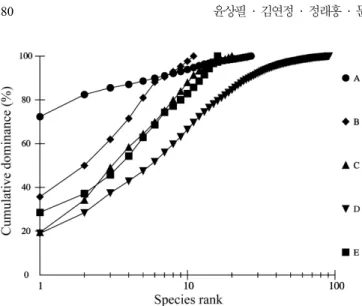
관련 문서
Since the two countries have established diplomatic relations in 1992, the trade between Korea and China has been developing rapidly, By the end of 2006, Korea
The matrix A show the cost per computer (in thousands of dollars) and B the production figures for the year 2005 (in multiples of 1000 units).. Find a matrix C that
Mudeung and Naejang, located on the southwestern part of Korea, were studied in terms of the benthic macroinvertebrate community with selecting eight study stations over
-Compared to Romanesque and Byzantine architecture, Gothic architecture present a clear awareness of the structure itself. Structure in Romanesque
As a affiliated company of Green Cross in the field of genome analysis, we provide disease diagnosis services, such as prenatal genetic test, oncogenomic analysis and
• Started in 2005, the third year’s report on the project, which is currently being edited, focused on the hospitality of local people.. • Studio-L in 2005 and he
The index is calculated with the latest 5-year auction data of 400 selected Classic, Modern, and Contemporary Chinese painting artists from major auction houses..
Key Words: Carbon tax, labor migration, tax regime, welfare effect, fossil fuel demand, computable general equilibrium model, double dividend hypothesis..