Development of Molecular Markers for Alternative Oxidase Synthesis Genes in Brassica rapa L.
Ye Sol Jeong and Sang-Min Chung*
Department of Life Science, Dongguk University-Seoul, Seoul, 100715, Korea Received November 16, 2009 /Accepted November 27, 2009
The low and high temperature stress might affect the yield and quality of many crop species.
Alternative oxidase (AOX) gene is known as factors related to stress resistance in plants. In order to develop molecular markers related to stress resistance in Chinese cabbage, fifteen ESTs sharing se- quence similarity to arabidopsis AOX genes were found usingBrassica rapa EST database from NCBI.
The polymorphic DNA sequences using the ESTs were then screened between Chinese cabbage,
‘Chiifu’ and ‘Kenshin’. We found four ESTs that have either insertion or deletion between the two cultivars. These polymorphic sites were then targeted for development of the four PCR based molec- ular markers. These molecular markers developed in this study could be useful for a test of their rela- tionship with abiotic stress resistance in Chinese cabbage.
Key words : Chinese cabbage, abiotic stress, alternative oxidase, molecular marker
*Corresponding author
*Tel:+82-2-2260-8915, Fax:+82-2-2260-8915
*E-mail : [email protected]
서 론
Brassica속은 씨앗을 이용한 오일, 야채, 및 조미료로 이용 되는 작물 등을 포함한다[8,10]. 그 중 배추, Brassica rapa L.
(2n=20, genome AA)는 김치의 주재료로 국내뿐 아니라 세계 시장에서도 식품영양학적 우수성을 인정받아 그 수요가 증가 하고 있다[9,14]. 배추는 호냉성 채소로 생육적온인 20oC 이상 이 되면 생육이 약해지며 이러한 고온 스트레스는 생산량 및 품질 저하를 유발할 수 있다[5,6,11]. 또한 재배 적온보다 기온 이 낮아지면 동해를 입게 되며 작물의 생산력을 저하시키는 요인이 될 뿐만 아니라[5,6,7] 작물종이 생산되는 지리적 위치 를 제한하는 등 수확량에 영향을 미쳐 경제적 손실을 입게 한다[17].
고온 또는 저온 스트레스가 작용하는 환경에서 활성산소종 의 과다한 축적을 방지해 식물체의 활동을 개선하는 것으로 추측되는 기능성 유전자인 Alternative oxidase (AOX) gene은 식물에 있는 전자전달계의 일부분을 형성하는 효소이다 [13,18]. AOX 유전자의 발현은 저온스트레스, 활성산소종, 그 리고 병원체의 침입 등에 영향을 주는 것으로 알려져 있는데 아마도 산화 스트레스의 감소를 통해 환경 스트레스에 대한 내성 향상을 강화할 것으로 추측된다[12,19]. Alternative oxi- dase는 미토콘드리아내막에 붙어있는 필수 막단백질로 추위 와 같은 스트레스가 작용하는 환경 속에서 활성 산소족의 과 다한 축적을 방지함으로써 식물체의 활동을 개선할 것으로 추측된다[3]. 고베대학의 Shigeo Takumi의 연구논문에 의하
면 AOX는 스트레스를 유발하는 환경에서 모든 조직 내의 과 다한 활성산소족의 형성을 방지하며 대사 작용에도 영향을 미칠 수 있는 것으로 발표하였다[16].
많은 연구진들이 작물 생산성을 증가시키고 작물의 재배 가능 지역을 확장 시키고자 스트레스를 유발하는 저온 또는 고온 환경에서 작물의 저항성을 개선시킨 새로운 품종 개발에 노력하고 있다. 스트레스 저항성 형질을 감수성 품종에 도입 하기 위해서 많은 시간과 비용이 필요하므로 이를 극복하기 위해 고안된 것이 바로 DNA marker를 이용한 선발이다. 이러 한 마커에는 단일염기다형성(single nucleotide polymorphism, SNP) 마커가 많이 사용되고 있다[2,15].
이러한 경향에 맞춰 본 연구는 배추 내서성 육종에 적용이 가능한 스트레스 저항성 관련 분자마커개발을 위해 AOX 유 전자와 관련된 다형성 분자 마커를 개발하였다. 이를 이용하 여 장기적으로 저온 저항성에 직접적으로 연관된 DNA 마커 개발을 하여 환경 스트레스 저항성 배추품종개발에 기초연구 자료로 활용될 수 있다고 생각된다.
재료 및 방법 배추의 AOX 합성 관련 유전자 검색
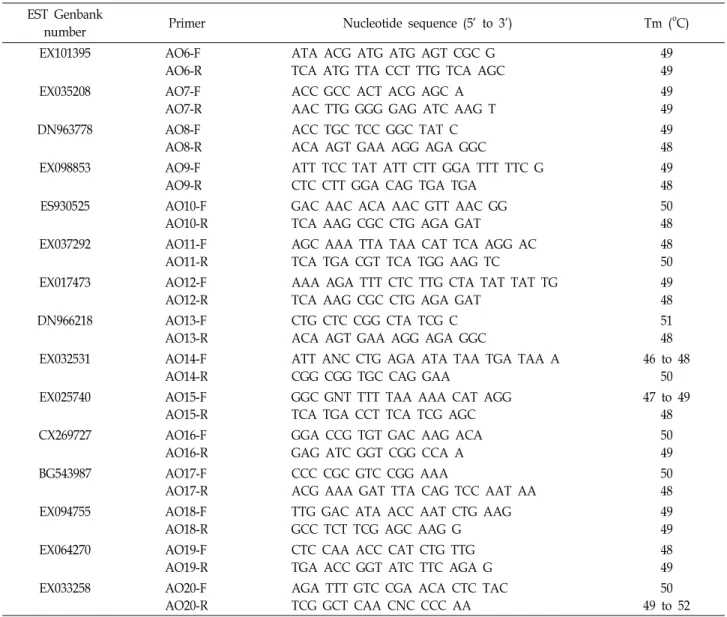
애기장대의 AOX 합성 관련 유전자를 NCBI 데이터베이스 를 이용하여 찾았다[4]. 애기장대에서 찾은 AOX 합성 관련 유전자의 염기서열을 이용하여 EST 데이터베이스를 이용해 검색을 하였으며,Brassica rapa로 제한을 두고, Brassica rapa가 아닌 것은 모두 제거를 하였다. 그 결과 얻은 ESTs를 서로 비교 를 하였으며, 동일염기서열을 보이는 ESTs들은 제거하여 총 15개의 ESTs를 얻을 수 있었다(Table 1). ‘지부(Chiifu)’와 ‘권
Table 1. Fifteen ESTs sharing sequence similarity to alternative oxidase synthesis genes in Brassica rapL. and primers used for PCR amplification
EST Genbank
number Primer Nucleotide sequence (5' to 3') Tm (oC)
EX101395 AO6-F
AO6-R ATA ACG ATG ATG AGT CGC G
TCA ATG TTA CCT TTG TCA AGC 49
49
EX035208 AO7-F
AO7-R ACC GCC ACT ACG AGC A
AAC TTG GGG GAG ATC AAG T 49
49
DN963778 AO8-F
AO8-R ACC TGC TCC GGC TAT C
ACA AGT GAA AGG AGA GGC 49
48
EX098853 AO9-F
AO9-R ATT TCC TAT ATT CTT GGA TTT TTC G
CTC CTT GGA CAG TGA TGA 49
48
ES930525 AO10-F
AO10-R GAC AAC ACA AAC GTT AAC GG
TCA AAG CGC CTG AGA GAT 50
48
EX037292 AO11-F
AO11-R AGC AAA TTA TAA CAT TCA AGG AC
TCA TGA CGT TCA TGG AAG TC 48
50
EX017473 AO12-F
AO12-R AAA AGA TTT CTC TTG CTA TAT TAT TG
TCA AAG CGC CTG AGA GAT 49
48
DN966218 AO13-F
AO13-R CTG CTC CGG CTA TCG C
ACA AGT GAA AGG AGA GGC 51
48
EX032531 AO14-F
AO14-R ATT ANC CTG AGA ATA TAA TGA TAA A
CGG CGG TGC CAG GAA 46 to 48
50
EX025740 AO15-F
AO15-R GGC GNT TTT TAA AAA CAT AGG
TCA TGA CCT TCA TCG AGC 47 to 49
48
CX269727 AO16-F
AO16-R GGA CCG TGT GAC AAG ACA
GAG ATC GGT CGG CCA A 50
49
BG543987 AO17-F
AO17-R CCC CGC GTC CGG AAA
ACG AAA GAT TTA CAG TCC AAT AA 50
48
EX094755 AO18-F
AO18-R TTG GAC ATA ACC AAT CTG AAG
GCC TCT TCG AGC AAG G 49
49
EX064270 AO19-F
AO19-R CTC CAA ACC CAT CTG TTG
TGA ACC GGT ATC TTC AGA G 48
49
EX033258 AO20-F
AO20-R AGA TTT GTC CGA ACA CTC TAC
TCG GCT CAA CNC CCC AA 50
49 to 52
심(Kenshin)’의 염기서열을 비교하기 위해 각 EST에 그 양 끝 염기서열을 이용하여 포워드 프라이머와 리버스 프라이머를 각각 제작하였다(Table 1).
식물 DNA 정제
한국배추게놈소재은행으로부터 분양받은 고온 스트레스에 약한 품종 ‘지부’와 비교적 강하다고 알려진 ‘권심’의 배추 종 자를 발아시켜 키운 후 어린잎을 수확하고 CTAB method를 이용해 DNA를 추출하였다. 곱게 마쇄된 시료에 CTAB buffer 를 750 μl을 가하고 30초 vortexing 하여 추가로 60oC에서 1시 간 배양하였다. 750 μl phenol:chloroform:isomyl alchol (25:
24:1)을 넣고 15초 vortexing, 5분 원심분리 하였다. 얻은 상층 액을 2 ml tube로 옮긴 후 두 번에 걸쳐 chloroform:isomyl alcohol (24:1) 용액으로 세척한 후 EtOH를 900 μl 넣고 -70oC
에서 1시간 보관 후에 4oC에서 10분간 원심분리 하였다. DNA 펠렛을 확인한 후 상층액을 버리고 자연건조를 시켰다. 추출 된 DNA는 증류수 50 μl로 현탁시켰다.
중합효소연쇄반응(PCR)과 gel 전기영동
CTAB method를 이용하여 얻은 ‘지부’와 ‘권심’ DNA를 각 각 디자인한 프라이머를 이용하여 PCR증폭을 하였다. PCR에 사용된 모든 시약은 솔젠트 회사의 e-taq kit를 사용하였다.
증폭에 이용된 DNA는 15 ng이고 total volume은 15 μl로 10 mM dNTP 0.3 μl, 10× buffer는 1.5 μl, taq polymerase는 0.4 unit으로 0.08 μl, 100 μM의 Foward primer, Reverse primer 각각 1 μl, ‘지부’와 ‘권심’의 DNA는 15 ng을 사용하였다. PCR 은 93oC에서 3분 후 93oC에서 30초, 50oC에서 1분, 68oC에서 3분 과정을 40회 반복하고, 72oC 7분 후 종료 또는 5oC에서
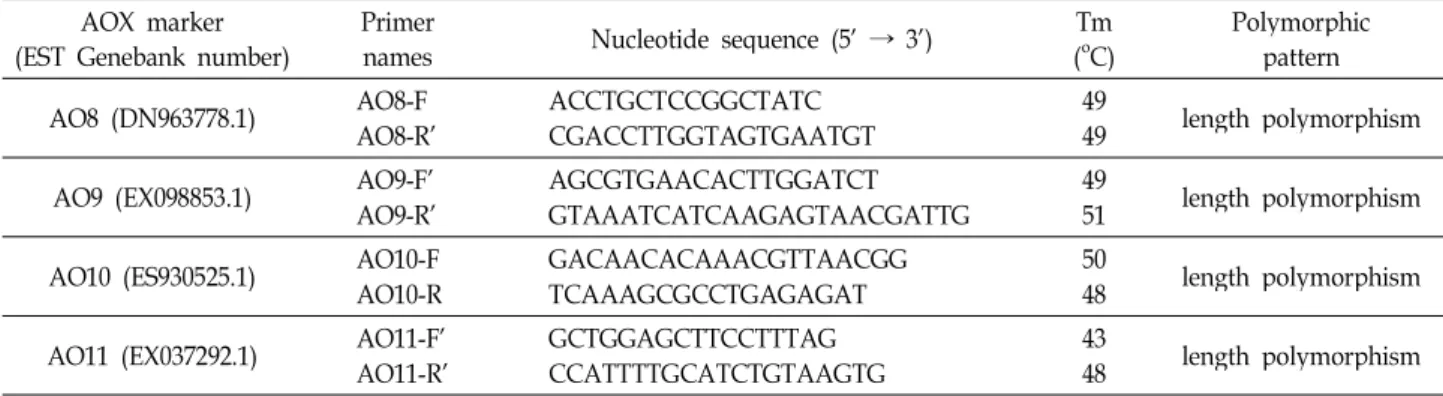
Table 2. Polymorpic DNA markers of ESTs sharing sequence similarity to AOX synthesis genes in Brassica rapa L.
AOX marker
(EST Genebank number) Primer
names Nucleotide sequence (5' → 3') Tm
(oC) Polymorphic pattern AO8 (DN963778.1) AO8-F
AO8-R' ACCTGCTCCGGCTATC
CGACCTTGGTAGTGAATGT 49
49 length polymorphism AO9 (EX098853.1) AO9-F'
AO9-R' AGCGTGAACACTTGGATCT
GTAAATCATCAAGAGTAACGATTG 49
51 length polymorphism AO10 (ES930525.1) AO10-F
AO10-R GACAACACAAACGTTAACGG
TCAAAGCGCCTGAGAGAT 50
48 length polymorphism AO11 (EX037292.1) AO11-F'
AO11-R' GCTGGAGCTTCCTTTAG
CCATTTTGCATCTGTAAGTG 43
48 length polymorphism 보관되었다. 증폭된 DNA는 2% 아가로스 gel에서 전기영동을
이용해 DNA 조각의 크기를 확인하였다.
염기서열분석과 유전자 다형성분석
PCR 증폭을 해 얻은 DNA를 쏠젠트(주)에 염기서열분석을 의뢰하였다. 얻어진 염기서열분석 결과는 staden package 프 로그램(http://staden.sourceforge.net/)을 이용하여 ‘지부’와
‘권심’ 간 염기서열을 비교하였다.
결과 및 고찰
염기 서열 분석을 위해 PCR 증폭을 위해 사용한 프라이머 목록은 Table 1과 같다. PCR 증폭 후 아가로스 gel 상에서 DNA 크기 차이(Fig. 1)를 보이는 EST DNA 증폭 밴드를 확인 하여 사용된 프라이머가 직접 분자 표지로 이용 가능함을 확 인할 수 있었다. 아가로스 gel 상에서 DNA 크기 차이를 확인
←
←
Fig. 1. Agarose gel analysis of codominant AO10 marker. C:
Chiifu, K: Kenshin, S: Size-marker
할 수 없었던 나머지 AOX 관련 유전자 증폭 DNA는 염기서열 분석을 통해 insertion 또는 deletion (In/del) 및 SNP 다형성 유무를 확인하였다. 염기서열 분석은 쏠젠트(주)에 의뢰를 하 였으며 얻어진 염기서열을 이용하여 SNP와 In/del 분석은 staden pakage program을 이용하여 수행했다. 이러한 AOX 합성 관련 유전자 염기서열 비교에서 세 개의 in/del이 추가로 발견되었다(Fig. 2).
In/del이 있는 경우(예: AO8, AO9, AO11) in/del을 대상으 로 가까운 주변 염기서열에서 프라이머를 제작하였다(Table 2). PCR 증폭되는 DNA 크기는 300 bp 이하가 될 수 있도록 프라이머 위치를 제한하여 상대적으로 작은 PCR 증폭 DNA 크기에서 in/del로부터 나타나는 염기서열 크기 차이를 관찰 이 가능하도록 하였다(Figs. 2, 3). AO8과 AO11의 경우 ‘지부’
‘권심’간 5 bp 차이를 보이므로 젤 전기영동 시 200 volt에서 2시간 이상의 충분한 시간으로 이동시켜야 젤 상에서의 크기 차이를 구분할 수 있다. 이러한 결과는 염기서열 분석처럼 높 은 비용과 고가의 장비 없이 다형성을 분석할 수 있으며, DNA 마커들은 유전적 다양성 및 유전자 지도 작성 등에 유용하게 활용될 수 있다.
본 연구에서 개발한Brassica rapa의 AOX 마커의 경우, 각기 다른 AOX유전자이지만 유사한 EST들이 많아 이를 마커로 개발하는데 어려움이 있었다. 다른 AOX ESTs에서 하나의 프 라이머 페어를 사용해서 여러 개의 DNA 밴드가 증폭되는 경 우는 추 후 클로닝을 통한 염기서열 분석을 수행하여 추가로 다형성을 확인하고 분자마커를 개발해야 할 필요성이 있는 것으로 생각된다.
AO8 AO8
Chiifu Kenshin
TGACCAAATC****CTAACCCTAACATTGTC*TGTGTGATTA TGACCAAAACCTAACTAACCCTTGCATTGTCTTGTGTGATTA AO9
AO9 Chiifu Kenshin
ACGAATCATCCACCGG*********AGGTAACAAAGGAATCG ACGAATCATCCACTGGAGGCAACAAGGTGAGCAAAGGTATTG AO11
AO11 Chiifu Kenshin
TAATCACACA*GACAATGTTAGGGTTAG****GATTTGGTCA TAATCACACAAGACAATGCAAGGGTTAGTTAGGTTTTGGTCA
Fig. 2. Nucleotide sequence alignment of AO8, AO9, and AO11 showed insertion or deletion. The bold letters indicate polymorphic sites.
→
→
→→
→
→
Fig. 3. Agarose gel analysis of AO8, AO9, AO11 markers. C:
Chiifu, K: Kenshin, S: Size-marker
앞서 연구된 벼에서의 AOX 관련 유전자에 대한 논문에 따 르면, OsAOX1a가 저온 내성에 관련되며, QTL에 매우 조밀하 게 존재해 서로 연관되어 있는 것을 밝혀냈다[1]. 따라서 본 연구에서는 배추작물에서 내냉성, 내서성 및 환경 스트레스 저항성 품종 육성에 사용할 수 있는 마커개발을 위해 배추의 AOX 합성 관련 EST를 대상으로 ‘지부’와 ‘권심' 사이에서 다 형성을 보이는 4개의 분자 마커를 개발하였는데 이러한 마커 들은 연구자가 손쉽게 이용이 가능한 PCR 증폭 분자 마커이 다. 이는 앞으로 배추작물의 AOX 유전자가 스트레스를 유발 하는 환경 하에서 어떤 세포기작을 이끌어 내는 지에 대한 연구와 환경 내성 형질과 어떤 연관관계를 가지는지를 조사하 는데 중요한 자료로 활용될 수 있으며 또한 환경 스트레스 저항성 배추 품종을 개발하는데 있어서 중요한 육종 도구로써 사용될 수 있다고 생각된다.
감사의 글
본 연구는 농림수산식품부 농림기술관리센터 배추분자마 커사업단 지원에 의하여 이루어 졌으며 이에 감사드립니다.
References
1. Abe, F., K. Saito, K. Miura, and K. Toriyama. 2002. A single nucleotide polymorphism in the alternative oxidase gene among rice varieties differing in low temperature tolerance.
FEBS Lett. 527, 181-185.
2. Ayeh, K. O. 2008. Expressed sequence tags (ESTs) and sin- gle nucleotide polymorphism (SNPs): Emerging molocular marker tools for improving agronomic traits in plant biotechnology. Afr. J. Biotechnol. 7, 331-341.
3. Berhold, D. A. and P. Stenmark. 2003. Membrane-bound diiron carboxylate proteins. Annu. Rev. Plant Biology 54, 497-517.
4. Daisuke, S., E. Nambara, S. Naito, N. Tsutsumi, A. Hirai, and M. Nakazono. 1997. Charactaerization of the gene fam- ily for alternative oxidase from Arabidopsis thaliana. Plant Molecular Biology 35, 585-596.
5. Kim, M. J. and J. S. Chun. 2005. Bacterial community struc- ture in kimchi, a Korean fermented vegetable food, as re- vealed by 16S rRNA gene analysis.Int. J. Food Microbiol.
103, 91-96.
6. Kim, Y. S., Z. B. Zheng, and D. H. Shin. 2008. Growth in- hibitory effects of kimchi (Korean traditional fermented vegetable product) against Bacillus cereus, Listeria mono- cytogenes, andStaphylococcus aureus.J. Food Prot.71, 325-332.
7. Kreps, J. A., Y. Wu, H. S. Chang, T. Zhu, X. Wang, and J. F. Harper. 2002. Transcriptome changes for arabidopsis in response to salt, osmotic, and cold stress.Plant Physiology 130, 2129-2141.
8. Labana, K. S. and M. L. Gupta. 1993. Importance and origin, pp. 1-20, In K. S. Labana, S. S. Banga, and S. K. Banga (eds.), Breeding Oilseed Brassicas,Springer-Verlag Press, Berlin.
9. Lagerctantz, U. and D. Lydiate. 1996. Comparative genome mapping in brassica.Genetics 144, 1903-1910.
10. Levadoux, W. L., M. L. Kalmokoff, M. D. Pickard, and J.
W. D. GrootWassink. 1987. Pigment removal from canola oil using chlorophyllase.J. Am. Oil Chem. Soc. 64, 139-144.
11. Mahmud, T. M. M., J. G. Atherton, C. J. Wright, M. F.
Ramlan, and S. H. Ahmad. 1999. Pak Choi (Brassica rapa ssp. Chinensis L.) quality response to pre-harvest salinity and temperature. J. Sci. Food Agric. 79, 1698-1702.
12. Maxwell, D. P., Y. Wang, and L. Mcintosh. 1999. The alter- native oxidase lower mitochondrial reactive oxygen pro- duction in plant cells. Proc. Natl. Acad. Sci. USA 96, 8271-8276.
13. McDonald, A. E. and G. C. Vanlerberghe. 2004. Branched mitochondrial electron transport in the animalia: Presence of alternative oxidase in several animal phyla.IUMBM Life 56, 333-341.
14. Nagaharu, U. 1935. Genome analysis in Brassica with spe- cial reference to the experimental formation of B. napus and peculiar mode of fertilization. Jpn. J. Bot. 7, 389-452.
15. Prasad, M., R. K. Varshney, J. K. Roy, H. S. Balyan, and P. K. Gupta. 2000. The use of microsatellite for detecting DNA polymorphism, genotype identification and genetic diersity in wheat. Theor. Appl. Genet. 100, 584-592.
16. Sugie, A., N. Naydenov, N. Mizuno, C. Nakamura, and S.
Takumi. 2006. Overexpression of wheat alternative oxidase gene Waox1a alters respiration capacity and response to re- active oxygen species under low temperature in transgenic Arabidopsis. Genes Genet. Syst. 81, 349-354.
17. Thomashow, M. F. 1998. Role of cold-responsive genes in plant freezing tolerance.Plant Physiol. 118, 1-8.
18. Umbach, A. L., F. Fiorani, and J. N. Siedow. 2005.
Characterization of transformed arabidopsis with altered al- ternative oxidase levels and analysis of effects on reactive oxygen species in tissue. Plant Physiol. 139, 1806-1820.
19. Vanlerberghe, G. C. and L. McIntosh. 1997. Alternative oxi- dase: From gene to function.Annu. Rev. Plant Physiol. Plant Mol. Biol.48, 703-734.
초록:배추 alternative oxidase 합성 유전자와 연관된 분자마커 개발 정예솔․정상민*
(동국대학교-서울 생명과학과)
작물의 수량과 품질은 저온 및 고온 스트레스에 영향을 받는 것으로 알려져 있다. 본 연구에서는 배추에서의 스트레스 저항성과 연관된 분자마커를 개발하기 위하여 저온에서의 스트레스 저항성에 영향을 미치는 것으로 알려져 있는 alternative oxidase (AOX) 합성 유전자 관련 분자 마커를 개발하였다. 총 15개의 AOX 합성 유전자 와 관련된Brassica rapa ESTs를 arabidopsis AOX 합성 유전자 염기서열을 이용하여 찾을 수 있었다. 이를 이용하 여 고온에서 상대적으로 약한 ‘지부’품종과 상대적으로 강한 ‘권심’ 사이에서 DNA 염기서열을 조사하여 4개의 ESTs에서 insertion 또는 deletion을 찾았고 PCR로 확인 가능한 4개의 공동우성 마커를 개발하였다. 본 연구에서 개발된 분자마커는 배추 작물에서 환경스트레스 저항성과 유전적 연관성을 확인하는데 유용하게 사용될 수 있다 고 기대된다.