헴철이 풍부한 영양원이 혐기성 세균의 생장과 생존에 미치는 영향:
락토바실러스 가세리 모델연구
이승기, 김 필*
가톨릭대학교생명공학과
Received: February 22, 2021 / Revised: March 6, 2021 / Accepted: March 15, 2021
서 론
소화기말단인대장(large intestine)은동물내관으로써 역할외에는크게주목받지못하다가최근들어서장에는많
은수의뉴런이존재하고, 95% 이상의세로토닌을생성한다
는결과가알려지면서제2의뇌라고지칭할정도로중요성
이심화되어인식되기시작했다[1]. 인간대장의길이와표
면적은각각 1.5 m와 300 m2로알려져있으며, 인간의대장
관(track)에 존재하는 전체 미생물 군집(flora)의 수는 약
1013-14개이상존재한다[2]. 이러한장내균총은차세대염기
서열분석(NGS, next generation sequencing) 기술이발전 함에따라서서식하는미생물들의다양성에대해서많은연 구가 활발히 진행중이다[3]. 인간의 장내균총은 후벽균문 (Firmicutes), 의간균문(Bacteroidetes), 프로테오박테리아문 (Proteobacteria), 방선균문(Actinobacteria) 등의 여러 문 (phylum)으로구성되어있으며, 그종(species)은 400여종이
상인것으로알려져있다[4]. 또한분변고형물의약 30% 이
상을차지할정도로비율이높으며이러한장내균총의활 성은인체의면역, 약효, 소화대사등많은영향을미치는것
으로알려지고있다[5]. 이러한장내균총의연구중에서후
Effect of Heme-rich Nutrient on Anaerobic Bacterial Growth and Survival: A Model Study on Lactobacillus gasseri Seungki Lee# and Pil Kim*
Department of Biotechnology, The Catholic University of Korea, Bucheon 14662, Republic of Korea
#Current address: SL: Dibiome Co., Anyang, Gyeonggi 14102, Korea
Lactic acid bacteria (LAB), belonging to the Firmicutes phylum, lack heme biosynthesis and, thus, are char- acterized as fermentative and catalase-negative organisms. To verify the hypothesis that heme-rich-nutrients might compensate the heme-biosynthesis incapability of non-respiratory LAB in animal gut, a heme-rich- nutrient was fed to a dog and its fecal microbiome was analyzed. Firmicutes abundance in the feces from the heme-rich-nutrient-fed dog was 99%, compared to 92% in the control dog. To clarify the reason of increased Firmicutes abundance in the feces from the heme-rich-nutrient-fed dog, Lacobacillus gasseri were used as model anerobic LAB to study a purified heme (hemin). The anaerobic growth of L. gasseri in the medium with 25 µM hemin supplementation was faster than that in the medium without hemin, while the growth in the 50 µM hemin-supplemented medium did not vary. Cellular activities of the cytochrome bd complex were 1.55 ± 0.19, 2.11 ± 0.14, and 2.20 ± 0.08 U/gcell in the cells from 0, 25, and 50 µM hemin-supplemented medium, while intracellular ATP concentrations were 7.90 ± 1.12, 11.95 ± 0.68, and 12.56 ± 0.58 µmolATP/gcell, respectively. The ROS-scavenging activities of the L. gasseri cytosol from 25 µM and 50 µM hemin-supple- mented medium were 68% and 82% greater than those of the cytosol from no hemin supplemented-medium, respectively. These findings indicate that external hemin could compensate the heme-biosynthesis incapa- bility of L. gasseri by increasing the cytosolic ROS-scavenging and extra ATP generation, possibly through increasing the electron transfer. Increase in the number of anaerobic bacteria in heme-rich-nutrient-fed animal gut is discussed based on the results.
Keywords: Heme-rich-nutrient, lactic acid bacteria, anaerobic bacteria, gut microbiome, dog
*Corresponding author
Tel: +82-2-2164-4922; F. +82-2-2164-4865 E-mail: [email protected]
벽균(Firmicutes)에속하는 Lactobacillus 종들과동물과의 상관관계에대한연구들이많이진행되고있는데, 대표적으 로외부의알레르겐과바이러스로부터인체를보호하는기 능이알려진 Lactobacillus johnsonii [6], 제 2형당뇨와연 관성이있다고알려진 Lactobacillus acidophilus [7], 유산균 종들의지방대사연관관계에따른항비만역할[8] 등을예 로들수있다. 이러한인체건강의영향력때문에유산균들 은여러프로바이오틱스제품들에포함되어관련시장이확 대되고 있는 추세이며, 국내 프로바이오틱스시장규모는
2019년기준으로약 1898억원규모인것으로알려지고있
다[9].
헴철(heme or haem)은동물, 식물, 미생물등모든호흡 대사(respiratory metabolism) 생명체에서발견되는분자로 서, 포르피린(porphyrin)과철이온(iron)의배위결합구조를
기본으로하여다양한곁사슬(side chain)을갖는분자들의
총칭이다. 헴철은보결분자단(prosthetic group)으로서단백
질과 결합된 여러 종류의 헴단백(hemoprotein = heme-
conjugated protein)의형태로자연계에폭넓게분포하며, 산소분자(O2)의전달(적혈구내헤모글로빈, 근육내미오 글로빈, 콩과식물의레그헤모글로빈), 전자의전달(여러종 류의시토크롬류, 리덕타제, 옥시게나제등), 독성산소의제 거(카탈라제, 페옥시다제등)와같은생명체의에너지생산 과정이나활성산소종(ROS: reactive oxygen species)에대한 제거에필수적인역할을담당한다. 유산균류와같이헴철생 합성능력이결실된혐기성미생물들은호흡대사(respiration) 를수행할수없으므로발효대사(fermentation)만을유일한 에너지생성대사로이용하게되며, 또한호기적환경에서자 연적으로발생되는활성산소종에대하여취약한공통된특 성을나타낸다. 따라서생태학적적소(ecological niche)도동 물의장관과같은혐기적환경(anaerobic environment)으로 제한되는특징을보인다.
본연구에서는헴철이풍부한영양원을공급받는경우에 동물의분변으로부터혐기성미생물군집의변화를확인하 고, 단일성분의헴철(헤민, hemin: 소혈액에서분리한 heme B의 염소 착염분자)을 혐기성 유산균으로서 Lactobacillus
gasseri에공급하였을때의생장특성과유산균세포에서의
에너지와활성산소방어능을조사하여동물장관에분포하 는전체혐기성세균들의변화를추론하였다.
재료 및 방법
동물식이에 따른 분변채취와 균총분석
헴철영양원의식이실험을위한동물로는동일한실내생 활로인간과장내미생물군집이유사할것으로추정되는소 형반려견을사용하였다. 4세령소형견 2 기(암컷푸들, 체
중은 모두 3.5 kg)에 동일한시중 사료(Ultragrainfree for adult dog, Natural Balance Korea Inc., Korea)를 하루에 체중 1 kg당 25 g씩공급하면서 4주간동일한실내환경
(전라도 광주소재실험자 거주아파트실내)에서 28일동
안 순화(domestication) 과정을 통해 장내 균총을 동기화
(synchronization)하였다. 순화과정동안동물에게는정량의 사료와음용수(자유급식) 이외에는추가적인식이를공급하 지않았고, 실험자이외에는다른사람과의접촉을제한하였 다. 29일부터소형견 2 기는분리된별실로이동하여시험기 간동안상호접촉이없도록유지하였다. 거주공간분리이 후 6일동안 1 기에게는 1 g의헴철이포함된사료과립을 매일 1회씩일반사료와함께공급하였고, 다른 1 기에게는 일반사료이외에는헴철포함사료과립을공급하지않았 다. 헴철이포함된사료과립(HemoP, Hemolab Co., Korea)을 구입하여동물의헴철이풍부한식이로사용하였고, 이과립 의헴철함량은 1 g당 16.9 mg(철분함량 1.5 mg/g)이었다.
소형견의체중은시험기간동안 2일마다 1회씩측정하였 고, 특이한행동이나건강상의변화를실험자가지속적으로 관찰하였다. 7일차에동물의분변시료는배출즉시 50 ml 튜 브에넣고밀봉상태로 -20℃로동결하였다. 분변시료의균총 변화는마이크로비옴분석기관(CheonLab Co., Korea)에서
16S rRNA 유전자의서열분석을통해확인하였다.
모델 유산균과 배양방법
각각후벽균문(Firmecutes)와방선균문(Actinobacteria)에 속한 유산균으로써 Lactobacillus gasseri KCTC3163와 Bifidobacterium bifidium KCTC3418 표준균주는한국생명 공학연구원생물자원센터(KCTC)에서분양받아실험에사용 하였다. 표준균주의유전체정보는미국국립생명공학정보 사이트(www.nlm.ncbi.gov)를통해검색하였다.
유산균의배양에는복합배지(MRS Broth, KisanBio Co.,
Seoul, Korea)를사용하였다. 헴철의표준으로사용한헤민
(hemin, Merck Inc., Germany)은 1 M NaOH 용액에헤민
25 mM가되도록용해시킨표준용액을제조한후실험에사
용하였다. 150 ml-시럼바이얼(Bellco Glass Inc., USA)에
MRS 배지와헤민표준용액또는동일부피의 1 M NaOH
용액을 100 ml에맞추어넣고, 알루미늄캡을씌운고무마
개로밀봉하였다. 주사바늘을연결한진공펌프로 40초간용 기내헤드스페이스의공기를제거한후고순도질소를 30초 간주입하기를 3회반복하여용기내잔존산소를제거한후 고압멸균기에서 121℃로 15분간멸균하였다. 활발히생장중 인유산균전배양액(seed culture)은초기흡광도가 OD600nm
= 0.1이되도록 10 ml 주사기로산소의접촉없이멸균된배
지에 접종하였고, 세포 침강을 막기 위해 낮은 진탕속도
(100 rpm)의진탕배양기에서 37℃를유지하며배양하였다.
배양액시료는일정시간간격으로주사기를이용하여 1 ml 씩채취하여분석에사용하였다. 세균의생장은분광광도계 (BioSpectrometer, Eppendorf Inc., Germany)를 이용하여 흡광도를측정하였고, 1 OD600nm = 0.265 g of cell/l의세포 농도로환산하여계산에사용하였다.
시토크롬 bd 복합체 활성도의 측정
시토크롬 bd 복합체의 활성도는 TMPD(N,N,N',N'-
tetramethyl-p-phenylene diamine 또는 Wuster’s blue) 산 화법을사용하였다[10−12]. 이방법은 TMPD로부터의전자 가세균표면의시토크롬 bd 복합체내부에결합된보결분 자단 heme B595나 heme D로전달되는경우산화된 TMPD 가정량적으로발색되는원리를이용한것이다. 접종후 4시 간동안배양한유산균의배양액을채취한후원심분리(4℃, 2600 g, 10분)하여균체를분리하고 0.9 % NaCl 용액으로 2회세척하였다. 획득한세포펠렛은 100 µl의 0.9% NaCl 용 액에분산시킨후 1.4 ml의인산완충용액(100 mM, pH 7.0) 과 5 µl의 TMPD 표준용액(0.54 M TMPD, Merck Inc.)를
혼합하고상온에서 10분동안 520 nm의파장에서흡광도를
측정하였다. 산화된 TMPD의환산계수는 1 OD520nm = 6.1 /mM·cm로사용하였고, 시토크롬 bd 복합체의활성 1 단위 (unit)는일정 세포량에의해 1분동안산화되는 TMPD의 mmol수(U = mmol of oxidized TMPD formation/min)로 정의하여사용하였다[13].
세포질 ATP 농도의 측정
세포질에 존재하는 ATP 농도는 비드비터(beadbeater
Model 607, Biospec Inc., Bartlesvile, USA)를사용하여물 리적으로세포를파쇄한후용출된세포질시료로부터결정 하였다. 배양액 1 ml으로부터원심분리(4℃, 3,000 g, 15분) 후펠렛을 증류수 800 µl에현탁하고 0.2 g의글래스비드 (glass beads 212-300 micron, Merck Inc.)를첨가했다. 현
탁액을포함한 1.5 ml-튜브는비드비터를이용하여 30초진
동(700 W)과 1분냉각과정을총 6회운전하여세포를파쇄 시켰다. 파쇄액은원심분리(4℃, 12,000 g, 5분)로잔여물을
제거한이후의상등액을채취하여포함된 ATP 농도를 ATP
assay kit (Merck Inc.)의시료로사용하였고, 제조자가공급한 매뉴얼에따라발광광도기(Turner BioSystems Luminometer, Thermo Fischer Scientific Inc., USA)를이용하여시료의 발광정도를측정하였다.
세포질 라디칼 소거능의 측정
세포질의라디칼소거능은세포파쇄액으로부터 DPPH 라 디칼(2,2-diphenyl-1-picrylhydrazyl radical) 분자가세포질 로부터전자를획득하여중화되면정량적으로발색되는원
리를사용하였다[14, 15]. 접종후 2시간과 4시간의배양액 1 ml의원심분리(4℃, 3,000 g, 10 min) 후얻어진 펠렛을 OD600nm = 1이되도록인산완충용액(100 mM, pH 7)에현탁 한후위에기술한방법으로세포를파쇄시켰다. 원심분리 (4℃, 12,000 g, 5분) 이후상등액 500 µl를채취한후동일 부피의 ethyl acetate를혼합하고강하게섞어주었다. 혼합 물을다시원심분리(4℃, 12,000 g, 5분)하여층분리를유도 한후 ethyl acetate 층만을분리하여잔여물을필터(0.22µm pore size syringe filter, Merck Inc.)를이용하여제거하였 다. 필터액에서 20 µl를채취하여 280 µl의 DPPH 표준용액 (5 mg DPPH를 100 ml ethanol에용해)과혼합시킨후 96-
well 플레이트에옮기고상온에서커버를씌워빛을차단하
고 30분동안정치하였다. 96-well 플레이트는다중플레이트 검출기(Multiplate reader, Biorad Inc., USA)를 사용하여
570 nm에서흡광도를측정하였다. 시료의라디칼소거능은
표준 DPPH 용액에서얻은표준곡선과비교하여다음공식
을통해전환하였다.
ROS-scavenging activity (%) = (Acontrol − Asample)/Acontrol× 100
결 과
헴철 영양원을 공급받은 동물 분변의 분석
28일의 순화과정 이후 헴철미생물 사균체 과립 총 6 g (16.9 mgheme/g 포함)을 6일동안공급받은소형견 1 기와공 급받지않은소형견 1기는모두시험기간 6일동안특이한 행동, 활동성의변화나발진과같은이상징후들이관찰되지 않았고, 체중의급격한변화도관찰되지않았다(Fig. 1A). 헴 철영양원공급개시후 7일차에발생한동물의분변은분석
기관에서 16S rRNA 서열분석을통해균총변화를추적하였
다(Fig. 1B, Table S1). 헴철미생물사균체과립을공급받지 않은대조군의분변에서는총 172 종(species)이발견되었으 며, 91.9%의 비중으로 후벽균문(Firmicutes)에 속한 세균 109종(species)과, 8.0%의 비중으로 프로테오박테리아문 (Proteobacteria)에속한세균 34종이메이져비중으로발견 되었다. 이외에도의간균문 Bacteriodetes) 12종, 방선균문 (Actinobacteria) 13종, 사카리박테리아문(Saccharibacteria) 1종, 시너지스테츠문(Synergistes) 1종, 베루코마이크로비아 문(Verrucomicrobia) 2종, 푸소박테리아문(Fusobacteria)
1종에속한종들이각각 1% 미만의마이너비중으로발견되
었다(Table S1 A). 반면헴철미생물사균체를공급받은시
험군의분변에는전체 202종류의종중에서 99.1%의비중
으로후벽균문(Firmicutes)에속한세균이 145종발견되었 다. 프로테오박테리아문(Proteobacteria) 17종의 비중은
0.5%로 감소하였다. 이외에도 방선균문(Actinobacteria) 19종, 의간균문(Bacteroidetes) 15종, 사카리박테리아문 (Saccharibacteria) 4종, 시피로체츠문(Spirochaetes) 1종, 푸 소박테리아문(Fusobacteria) 1종이 1% 미만의마이너비중 으로발견되었다(Table S1 B).
따라서헴철이풍부한식이 1 g씩 6일동안공급받은개의
분변은공급받지않은개의분변보다후벽균문(Firmicutes)
의균총비중이증가되면서프로테오박테리아문(Proteobacteria) 의균총비중이감소하는양상을나타났고, 발견되는장내 균총의종류도 172종에서 202종으로다양성이증가하였다.
헤민 공급에 의한 L. gasseri의 생장 변화
헴철미생물사균체과립공급에의해개의장내균총에서 유산균류가 다수 포함된 후벽균문(Firmicutes)이 증가된
Fig. 1의결과로부터식이에포함된헴철분자가후벽문군에
속한혐기성유산균류의균총의비중을증가시키는데기여 했을것으로추정되었다. 그러나상기실험에사용된헴철미 생물사균체과립에는헴철분자뿐만아니라여러가지세 포조성물질들이함께혼재되어있었으므로, 헴철분자가유 효물질임을명확하게증명하기위하여헴철의염소(Cl) 착염 분자인헤민(hemin, Merck Inc.)을단일성분으로하고, 혐 Fig. 1. Variations of weight and fecal microbiomes in bacterial phylum from a heme-rich-nutrient-fed dog.
Panel A indicates the graph for weight variation of dogs (white bar for control-nutrient-fed and black bar for the heme-rich-nutrient- fed dog). Panel B indicates the fecal microbiomes of a control and heme-rich-nutrient-fed dogs.
The variation of fecal microbiome in bacterial species is listed in detail at supplementary Table S1.
기성유산균류의모델로결정한 Lactobacillus gasseri의배 양에농도별로추가하면서헤민첨가가유산균생장에미치 는영향을조사하였다. 혐기성유산균류의모델은프로바이 오틱스로써의기능성과전체유전체서열정보가알려진후
벽군문에속한종을기준으로선택하였다.
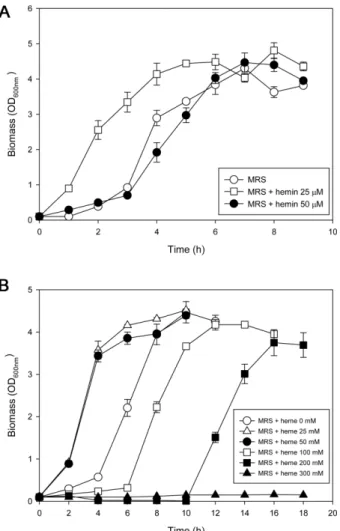
MRS를기본배지성분으로하여다른농도의헤민을포
함한시럼바이알내부를혐기조건으로유지하면서 L. gasseri 를접종하여생장을비교하였다(Fig. 2A). 최대비생장속도 (specific growth rate, 1/h)에는헤민첨가에따라큰차이가 없었으나, 헤민 25 µM 첨가배지에서생장한 L. gasseri는헤 민을무첨가한대조군보다지연기(lag phase)가단축되면서 빠르게 생장했다. 반면 헤민 50 µM 첨가 배지에서는 L.
gasseri의생장이다시느려져서헤민무첨가배지에서의생
장곡선과크게차이가없었다. L. gasseri는낮은농도범위
의헤민이존재하는배지에서는헤민무첨가의경우보다일 찍생장이시작되었고, 높은농도범위의헤민이존재하는배 지에서는생장의시작시간이느려졌다. 헤민첨가농도에따 른호기조건(고무셉터로막은시럼바이얼용기대신에면전
으로막은 250 ml-플라스크용기에서의배양, 그외배양조
건은혐기조건배양과동일)의 L. gasseri 생장곡선도헤민 추가에따라혐기조건에서와유사한양상을보였다(Fig. 2B).
즉, 헤민 25 및 50 µM 첨가배지에서는무첨가배지의경우 보다빠르게생장이시작되었고, 100 µM 이상의헤민첨가 조건에서는생장의시작시간이점차느려져서헤민 300 µM 첨가배지에서는생장이거의일어나지않았다. 따라서낮은 농도구간의헤민첨가에의해 L. gasseri의생장지연기가단 축됨에의한생장이빨라졌고, 산소접촉의정도에따라서 생장이촉진되는헤민의적정한농도범위가다르게나타났다. 헤민 공급에 의한 L. gasseri의 시토크롬 bd 복합체 활성도 변화
L. gasseri는호흡대사를하지않음에도불구하고유전체
검색에서는다른호흡대사종에서발견되는전자전달계의 단백질성분인시토크롬 bd 복합체를구성하는단백질들의 유전자(cydABCD)가존재한다[16]. 따라서 L. gasseri는시 토크롬 bd 복합체를불활성상태인아포단백질(apoproteins:
보결분자단이포함되지않은단백질부분) 형태로포함하고 있으며, 외부로부터공급된헴철에의해생물학적으로활성 상태인헴-접합단백질(heme-conjugated protein)로바뀔수 있음을암시하였다. 이가설을확인하기위해혐기배양중 Fig. 2. Growth curves of L. gasseri in media with and without
hemin supplementation.
Panel A indicates the anaerobic conditional growth curve and panel B the aerobic conditional growth curve. Legends are in the graph panels.
Table 1. Activity of cytochrome bd complex and cytosolic ATP concentration of L. gasseri grown in hemin supplemented medium.
Growth conditions No hemin
supplementation
25 μM hemin supplementation
50 μM hemin supplementation A. Activity of cytochrome bd complexa (U/g of cell) 1.55 ± 0.19 2.11 ± 0.14 2.20 ± 0.08 B. ATP concentration in cytosol (μmol/g of cell) 7.90 ± 1.12 11.95 ± 0.68 12.56 ± 0.59
aOne unit of cytochrome bd complex activity was defined as the amount of oxidized TMPD formation per minute by whole cell (U = mmol of oxidized TMPD/min). Actively growing cells in MRS-basal medium were sampled from anaerobic culture at 4 h after inoc- ulation. Data are represented as mean ± SD from three biological repeats.
활발히생장하는시기의세포(접종후 4시간)를채취하여호 기조건에서시토크롬 bd 복합체로의전자흐름을측정하였다
(Table 1A). 헤민무첨가 MRS 배지에서혐기적으로생장하
던대조군 L. gasseri는 외부에서공급된 TMPD를 1.55 ± 0.19 mmol/min·g of cell의속도로산화시켰다. 반면 25 및
50 µM 헤민이첨가된 MRS 배지에서혐기적으로생장하던
L. gasseri는더 높은 속도로 (각각 2.11 ± 0.14 및 2.20 ± 0.08 mmol/min· g of cell) TMPD를산화시켰다. 따라서외부
에서공급된헴철이포함된배지에서생장한 L. gasseri의시
토크롬 bd 복합체로의전자전달활성이증가하였다.
헤민 공급에 의한 L. gasseri의 세포 내 ATP 농도 변화
세포표면에존재하는시토크롬 bd 복합체의전자전달활
성에따라세포내 ATP 변화를추적하기위하여위시료의
세포를물리적으로파쇄하였다(Table 1B). 혐기적환경에서
생장한 대조군 L. gasseri의 세포질에 존재하는 ATP는
7.90 ± 1.12 µmol/g of cell인반면 25 및 50 µM 헤민이첨가 된배지에서혐기적으로생장하던 L. gasseri의세포질에는 각각 11.95 ± 0.68과 12.56 ± 0.59 µmol/g of cell이측정되 었다. 따라서헤민이무첨가된배지에서생장하던세포의세 포질보다헤민이공급된배지에서생장하던세포의세포질 에는대조군보다 50% 가량높은에너지상태를나타냈다. 헤민 공급에 의한 L. gasseri 세포질의 라디칼 소거능 변화
외부헤민공급에의해 L. gasseri에서시토크롬 bd 복합 체로의전자전달활성이높아지고동시에세포질내부의 ATP 가높아진것을확인하였으나, 절대혐기환경인동물의장관 에서후벽균문에속한유산균류가헴철식이에의해에너지 증가로인한개체수증가를설명하기에는충분하지않았다. 호기성생물종에서의헴철은전자전달과정의산화·환원효 소들뿐아니라활성산소방어에필요한카탈라제, 페옥시다 제에필요하므로, 외부에서공급된헴철이 L. gasseri 세포 질에서불활성아포단백질으로존재하는 ROS-방어효소들을 활성화시킬수도있었다. 이가설을증명하기위해헤민첨
가에따라혐기적환경에서생장하는 L. gasseri 세포질의라
디칼소거능의변화를측정하였다(Fig. 3). 25 및 50 µM 헤 민첨가조건에서혐기적으로생장한 L. gasseri 세포질시 료(접종후 2시간)는헤민무첨가조건의세포질시료보다 각각 68%와 82%가상대적으로높은 DPPH 라디칼소거능 을보였다. 접종후 4시간의 L. gasseri 세포질시료들도접 종후 2시간대의라디칼소거능과동일한경향을보였다. 4시간대의헤민무첨가조건에서의대조군시료보다 25 및
50µM 헤민첨가조건의시료는라디칼소거능이각각 13%
및 10% 더높았다. 또한 L. gasseri의유전체정보에서는헴 철의존성카탈라제의유전정보가확인되었다[17]. 그러므
로 L. gasseri 세포질에아포단백질형태로존재하는헴철의
존성페옥시다제또는카탈라제가외부헴철공급에의해활 성형인헴-접합형태로전환되어세포질의전체활성산소에 대한방어능력이증대되었으며이로인해생장개시에필요 한세포질의적정환원도에일찍다다른것으로생각되었다.
고 찰
1일당 16.9 mg씩의헴철을 6회추가로공급받은개의분 변에서유산균류가다수포함된후벽균문(Firmecutes)의균
총비중이증가된것(Fig. 1)은헴철생합성이결핍된혐기
성세균의호흡성에너지생성이나활성산소에방어에관계 되는 아포단백질들(apoproteins)에 보결분자단(prosthetic
group)인 heme이외부로부터공급되어활성형인헴-결합단
백(heme-conjugated proteins) 형태로전환되어활성을가짐 으로써생장이나생존성이증대된것으로사료되었다. 비록 본연구에서사용한 2 기의개분변에서의미생물군집의변 화는통계적으로유의미하지는않았으나, 타동물의사양시 험결과들(헴철사균체과립을전체식이량의 1%인사료를 5일간공급받은마우스 5 기의분변균총에서후벽균문의비
중이증가했던결과나동일과립이전체식이량의 0.5%인
사료를 1개월간공급받은닭들 200 기의맹장및분변에서
의 MRS 배지에서생장하는전체유산균류의생균수가증가
Fig. 3. Cytosolic radical scavenging activity of L. gasseri grown in media with and without hemin supplementation.
The DPPH radical scavenging by the cytosol from MRS medium is in white bar, the cytosol from hemin 25 µM supplementation is in shade bar, and the cytosol from hemin 50 µM supplemen- tation is in black bar. Cells were taken from anaerobic culture at 2 and 4 h of cultivation and set to OD600nm = 1 before disruption to release cytosol. The error bars indicate the SD from three bio- logical repeats.
했던결과)와유사하였다[18]. 또한소혈액유래의헴철을 배지에첨가하는경우 Lactobacillus lactis의균체량이증가
나[16] 사멸기의지연[19]을보고한논문들도본논문에서의
결론과 일치한다. Lactobacillus plantarum이담즙산(bile acid)에의해용해된 B. subtilis의헴철-포함세포질용출물 에의해과산화수소수(H2O2)에대한해독능력이증가되었던 논문도본연구의결론을지지하였다[20].
헴철공급의영향을연구하기위해본논문의혐기성유 산균으로 사용한 L. gasseri는 1980년대에 Lauer and Kandler가여성의질에서처음분리한유산균[21]으로, 인체 섭취결과지방의약 8%를감소시키는효과나[22] 자궁내
막증, 생리통및월경통등의완화기능[23] 등의기능성이보
고된유산균이다. 후벽균문(Firmecutes)에속한 L. gasseri 이외에도방선균문(Actinobacteria)에속한혐기성유산균인 Bifidobacterium bifidium에서도헤민첨가에따라정도의 차이가있으나소규모의생장촉진효과(Fig. S1)나세포질의
라디칼소거능력증대(Fig. S2)가발견되었다. 따라서여러
프로바이오틱스제품들에사용되는유산균류에헴철을공급 하는경우에너지추가생산에의한생장증대나산소접촉 환경에서의활성산소방어력증대에의한생존성증대가가 능할수있을것이다. 단지, 공급되는헴철의농도에따른 L.
gasseri의생장결과(Fig. 2)가암시하듯이공급되는헴철의
농도가과도한경우에는오히려생장이나생존력이감소할 가능성도존재하기때문에, 유산균의종에따라적절한헴철 공급농도범위가별도로결정되는후속연구들이필요할것 이다.
Acknowledgment
The authors appreciate Dibiome Co. for the helps testing dogs and collecting feces samples.
Conflict of Interest
The authors have no financial conflicts of interest to declare.
References
1. Kim DY, Camilleri M. 2000. Serotonin: a mediator of the brain-gut connection. Am. J. Gastroenterol. 95: 2698.
2. Savage D. 1977. Microbial ecology of the gastrointestinal track.
Ann. Rev. Microbiol. 31: 107-133.
3. Jovel J, Patterson J, Wang W, Hotte N, O'Keefe S, Mitchel T, et al.
2016. Characterization of the gut microbiome using 16S or shotgun metagenomics. Front. Microbiol. 7: 459.
4. D’Argenio V, Salvatore F. 2015. The role of the gut microbiome in the healthy adult status. Clin. Chim. Acta 451: 97-102.
5. Cummings JH, Macfarlane GT. 1991. The control and conse- quences of bacterial fermentation in the human colon. J. Appl.
Bacteriol. 70: 443-459.
6. Fujimura KE, Demoor T, Rauch M, Faruqi AA, Jang S, Johnson CC, et al. 2014. House dust exposure mediates gut microbiome Lactobacillus enrichment and airway immune defense against allergens and virus infection. Proc. Natl. Acad. Sci. 111: 805-810.
7. Halawa MR, El-Salam MA, Mostafa BM, Sallout SS. 2019. The gut microbiome, Lactobacillus acidophilus; relation with type 2 diabetes mellitus. Curr. Diabetes Rev. 15: 480-485.
8. Arora T, Singh S, Sharma RK. 2013. Probiotics: interaction with gut microbiome and antiobesity potential. Nutrition 29: 591- 596.
9. Korean Ministry of Food and Drug Safety. 2019. KFDA Press release https://www.mfds.go.kr/brd/m_99/down.do?brd_id=
ntc0021&seq=43656&data_tp=A&file_seq=43651.
10. Lorence R, Carter K, Gennis RB, Matsushita K, Kaback HR. 1988.
Trypsin proteolysis of the cytochrome d complex of Escherichia coli selectively inhibits ubiquinol oxidase activity while not affecting N, N, N', N'-tetramethyl-p-phenylenediamine oxidase activity. J. Biol. Chem. 263: 5271-5276.
11. Jünemann S, Wrigglesworth JM. 1994. Antimycin inhibition of the cytochrome bd complex from Azotobacter vinelandii indi- cates the presence of a branched electron transfer pathway for the oxidation of ubiquinol. FEBS Lett. 345: 198-202.
12. Fujimoto M, Yamada A, Kurosawa J, Kawata A, Beppu T, Takano H, Ueda K. 2012. Pleiotropic role of the Sco1/SenC family copper chaperone in the physiology of Streptomyces. Microb.
Biotechnol. 5: 477-488.
13. Frangipani E, Haas D. 2009. Copper acquisition by the SenC protein regulates aerobic respiration in Pseudomonas aeruginosa PAO1. FEMS Microbiol. Lett. 298: 234-240.
14. Okawa M, Kinjo J, Nohara T, Ono M. 2001. DPPH (1,1-diphenyl- 2-picrylhydrazyl) radical scavenging activity of flavonoids obtained from some medicinal plants. Biol. Pharm. Bull. 24:
1202-1205.
15. Afify AE-MM, Romeilah RM, Sultan SI, Hussein MM. 2012. Anti- oxidant activity and biological evaluations of probiotic bacte- ria strains. Int. J. Acad. Res. 4: 131-139.
16. Brooijmans R, Smit B, Santos F, Van Riel J, de Vos WM, Hugen- holtz J. 2009. Heme and menaquinone induced electron trans- port in lactic acid bacteria. Microb. Cell Fact. 8: 28.
17. Maresca D, Zotta T, Mauriello G. 2018. Adaptation to aerobic environment of Lactobacillus johnsonii/gasseri strains. Front.
Microbiol. 9: 157.
18. Hemolab Inc., 2020. Development of heme-iron containing non-GMO feed from breeding. IPET project final report. Reg- istered number: 11-15430001003152-1543001.
19. Lechardeur D, Cesselin B, Fernandez A, Lamberet G, Garrigues C, Pedersen M, et al. 2011. Using heme as an energy boost for lactic acid bacteria. Curr. Opin. Biotechnol. 22: 143-149.
20. Yu T, Kong J, Zhang L, Gu X, Wang M, Guo T. 2019. New crosstalk between probiotics Lactobacillus plantarum and Bacillus subtilis.
Sci. Rep. 9: 13151.
21. Lauer E, Kandler O. 1980. Lactobacillus gasseri sp. nov., a new species of the subgenus Thermobacterium. Zentralblatt für Bakteriologie: I. Abt. Originale C: Allgemeine, angewandte und ökologische Mikrobiologie. 1: 75-78.
22. Kadooka Y, Sato M, Ogawa A, Miyoshi M, Uenishi H, Ogawa H, et al. 2013. Effect of Lactobacillus gasseri SBT2055 in fermented
milk on abdominal adiposity in adults in a randomised con- trolled trial. Br. J. Nutr. 110: 1696-1703.
23. Itoh H, Uchida M, Sashihara T, Ji Z-S, Li J, Tang Q, et al. 2011.
Lactobacillus gasseri OLL2809 is effective especially on the menstrual pain and dysmenorrhea in endometriosis patients:
randomized, double-blind, placebo-controlled study. Cyto- technology 63: 153-161.