*Received: August 7, 2013 / Revised: August 30, 2013 Accepted: September 2, 2013
†Corresponding author: Jin‐Kyung Kim. Department of Biomedical Science, College of Natural Science, Catholic University of Daegu, 330 Geumrak‐Ri, Gyeoungsan‐Si 700‐712, Korea.
Tel: +82‐53‐850‐3774, Fax: +82‐53‐850‐3774 e-mail: [email protected]
ⒸThe Korean Society for Biomedical Laboratory Sciences. All rights reserved.
J. Exp. Biomed. Sci. 2013, 19(3): 224~232 pISSN : 1738-3226
Cucurbitacin I, a Natural Cell‐permeable Triterpenoid, Suppresses Colitis‐associated Colon Carcinogenesis in Mice
Hyeon Jin Kim and Jin‐Kyung Kim†
Department of Biomedical Science, Catholic University of Daegu, Gyeongsan‐Si 700-712, Korea
Cucurbitacins are a natural cell‐permeable triterpenoid compound isolated from Cucurbitaceae and Cruciferae.
Cucurbitacins have been used as folk medicine because of their anti‐inflammatory and analgesic effects. In the present study, we investigate the anti‐cancer effects of cucurbitacin I on colitis‐associated colon carcinogenesis induced by azoxymethane (AOM)/dextran sodium sulfate (DSS) in BALB/c mice. Cucurbitacin I treatment attenuated loss of body weight and decreased the number of colon tumors. Western blot analysis showed that cucurbitacin I treatment significantly inhibited the protein expression of inducible nitric oxide synthase (iNOS), tumor necrosis factor (TNF)‐α and interleukin (IL)‐6. These results suggest that cucurbitacin I suppressed inflammatory reaction and tumor development in colitis‐associated colon carcinogenesis.
Key Words: Cucurbitacin I, AOM/DSS, Colitis‐associated colon carcinogenesis
서 론
대장암은 결장과 직장에 생기는 악성종양으로 대부분 대장의 점막에 발생한다. 2012년 통계청이 발표한 국가암 등록통계에 의하면, 2010년 대장암의 총 발생자는 인구 십만 명당 20,711명으로 1999년부터 계속 증가하는 추세 를 보이고 있다 (Korean National Statistical Office, 2012).
대장암의 주요한 발생요인으로는 식생활의 서구화로 인 한 육류의 과다 섭취, 비만, 음주, 가족력, 염증성 장질환 등이 거론되고 있다. 염증성 장질환 (IBD, inflammatory bowel disease)은 대장 내벽에 염증이 발생하는 질병으로 복통, 설사, 발열 등의 임상적인 증세를 보이는 질환이다.
IBD는 궤양성 대장염 (UC, ulcerative colitis)과 크론병 (CD, Crohn’s disease)으로 분류하는데 (Gillen et al., 1994), UC 환자의 대장암 발병비율은 일반인에 비해 10배 이상
높은 것으로, CD 환자의 경우에는 4~7배 이상 높은 것으 로 알려져 있다 (Hale et al., 2012).
대장염으로 인한 대장 상피 세포의 손상은 대장 내에 존재하는 다수의 면역세포들을 활성화시켜 염증성 매개 물질인 산화질소 (NO, nitric oxide), 프로스타글란딘 (PG, prostaglandin)E, tumor necrosis factor (TNF)‐α, interlukin (IL)‐6 등을 분비하게 된다 (Zhang, 2007). NO는 L‐arginine 의 guanidinonitrogen이 NO synthase (NOS)에 의해 변환되 면서 생성되는데, 이렇게 생성된 과도한 NO는 조직손상, 신경 퇴행성 질병을 유발하고 혈관 투과성을 증가시켜 부종을 일으킨다 (Yun et al., 1996). NO 합성효소인 NOS 는 neuronal NOS (nNOS), endothelial NOS (eNOS), inducible NOS (iNOS)의 3종류가 존재하며 이중 iNOS가 면역반응에 의한 염증 조절 인자로 중요한 역할을 한다 (McCartney‐Fransis et al., 1998). PG는 면역세포를 포함한 다양한 세포에 존재하는 cyclooxygenase (COX)라는 효소 에 의해 생성된다. COX는 COX‐1과 COX‐2의 두 종류의 아형이 존재하며, COX‐1이 생체 내의 대부분 조직에서 일정하게 발현하는 것과는 대조적으로 COX‐2는 사이토 카인, 성장인자, 산화적 스트레스 등과 같은 이차적인 자 극에 의해 발현이 증강되어 염증 반응을 매개하는 것으 로 알려져 있다 (Sarkar et al., 2008).
Original Article
TNF‐α와 IL‐6는 대표적인 염증성 사이토카인으로 면역 세포에서 분비되어 또 다른 염증매개인자의 생성과 분비 를 유도하여 염증 반응을 촉진하는 것으로 알려져 있다 (Tracey, 2002). 일부 연구에 의해 염증성 사이토카인이 암 의 발생 과정에 매우 밀접하게 관여한다는 것이 밝혀져 있 다 (Sandborn and Hanauer, 1999). TNF‐α와 IL‐6의 분비를 억제하면 대장염의 완화 및 염증으로 매개된 대장암의 발 생을 억제하였다는 결과가 도출되었다 (Popovanova et al., 2008).
Phytochemical은 식물이 자신과 경쟁하는 식물의 생장 방해 및 각종 미생물, 해충 등으로부터 자신을 보호하기 위해 생성하는 물질로, 현재 1,000여 종류가 밝혀져 있다.
Phytochemical은 DNA의 손상 억제, 산화와 노화방지에 관여할 뿐만 아니라 인체 내에서 면역시스템을 자극하거 나 암의 성장을 돕는 발암 물질로부터 보호하는 역할을 하는 것으로 알려져 있다 (Sun et al., 2002). 이러한 생리 활성 기능을 바탕으로, 최근 phytochemical을 이용하여 보 다 효과적으로 질병을 완화하는 기능을 갖는 의약품 및 기능성 식품의 개발에 대한 연구가 활발이 이루어지고 있다 (Kim et al., 2009).
Cucurbitacin은 박과작물에 함유되어 있는 tetracyclic triterpene 성분으로 cucurbitacin A로부터 cucurbitacin T에 이르기까지, 약 18여종이 밝혀져 있다 (Chen et al., 2012).
Cucurbitacin의 생리활성에 대한 연구가 활발하게 이루어 져 항염증, 항암, 진통 효과에 효과가 있는 것으로 알려져 있다 (Alghasham, 2013). Cucurbitacin B는 유방암 세포와 췌장암 세포의 세포주기의 진행을 억제하고 세포사멸을 촉진함으로써 항종양 활성을 나타내는 것으로 밝혀졌다 (Thoennissen et al., 2009; Duangmano et al., 2012).
Cucurbitacin E는 전립선암 세포의 증식을 억제하였고 (Duncan et al., 1996), 유방암 세포의 전이와 침윤을 억제 하여 간으로의 전이를 차단하는 것으로 밝혀졌다 (Zhang et al., 2012). 또한 cucurbitacin Q는 signal transducer and activator of transcription (STAT)‐3의 인산화를 저해하여 폐암 세포와 유방암 세포의 활성을 억제한다는 것이 보 고되었다 (Sun et al., 2005). 이와 같이 cucurbitacin B, E와 Q의 항종양 활성 및 항전이 작용과 그 작용 기전의 일부 가 밝혀져 있으나, 염증에 의해 유발되는 대장암에 있어 서 cucurbitacin I의 효과는 아직 알려지지 않고 있다. 이 에 본 연구에서는 염증성 대장암 실험 모델을 이용하여 cucurbitacin I의 항종양 활성 및 그 기전을 밝히고자 하 였다.
재료 및 방법 실험 재료
실험에 사용된 cucurbitacin I (Fig. 1A)는 Santa Cruz Biotechnology (Santa Cruz, CA, USA), dimethylsulfoxide (DMSO)는 Duchefa Biochemie (Haarlem, The Netherlands) 로 부터 구입하여 사용하였다. Cucurbitacin I는 2.5 mg/ml 의 농도로 DMSO에 희석하여 사용하였으며 ‐20℃에서 보 관하였다. Azoxymethane (AOM)은 Sigma‐Aldrich (St.
Louis, MO, USA), dextran sodium sulfate (DSS, MW 36,000‐50,000)는 MP Biomedicals (Solon, OH, USA)에서 구입하여 사용하였다.
실험 동물
본 실험에 사용한 동물은 대장암 동물실험모델에서 사 용되는 종인 BALB/c female 마우스 (RaonBio, Gyeonggj‐
do, Korea)로, 5주령의 체중이 16 ‐ 18 g인 것을 사용하였 다. 음수는 일주일에 2회 교환하여 공급하였고, 사료는 설치류용 표준사료를 충분히 공급하였다. 동물실험실은 대구가톨릭대학교 실험동물 사육실을 사용하였으며 온도 23 ± 2℃, 습도 45 ± 10%를 유지하고, 12시간 간격으로 light‐dark cycle을 유지하였다. 본 동물실험을 수행하기 전 대구가톨릭대학교 동물실험 윤리 위원회 (IACUC)의 심의 및 승인 (승인번호: CUD‐2012‐018)을 받았다.
대장암 유발 및 cucurbitacin I 투여
구입 후 7일간 안정기를 거친 BALB/c 마우스를 AOM/DSS를 투여하지 않은 음성대조군, AOM/DSS 만을 처치한 양성대조군, AOM/DSS를 처치하고 0.25 mg/kg 및 0.5 mg/kg cucurbitacin I 투여한 군의 총 4군으로 나누었 다 (Fig. 1B). AOM은 phosphate buffered saline (PBS)과 혼 합하여 10 mg/kg의 농도로 복강투여하고, 2주 후부터 8주 동안, 주 3회의 빈도로 총 24회에 걸쳐 cucurbitacin I를 복 강투여 하였다. AOM 투여 1주 후부터 DSS를 1주간 음수 로 공급시킨 후 2주간 휴식기를 가졌고, 또 다시 1주간 DSS를 음수로 공급시킨 후 2주간 휴식기를 가지는 방법 으로, 3회 반복 실시하였다. DSS를 공급하지 않는 기간에 는 일반 음용수를 공급하였다. 마우스에 공급할 DSS는 음수로 용해하여 조제하였으며, cucurbitacin I는 매 회 복 강투여 직전에 농도에 맞게 PBS로 희석하여 조제하였다.
총 10주간의 실험 기간 후 부검하였다.
(A)
Fig. 1. The chemical structure of cucurbitacin I and experimental design. (A) The chemical structure of cucurbitacin I. (B) BALB/c mice at 6 weeks of age received a single intraperitoneal injection of the mutagenic agent AOM (10 mg/kg body weight). Starting at day seven after the AOM application, 2% DSS was dissolved in the drinking water for three cycles of five days each with intermittent 14‐days intervals of regular drinking water. Vehicle (PBS) or cucurbitacin I was administered three times a week for 8 weeks. 10 weeks later, the mice were killed for analysis.
본 실험을 진행하는 기간 중 1주일에 한번씩 마우스의 체중을 측정하였다. 매 측정 시 오차를 최소화하기 위하 여 동일한 요일과 동일한 시간에 동일한 저울을 사용하 여 측정하였다.
대장의 길이 및 장기 무게 측정
마우스 부검 후 대장을 적출하여 맹장에서 항문까지의 길이를 측정하였고, 간과 비장의 장기 무게를 저울을 이 용하여 측정하였다. 그리고 대장 내 잔여 분뇨를 PBS을
사용하여 깨끗이 제거하고 세로로 절개하였다. 대장 내부 조직에 형성된 종양의 개수를 측정하였다.
대장 조직의 Western blotting 분석
적출한 대장 조직에 PRO‐PREP Protein Extraction Solution (iNtRON Biotechnology, Seongnam‐Si, Korea)을 넣어 homogenizer로 분쇄시킨 후 원심 분리 (13,000 rpm, 20분, 4℃)하여 단백질이 함유된 상층액을 얻었다. 정제 한 단백질 30 µg을 10% SDS‐polyacrylamide gel 전기 영 (B)
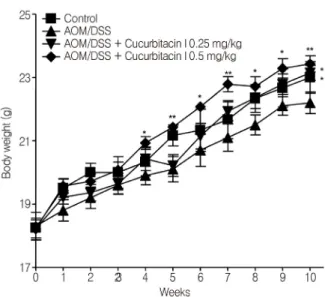
Fig. 2. Change in the body weights in BALB/c mice.
Body weight changes were monitored once a week for experiment period. The results are reported as mean ± SEM.
Significant differences in the AOM/DSS group, *P < 0.05 and **P < 0.01.
동으로 분리시킨 후 Bio‐Rad의 Electroblot system (Hercules, CA, USA)을 사용하여 PVDF membrane (Roche, Indianapolis, IN, USA)에 이동시켰다. 항체의 비특이적인 결합을 억제하기 위해 3% skim milk로 실온에서 1시간 동안 blocking한 후, 1차 항체인 iNOS (BD Bioscience, San Jose, CA, USA), COX‐2 (Cayman Chemical, Ann Arbor, MI, USA), TNF‐α와 IL‐6 (Santa Cruz Biotechnology)을 1:1000 희석하여 4℃에서 24시간 반응시켰다. 세척 후, horseradish peroxide (HRP)가 표지된 2차 항체를 실온에 서 1시간 반응 후 WEST‐ZOL plus Western Blot Detection System (iNtRON Biotechnology)을 이용하여 iNOS, COX‐
2, TNF‐α, IL‐6의 단백질 발현양을 관찰하였다. 검출된 단 백질 정량은 Davinch‐Chemi CAS‐400SM Western Imaging System (Davinch‐K, Seoul, Korea)과 Total Lab Software (Davinch‐K)를 이용하였다.
대장 조직의 병리학적 분석
적출한 대장을 10% formalin (Sigma‐Aldrich) 용액으로 3일 동안 고정 후 파라핀 블록을 제작하여 5 µm 두께로 각 조직표본을 박절하였다. 대장 조직은 hematoxylin과 eosin으로 염색하여 광학현미경으로 관찰하였다.
통계학적 분석
통계 처리는 GraphPad Prism 4.0 software (GraphPad Software Inc., San Diego, CA, USA)를 이용하여 표준편 차 (mean ± SEM)로 표기하였다. 대조군과 실험군의 유 의한 차이가 있는지 알아보기 위해 Student’s t‐test 분석 을 통해 P‐value < 0.05일 때 통계적으로 유의한 것으로 간주하였다.
결 과
Cucurbitacin I에 의한 염증성 대장암의 억제효과 실험 기간 동안 마우스의 체중의 증가는 AOM/DSS 만 을 처리한 양성 대조군이 가장 낮게 나타났으며, 그 외 다 른 군들은 모두 비슷한 양상의 체중 증가를 나타내었다.
Cucurbitacin I를 투여한 동물의 체중은 AOM/DSS 만을 처리한 양성 대조군에 비해 높은 것을 확인할 수 있었고 특히 0.5 mg/kg의 cucurbitacin I 투여군은 AOM/DSS 단독 처리군에 비해 AOM 투여 4주 후부터 유의적인 체중의 증가가 관찰되었다 (Fig. 2).
Cucurbitacin I 투여가 각 장기에 미치는 영향을 조사하 기 위해 해부 시 장기를 관찰한 결과, 특이적인 이상은 관 찰되지 않았다 (data no shown). 또한 각 군의 마우스에서 간 과 비장을 적출하여 그 무게를 측정한 결과 각 군간의 유의적인 차이는 관찰할 수 없었다 (Table 1).
대장의 변화를 관찰하기 위해 마우스의 대장을 적출하 여 길이를 측정하였다. AOM과 DSS로 대장암을 유발하 면 그 길이가 정상적인 대장길이 보다 짧게 나타나는 것 으로 알려져 있다 (Li et al., 2010). 각 군의 대장의 길이를 비교한 결과, 음성 대조군인 control 군이 16.07 ± 0.13 cm, 양성 대조군인 AOM/DSS 단독 처리군이 13.73 ± 0.18 cm 로 AOM/DSS 처리에 의해 대장길이가 유의적으로 감소 한 것을 확인할 수 있었다. AOM/DSS 처리에 의해 감소 한 대장의 길이가 0.5 mg/kg의 cucurbitacin I 투여에 의해 완화된 경향을 확인할 수 있었으나 (14.24 ± 0.35 cm), 통 계학적인 유의 차는 관찰할 수 없었다 (Fig. 3A).
다음은 AOM/DSS에 의해 유발된 대장 조직 내의 생성 된 종양의 수를 관찰하였다 (Fig. 3B, C). AOM/DSS에 의 해 유발된 대장 조직 내의 종양 개수는 26.20 ± 1.96개로 가장 많은 개수를 나타내었다. 0.25 mg/kg 및 0.5 mg/kg cucurbitacin I 투여군의 종양 개수는 각각 13.57 ± 1.74, 11.71 ± 1.95개로 AOM/DSS 처리군 보다 유의적으로 적 은 종양 개수가 관찰되었다.
Experiment group Body weight (g)
Liver Spleen
Relative weight (g/100 g body weight)
Control 23.00 ± 0.50* 4.36 ± 0.19 0.43 ± 0.03
AOM/DSS 22.20 ± 0.34 4.26 ± 0.07 0.47 ± 0.02
AOM/DSS
+ Cucurbitacin I 0.25 mg/kg 23.14 ± 0.24* 3.97 ± 0.10 0.44 ± 0.02 AOM/DSS
+ Cucurbitacin I 0.5 mg/kg 23.43 ± 0.25** 4.07 ± 0.05 0.51 ± 0.01
AOM/DSS induced colitis‐associated colon cancer mice treated with or without cucurbitacin I. The results are reported as mean ± SEM. Statistical significance is based on the difference when compared with the AOM/DSS group, *P < 0.05,
**P < 0.01.
Table 1. Body weight, relative liver and spleen weight in BALB/c mice.
(A) (B)
(C)
Fig. 3. Cucurbitacin I reduced tumor formation in BALB/c mice. Colon tissues were removed at ten weeks to determine (A) colon length, (B) the number of tumors. (C) Representative images from BLAB/c mice treated with or without cucurbitacin I administration. Arrows indicate tumors. The results are reported as mean ± SEM. Significant differences in the AOM/DSS group, ***P < 0.001.
Fig. 5. Cucurbitacin I inhibited AOM/DSS‐induced iNOS, TNF‐α and IL‐6 in mouse colon. (A) Western blots of colon lysates from the indicated groups for iNOS, COX‐2, TNF‐α and IL‐6. (B) The bands were quantified using image analysis software and their relative intensity was expressed as fold against the image of the AOM/DSS groups, *P < 0.05,
**P < 0.01, ***P < 0.001.
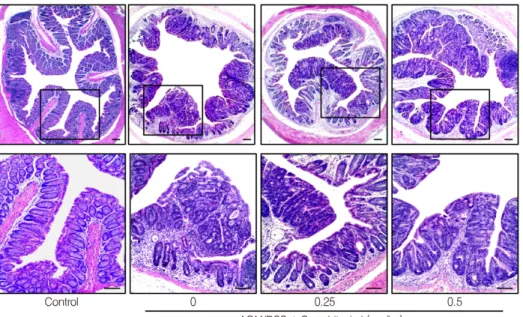
Fig. 4. Histological image of colon tissues in BALB/c mice. Representative hema- toxy-lin and eosin stained sections from AOM / DSS induced colitis‐associated colon cancer mice with or without cucurbitacin I administration.
Original magni-fication was 40x (upper panels) and 100x (lower panels). Scale bars represent 100 μm.
Cucurbitacin I에 의한 대장 조직의 병리학적 관찰 대장 조직을 병리학적으로 관찰한 사진을 Fig. 4에 나 타내었다. 동물 모델을 이용한 대장암 억제 실험에서는 대장 조직이 비정상적으로 변형되거나 암종 또는 종양이 생기는 것을 통하여 대장암이 유발되었음을 확인하는 것 이 일반적이다 (De Robertis et al., 2011). AOM/DSS를 처 리하여 대장암을 유발시킨 마우스의 대장 조직은 정상적 인 장 조직의 장샘 (intestinal glands)세포들이 사라지고 점막의 종양을 나타내며, 조직 내 암세포들이 침윤한 것 을 관찰 할 수 있었다. 또한 염증세포들이 점막 층에 축적 되고 상피세포 층의 구조가 파괴된 것을 된 확인할 수 있 었다. Cucurbitacin I 처리군 (0.5 mg/kg)의 대장 조직 내 상피세포의 모양은 정상 대장 조직과 유사한 모양을 형 성하고 있었고 대장조직의 파괴 정도가 상당히 완화된 것을 확인할 수 있었다.
Cucurbitacin I에 의한 iNOS, COX‐2, TNF‐α와 IL‐
6의 단백질 발현 변화
Cucurbitacin I 투여에 의한 대장암 발생의 억제가 대장에 서의 염증 반응의 억제에 기인하는 것인지를 확인하기 위 해 대장 조직에서 얻은 단백질을 이용하여 iNOS, COX‐2, TNF‐α 및 IL‐6의 단백질 발현양을 조사하였다 (Fig. 5).
AOM/DSS 처리에 의해 대장 조직의 iNOS, COX‐2, TNF‐α 및 IL‐6의 단백질 발현양이 현저히 증가하였으나 cucurbitacin I의 처리에 인해 iNOS, TNF‐α와 IL‐6의 단백질 발현양이 감소함을 확인할 수 있었다 (Fig. 5A, B).
(A)
(B)
Cucurbitacin I의 처리에 의한 대장 조직 내의 COX‐2 단백질 의 발현 변화는 관찰할 수 없었다. 위의 결과는 cucurbitacin I의 투여가 염증 반응을 유발하는 iNOS 및 염증성 사이토 카인의 단백질 발현을 억제함으로써 염증 반응에 의해 촉 진되는 대장암의 유발을 억제하는 것을 시사하고 있다.
고 찰
대장암은 서구사회에서 흔히 발병하는 악성종양의 하 나로 미국과 서구 유럽에서는 모든 악성종양의 약 10%를 차지하며 미국의 경우 암에 의한 사망률 2위에 해당된다 (Jemal et al., 2003). 서구 사회에서는 지난 40년간 대장암 발생이 약간 감소하는 추세를 보이고 있으나 국내에서는 꾸준한 증가추세를 보이고 있어, 1999년 10만 명당 21.1 명에 불과한 대장암 발병률이 2010년 51.7명으로 급격하 게 증가되었다. 대장암 발병률의 증가의 중요 원인으로 서구화된 식습관을 들 수 있으나, 만성염증 질환인 염증 성 장질환 또한 그 주요원인의 하나로 간주된다 (Tanaka, 2012).
염증 반응은 인체에 유해한 자극의 노출로부터 대장 조직의 세포 손상을 일으키는 원인을 제거하거나 희석하 기 위한 생체 반응으로, 대장 상피 세포에 병원균과 같은 외부 자극이 전해지면 면역세포들이 활성화되어 NO, PGE2, 사이토카인 등의 염증 유발 물질을 다량으로 분비 하여 생체를 보호하게 된다.
우리나라에서의 대장암의 발생빈도는 지속적으로 증 가 추세이지만 이를 효과적으로 치료할 방법의 개발은 아직 미비하다고 할 수 있다. 현재 사용되고 있는 항암제 들은 식욕부진, 과민반응, 구토 등과 같은 다양하고 심각 한 부작용을 동반하고 있어 부작용이 비교적 적고 암세 포만을 표적으로 하는 특이적인 항암제의 개발이 시급하 다. 이를 보완하기 위해 자연 상태에 존재하는 식물체로 부터 분리된 생리활성 성분인 phytochemical을 이용한 연 구들이 활발하게 진행되고 있다. 본 연구에서는 호박이나 표주박과 같은 박과 식물들에서 해충으로부터 자신을 방 어하기 위해 만들어내는 phytochemical인 cucurbitacin I를 이용하여 대장암에서의 효과를 조사하였다
Cucurbitacin I를 처리한 결과, 1) AOM/DSS를 처리로 인한 체중감소를 완화 시키고, 2) 염증 및 암화 현상에 의 한 대장 길이의 감소를 억제하였으며, 3) 대장조직 내의 염증 반응을 매개하는 효소 및 염증성 사이토카인의 발 현을 억제하여, 결과적으로 대장 조직내의 종양의 수를
감소시켰다.
과량 생성된 NO는 사람이나 설치류의 대장암 발생 촉 진에 관여하는 것으로 알려져 있다 (Youn et al., 2009). 또 한 iNOS 유전자가 결핍된 마우스는 AOM/DSS를 이용한 대장암의 유발이 정상마우스에 비해 현저하게 억제된다 는 연구 결과가 보고되었다 (Choi et al., 2006). 본 연구에 서 cucurbitacin I의 처리가 대장조직에서의 iNOS의 발현 억제를 유도하였다. 따라서 cucurbitacin I에 의한 대장암 의 발생을 억제하는 하나의 기전으로 iNOS 발현 억제에 의한 NO의 생성 억제를 들 수 있을 것이다. 대장암 환자 의 혈액과 병변 조직에서 높은 농도의 IL‐6가 관찰되는 것이 밝혀져 있다 (Atreya and Neurath, 2005). Grivennikov 등의 연구에 의하면 IL‐6 유전자 결핍 마우스에 AOM/DSS 모델을 적용하여 대장암을 유발한 결과, 정상 마우스에 비해 대장암의 발생이 현저하게 저하됨이 밝혀 졌다 (Grivennikov et al., 2009). 이와 같은 결과는 IL‐6에 의해 유도되는 STAT3의 활성이 대장암세포의 증식과 세 포사멸의 억제를 조절에 중요한 역할을 하고 있음을 제 시하고 있다 (Hodge et al., 2005). 본 연구에서 STAT3의 활성 정도는 관찰하지 못하였으나, cucurbitacin I의 처리 에 의해 대장 조직에서의 IL‐6의 발현 감소가 STAT‐3의 활성을 억제하여 대장암세포의 증식 억제와 세포사멸을 촉진을 유도하여 대장암의 발생을 억제하였을 것으로 사 료된다.
Cucurbitacin I의 처리가 대장 조직 내에서 iNOS 및 염 증성 사이토카인의 발현을 현저하게 감소시킨 것과는 대 조적으로 COX‐2의 단백질 수준에서의 발현 억제는 관찰 되지 않았다. COX는 arachidonic acid 로부터 PG가 합성 되는 첫 번째 단계를 촉매 하는 효소로, COX‐2의 과 발현 은 암세포의 세포사멸을 억제시키고 침윤성을 증가시켜 암화 및 전이를 촉진한다고 알려져 있다 (Greengough et al., 2009). 대장암을 비롯하여 위암, 폐암, 전립선암, 유방 암 등의 대부분의 암 조직에서 COX‐2의 과 발현이 보고 되었다 (Hida et al., 1998). 본 연구에서는 cucurbitacin I 투 여에 의한 대장조직 내에서의 COX‐2 발현의 억제는 관찰 할 수 없었으나, iNOS 및 염증성 사이토카인의 발현억제 가 대장 조직내의 염증 반응을 완화시킴으로 염증으로 유도되는 대장암의 발생을 억제하였을 것으로 사료된다.
감사의 글
본 연구는 대구가톨릭대학교 학술연구비 지원으로 수 행하였으므로 이에 감사 드립니다 (과제번호: 20131028).
REFERENCES
Alghasham AA. Cucurbitacins – a promising target for cancer therapy. Int J Health Sci (Qassim). 2013. 7: 77‐89.
Atreya R, Neurath MF. Involvement of IL‐6 in the pathogenesis of inflammatory bowel disease and colon cancer. Clin Rev Allergy Immunol. 2005. 28: 187‐196.
Chen X, Bao J, Guo J, Ding Q, Huang M, Wang Y. Biological activities and potential molecular targets of cucurbitacins:
a focus on cancer. Anticancer Drugs. 2012. 23: 777‐787.
Choi MN, Jo HY, Han BS, Jang DD, Kim JD, Nam SY, Kim YB, Lee BJ, Yun YW, Ahn BW. Supression of azoxymethane‐induced colorectal tumor in iNOS‐/‐ C57BL/6 mice. Lab Anim Res. 2006. 22: 135‐138.
De Robertis M, Massi E, Poeta ML, Carotti S, Morini S, Cecchetelli L, Signori E, Fazio VM. The AOM/DSS murine model for the study of colon carcinogenesis: From pathways to diagnosis and therapy studies. J Carcinog.
2011. 10: 9.
Duangmano S, Sae‐Lim P, Suksamrarn A, Patmasiriwat P, Domann FE. Cucurbitacin B causes increased radiation sensitivity of human breast cancer cells via G2/M cell cycle arrest. J Oncol. 2012. 2012: 601‐682.
Duncan KL, Ducan MD, Alley MC, Sauscille EA.
Cucurbitacin E‐induced disruption of the actin and vimentin cytoskeleton in prostate carcinoma cells. Biochem Pharmacol. 1996. 52: 1553‐1560.
Gillen CD, Walmsley RS, Zack M, Adami HO. Ulcerative colitis and Crohn’s disease: a comparison of the colorectal cancer risk in extensive colitis. Gut. 1994. 35: 1590‐1592.
Greengough A, Samrtt HJ, Moore AE, Roverts HR, Willians AC, Paraskeva C, Kaidi A. The COX‐2/PGE2 pathway: key roles in the hallmarks of cancer and adaptation to the tumor microenvironment. Carcinogenesis. 2009. 30: 377‐386.
Grivennikov S, Karin E, Terzic J, Mucida D, Yu GY, Vallabhapurapu S, Scheller J, Rose‐John S, Cheroutre H, Eckmann L, Karin M. IL‐6 and Stat3 are required for survival of intestinal epithelial cells and development of colitis‐associated cancer. Cancer Cell. 2009. 15: 103‐113.
Hale LP, Greer PK. A novel murine model of inflammatory bowel disease and inflammation‐associated colon cancer with ulcerative colitis‐like features. PLoS One. 2012. 7:
e41797.
Hida T, Yatabe Y, Achiwa H, Muramatsu H, Kozaki K,
Nakamura S, Ogawa M, Mitsudomi T, Sugiura T, Takahashi T. Increase expression of cyclooxygenase 2 occurs frequently in human lung cancer, specifically in adenocarcinomas. Cancer Res. 1998. 58: 3761‐3764.
Hodge DR, Hurt EM, Farrar WL. The role of IL‐6 and STAT3 in inflammation and cancer. Eur J Cancer. 2005. 41: 2502‐ 2512.
Jemal A, Murray T, Samuels A, Ghafoor A. Ward E, Thun MJ. Cancer statistics, 2003. CA cancer J Clin. 2003. 53:
5‐26.
Kim YH, Kwon HS, Kim DH, Shin EK, Kang YH, Park JH, Shin HK, Kim JK. 3, 3'‐diindolylmethane attenuates colonic inflammation and tumorigenesis in mice. Inflamm Bowel Dis. 2009. 15: 1164‐1173.
Korea Central Cancer Registry, Ministry of Health & Welfare, National Cancer Center. Annual report of cancer statistics in Korea in 2010. Available at http://www.cancer.go.kr/
ncic/cics_g02/cics_g027/1389370_6095.html
Li H, Wu WK, Li ZJ, Chan KM, Wong CC, Ye CG, Sung JJ, Cho CH, Wang M. 2, 3’, 4, 4’, 5’‐Pentamethoxy‐trans‐ stilbene, a resveratrol derivative, inhibits colitis‐associated colorectal carcinogenesis in mice. Br J Pharmacol. 2010.
160: 1352‐1361.
McCartney‐Francis N, Allen JB, Mizel DE, Albina JE, Xie QW, Nathan CF, Wahl SM. Suppression of arthritis by an inhibitor of nitric oxide synthase. J Exp Med. 1998. 178:
749‐754.
Popivanova BK, Kitamura K, Wu Y, Kondo T, Kagaya T, Kaneko S, Oshima M, Fujii C, Mukaida N. Blocking TNF‐ alpha in mice reduces colorectal carcinogenesis associated with chronic colitis. J Clin Invest. 2008. 118: 560‐570.
Sandborn WJ, Hanauer SB. Antitumor necrosis factor therapy for inflammatory bowel disease: a review of agents, pharmacology, clinical results, and safety. Inflamm Bowel Dis. 1999. 5: 119‐133.
Sarkar D, Saha P, Gamre S, Bhattacharjee S, Hariharan C, Ganguly S, Sen R, Mandal G, Chattopadhyay S, Majumdar S, Chatterjee M. Anti‐inflammatory effect of allylpyrocatechol in LPS‐induced macrophages is mediated by suppression of iNOS and COX‐2 via the NF‐kappaB pathway. Int Immunopharmacol. 2008. 8: 1264‐1271.
Sun J, Chu YF, Wu X, Liu RH. Antioxidant and antiproliferative activities of common fruits. J Agric Food Chem. 2002. 50: 7449‐7454.
Sun J, Blskovich MA, Jove R, Livingston SK, Coppola D,
Sebti SM. Cucurbitacin I Q: a selective STAT3 activation inhibitor with potent antitumor activity. Oncogene. 2005.
24: 3236‐3245.
Tanaka T. Development of an inflammation‐associated colorectal cancer model and its application for research on carcinogenesis and chemoprevention. Int J Inflam. 2012.
2012: 658‐786.
Thoennissen NH, Iwanski GB, Doan NB, Okamoto R, Lin P, Abbassi S, Song JH, Yin D, Toh M, Xie WD, Said JW, Koeffler HP. Cucurbitacin B induces apoptosis by inhibition of the JAK/STAT pathway and potentiates antiproliferative effects of gemcitabine on pancreatic cancer cells. Cancer Res. 2009. 69: 5876‐5884.
Tracey KJ. The inflammatory reflex. Nature. 2002. 420: 853‐
859.
Youn J, Lee JS, Na HK, Kundu JK, Surh YJ. Resveratrol and piceatannol inhibit iNOS expression and NF‐kappaB activation in dextran sulfate sodium‐induced mouse colitis.
Nutr Cancer. 2009. 61: 847‐854.
Yun HY, Dawson VL, Dawson TM. Neurobiology of nitric oxide. Crit Rev Neurobiol. 1996. 10: 291‐316.
Zhang R. Induction of inducible nitric oxide synthase: a protective mechanism in colitis‐induced adenocarcinoma.
Carcinogenesis. 2007. 28: 1120‐1130.
Zhang T, Li J, Dong Y, Zhai D, Lai L, Dai F, Deng H, Chen Y, Liu M, Yi Z. Cucurbitacin E inhibits breast tumor metastasis by suppressing cell migration and invasion.
Breast Cancer Res Treat. 2012. 135: 445‐458.