Role of Nox4 in Neuronal Differentiation of Mouse Subventricular Zone Neural Stem Cells
Ki-Youb Park
1*, Yerin Na
1and Man Su Kim
21
Korea Science Academy of KAIST 105-47 Baegyanggwanmun-ro, Busanjin-Gu, Busan 614-100, Korea
2
전체 글
1
2
수치
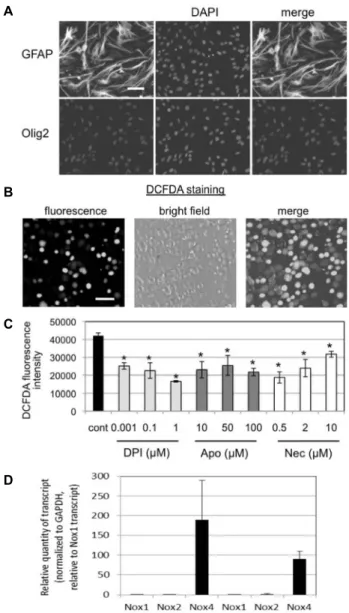
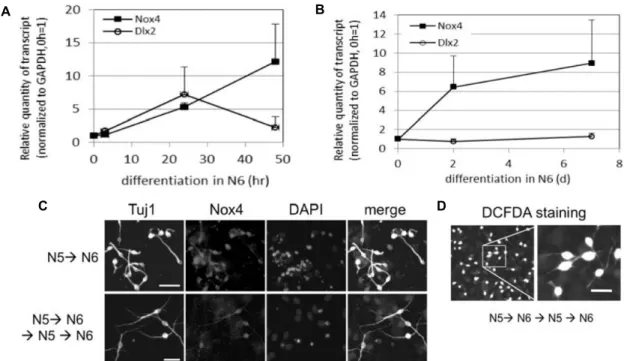
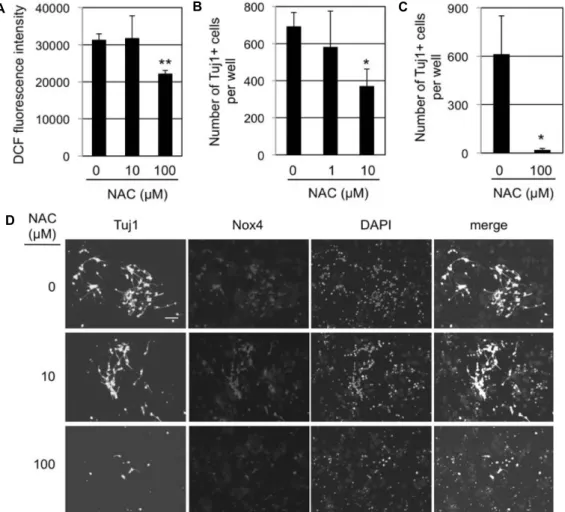
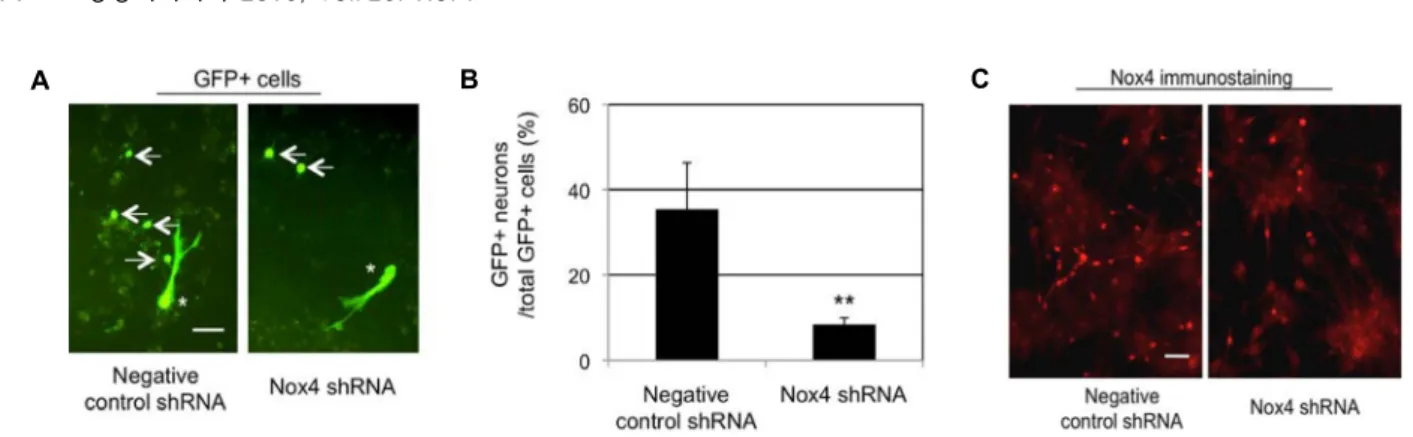
관련 문서
In Jangsu stream, blue-green algae were temporarily increased by 59% due to the eutrophication and reduction of water level in July, but no toxins
We found that FoxO1 is constantly increased in MCF-7/ADR, adriamycin-resistant breast cancer cells, and FoxO1 has a critical role in the MDR1 gene expression (16).. There is
Approved clinical use of bone marrow stem cells for myocardial infarction treatment... Cardiac
In this study, we found that Pin1 plays a pivotal role in insulin- induced AP-1 activation through its interaction with p70S6K and prolongs activity of
In this context, the present study found that the emphasis of art education was changing from expression-oriented practical classes to systematic
A Study on the Evaluation of Space Recognition Space Recognition Space Recognition in the Space Recognition in the in the in the Concourse of Hospital using the
Meterials and Methods: Our study involved three stem cell sources-subacromial bursal tissue, bone marrow, and umbilical cord-derived stem cells. Under consent we
Recent stem cell studies have reported that cultured hematopoietic stem cells (HSCs) are reactivated through fetal hemoglobin expression by treatment with
