53 http://dx.doi.org/10.7850/jkso.2014.19.1.53
Free Access
광양만에서 춘계와 하계 영양염류 첨가가 식물플랑크톤군집의 성장에 미치는 영향
배시우1,2·김동선3·최현우4·김영옥1·문창호2·백승호1,*
1
한국해양과학기술원 남해특성연구부
2
부경대학교 해양학과
3
한국해양과학기술원 해양순환 기후연구부
4
한국해양과학기술원 해양과학데이터센터
The Influence of Nutrients Addition on Phytoplankton Communities Between Spring and Summer Season in Gwangyang Bay, Korea
S I W OO B AE
1,2, D ONGSEON K IM
3, H YUN -W OO C HOI
4, Y OUNG O K K IM
1, C HANG H O M OON
2AND S EUNG H O B AEK
1,*
1
Korea Institute of Ocean Science and Technology / South Sea Institute, Geoje 656-830, Korea
2
Departmaent of Oceanography, Pukyong National University, Busan 608-737, Korea
3
Ocean Circulation and Climate Research, KIOST Ansan 425-600, Korea
4
Ocean Data & Cilimate Research Department, KIOST Ansan 425-600, Korea
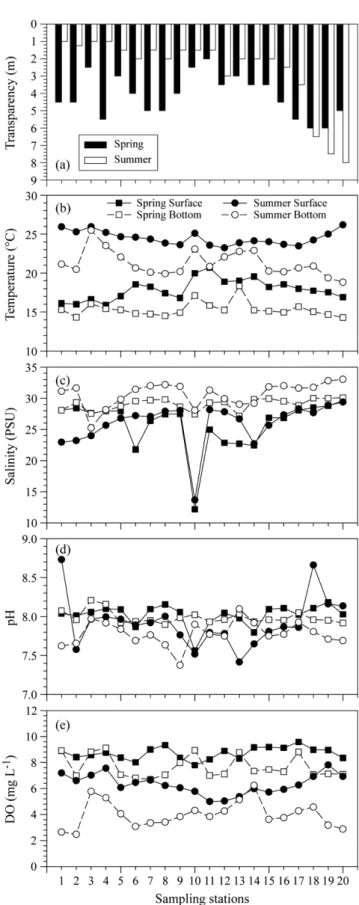
춘계와 하계 광양만에서 식물플랑크톤 군집구조와 그들의 성장에 영향을 미치는 영양염제한 특성을 파악하기 위 해서 만내외측의 20개 정점에서 생물학적 요인과 무생물학적 요인을 조사했다. 또한 식물플랑크톤 군집에 대한 영 양염 첨가 효과를 알아보기 위해서 실험실에서 현장 20개 정점의 표층수를 이용하여 생물검정실험을 수행하였다.
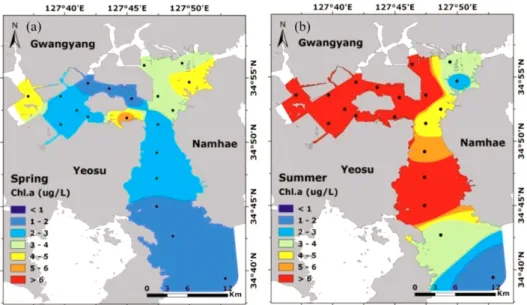
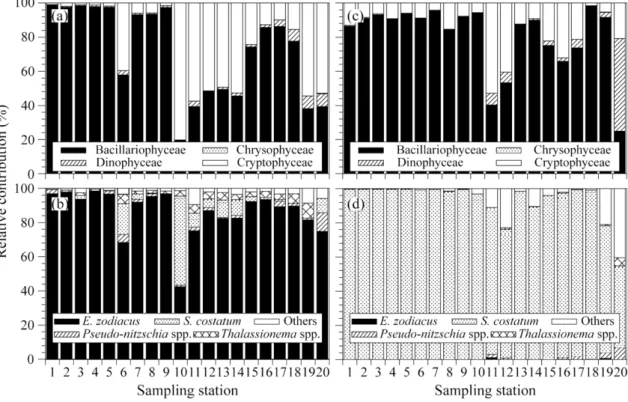
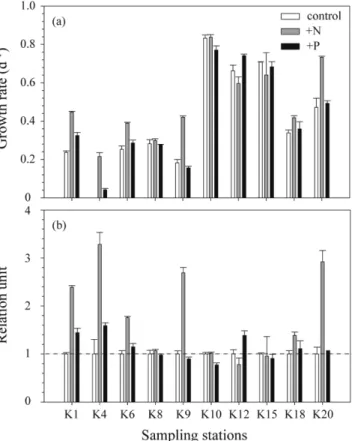
전체 식물플랑크톤 군집의 90%이상을 규조류가 차지하였다. 이들 규조류중 Eucampia zodiacus와 Skeletonema costatum-like 종이 춘계와 하계에 각각 우점하였다. E. zodiacus와 S. costatum-like 규조류의 개체군 밀도가 춘계와 하계에 높은 밀도를 유지하게 된 이유를 간단히 설명하면, E. zodiacus 의 성장은 춘계 투명도가 높게 나타나 유광 층내 광량의 증가가 원인으로 생각된다. 즉 유광층내 광량의 증가는 E. zodiacus의 개체수를 폭발적으로 증가시킬 수 있는 방아쇠 역할을 한 것으로 판단된다. 하계에 S. costatum-like이 전해역에서 우점한 이유는, 섬진강 담수 유 입에 의한 낮은 염분과 함께 공급된 다량의 영양염은 그들 생물의 증식에 중요한 bottom-up 효과를 보였다고 판단 된다. 실험실의 생물검정실험에서는 비록 내만(정점 8)과 외해(정점 20)에서 식물플랑크톤 군집의 최대 성장율은 유 사하였지만, 인산염에 대한 반포화계수(K
s)는 내만정점보다 약간 낮았다. 상대적으로 낮은 영양염농도에 적응한 세 포는 낮은 인산염농도에서 충분히 성장할 수 있고, 다른 미세조류에 비하여 낮은 영양염농도의 조건에서 경쟁의 우 위를 차지 할 것이다. 특히, 하계의 N영양염 첨가군의 효율은 대조군과 P영양염 첨가군에 비해서 높았다. 이는 광 양만에서 하계에 N영양염의 공급이 섬진강을 통하여 계속적으로 유입되지만, 빠른 식물플랑크톤의 증식으로 인하 여 N영양염 제한이 일어날 수 있다는 것을 시사할 수 있다. 반면, 규산염은 식물플랑크톤의 성장에 영향을 미치는 제한인자로 나타나지 않았고, 규조류의 분해로 인하여 Si의 재순환과 담수로부터 공급된 높은 규산염농도는 광양만 에서 규조류 생태계를 유지할수 있는 유리한 조건이라 생각된다.
In order to estimate the effect of nutrients addition for phytoplankton growth and community compositons in spring and summer season, we investigated the abiotic and biotic factors of surface and bottom waters at 20 sta- tions of inner and offshore areas in Gwangyang Bay, Korea. Nutrient additional experiments were also con- ducted to identify any additional nutrient effects on phytoplankton assemblage using the surface water for the assay. Bacillariophyceae occupied more than 90% of total phytoplankton assembleges. Of these, diatom Eucampia
Received November 22, 2013; Revised February 5, 2014; Accepted February 7, 2014