Yeungnam Univ J Med 2017;34(2):149-160
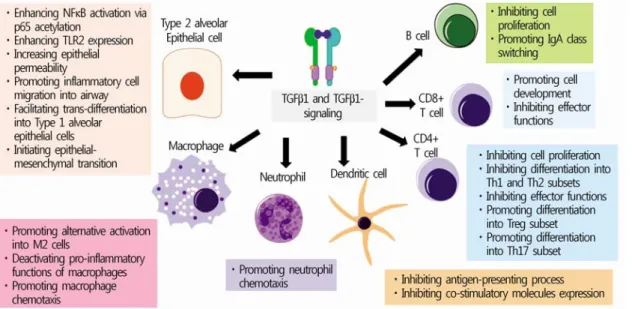
Role of the transforming growth factor (TGF)-β1 and TGF-β1 signaling pathway on the pathophysiology of respiratory pneumococcal infections
12
0
0
전체 글
(2)
(3)
(4)
(5)
(6)
(7)
(8)
(9)
(10)
(11)
(12)
수치

관련 문서
