INTRODUCTION
Radiation-induced late injury to normal tissue is one of the primary areas of radiation biology research (Zhao et al. 2007). Radiation-induced late normal tissue injury has been viewed as a combination of loss of normal cellular function as well as an orchestrated response to injury that involves interactions between multiple cell types within a particular organ. Robins and Zhao suggest that the development and progression of radiation-induced late effects are driven by an acute and chronic oxidative stress (Robbins and Zhao 2004).
What is the biological basis for the effects of ionizing radiation on the aging? The question has remained virtually unanswered due to uncertainty concerning radiation’s ability
to accelerate the normal aging process. The effect of ionizing radiation on the aging has been vigorously studied from late 1950s (Mewissen et al. 1957; Upton et al. 1960). One of main aging mechanistic theories of aging with the health effects of ionizing radiation (IR) has been free radical damage and oxidative stress (Richardson 2009) since IR is known to generate reactive oxygen species (ROS). ROS are the main sources of free radical damage. Irradiating eukaryotic cells results in a rapid burst of ROS generated primarily as a result of ionizing of water molecules in vitro (Riley 1994) as well as in vivo. Due to the transient nature of the ROS species generated, direct measurements are extremely challenging.
Currently IR has become an inevitable health concerns from natural source like space travel and from artificial sources like medical therapies. The brain is especially vulner-able to oxidative damage as a result of its high oxygen con-sumption rate, its abundant lipid content, and the relative paucity of antioxidant enzymes compared to other tissues
─ ─ 203 ──
No Late Effect of Ionizing Radiation on the Aging-Related
Oxidative Changes in the Mouse Brain
Beom-Su Jang*, Seolwha Kim, Uhee Jung and Sung-Kee Jo
Radiation Biotechnology Research Division, Advanced Radiation Technology Institute, Korea Atomic Energy Research Institute, Jeongeup 580-185, Korea
Abstract -- Radiation-induced late injury to normal tissue is a primary area of radiation biology research. The present study was undertaken to investigate whether the late effect of the ionizing radiation appears as an age-related oxidative status in the brain. Three groups of 4-month old C57BL/6 mice that were exposed to 137Cs γγ-rays at a single dose (5 Gy) or fractionated doses (1 Gy ×
×5 times, or 0.2 Gy××25 times) at 2 months old were investigated for the oxidative status of their brains with both young (2-month) and old (24-month) mice. A significant ( p⁄⁄0.05) decrease in superoxide dismutase (SOD) activity was observed in old mice brains compared with that of the young mice. malondialdehyde (MDA) content was significantly ( p⁄⁄0.05) increased in the old mice brain. However, any significant difference in SOD activity and MDA contents of the irradiated brain was not observed compared to age-matched control group mice. SOD activity and MDA content were observed within good parameters of brain aging and there were no late effects on the age-related oxidative level in the γγ-ray irradiated mice brains.
Key words : Late effect, Ionizing radiation, Brain oxidative change, Aging
* Corresponding authors: Beom-Su Jang, Tel. +82-63-570-3228, Fax. +82-63-570-3229, E-mail. [email protected]
(Leutner et al. 2001). Cranial radiation therapy is crucial to successful treatment of many primary brain tumors, cancers metastatic to the brain, central nervous system involvement of leukemia/lymphoma, and head and neck cancers.
ROS attack macromolecules causing oxidative stress, a process involved in the etiology of many diseases, and even at low levels in some organs such as the brain probably con-tributes to aging (Floyd 1999). Oxidative stress and ROS have been proposed to be major cause of aging and other age-related conditions such as Alzheimer’s disease and Parkin-son’s disease (Athar 2002). The overproduction of ROS or decrease in antioxidant enzymes in the brain can cause aging-related oxidative damage through macromolecules peroxida-tion (Stadtman 2006). Lipid peroxides readily decompose to librate highly reactive carbonyl fragments, the most promi-nent being malondialdehyde (MDA). On the other hand, the mammalian body has endogenous enzymatic defenses to fight oxidative stress such as superoxide dismutase (SOD), catalase, glutathione peroxidase, glutathione S-transferase etc.
Although it is generally acknowledged that antioxidant defenses decline with age, there are many conflicting reports on age-associated alterations in oxidative damage and antiox-idant enzyme activity in brain (Hiramatsu et al. 1992; Mo et al. 1995; Kurokawa et al. 2001; Leutner et al. 2001). These inconsistent results may arise from differences in strains or ages of the animals investigated or from the variety of exper-imental procedures used (Benzi and Moretti 1995).
Until now, there was a lack of experimental data about changes in the oxidative status on the brain of irradiated animals in comparison with aging. Since SOD and MDA are well studied parameters for the oxidative status in brain aging, we observed these parameters in the irradiated mice brain to compare with natural aging. For mice, 6.5 Gy is the median lethal dose (LD50) for 30 days and 5 Gy is accepted as sub-lethal high dose. In this study, the late effects of 5 Gy as sub-lethal high dose of ionizing radiation on the SOD and MDA in brain was evaluated with various irradiation protocols from single high dose to multiple low dose.
MATERIALS AND METHODS
1. Animals
Six-week-old C57BL/6 female mice were purchased from
Orient Bio Co., Ltd. (Charles River Technology, Korea) and were aged at the animal facility of the Advanced Radiation Technology Institute of Korea Atomic Energy Research Institute (KAERI). They were housed in 5 per cages under specific pathogen-free conditions at 23±3�C and 50±10% relative humidity, and were provided with nutritional chow (5 L79, PMI Nutrition LLC, USA). The mice were given free access to water and a standard diet. Animal experiment protocol was approved by the Animal Care and Use Commit-tee of KAERI and conducted in accordance with the princi-ples enunciated in the ‘Animal Care Act’ and ‘Laboratory Animal Law’.
2. Irradiation
For evaluation of late effect of ionizing radiation, 2-month-old mice were divided into four groups with twelve mice in each group. The mice were exposed to whole-body γ-irradia-tion in a plastic chamber with 137Cs γ-rays of Gamma Cell 40 Exactor (Nordion International Inc., Canada) without physical restraint or anesthesia. Single irradiated mice were given 5 Gy (1.1 Gy min-1). Fractionated irradiation groups were given 5 Gy cumulative doses for 5 weeks; (1) 1 Gy×5 (once a week for 5 weeks), (2) 0.2 Gy (0.11 Gy min-1)×25 (five times a week for 5 weeks), respectively. We used atten-uator to minimize the variation for 0.2 Gy group because the exposure time for 0.2 Gy at the dose rate as 1.1 Gy min-1 was quite short (about 11 sec). After exposing to whole-body γ-irradiation, the mice were monitored daily for the develop-ment of symptoms of radiation sickness and mortality. At 2 months after the beginning of the exposure, animals were sacrificed and all analyses were performed. In addition, the young (2-month) and old (24-month) group with 10 mice were studied to evaluate the aging effects on the oxidative status in the brain.
3. SOD and MDA assay
To address the changes of the oxidative status of the brain by aging and exposure to ionizing radiation, SOD activity and MDA level were investigated. Mice were sacrificed with decapitation and the brain was quickly removed on ice. And then it was homogenized (10%) in cold saline, centrifug-ed at 4,000×g at 4�C for 20 min. SOD activity was deter-mined by measuring the inhibition of dismutation of super-oxide radical compared with standard SOD enzymes and
the results were expressed as units per mg of protein (Peskin and Winteerbourn 2000). The level of lipid peroxidation in brain homogenate was indicated by the content of the MDA. Thiobarbituric acid reaction (TBAR) method was used to determine the MDA level on the basis of MDA reacting with thiobarbituric acid (detected at 532 nm) (Ohkawa et al. 1979). MDA content was expressed as nmol per mg of protein. The protein concentrations were measured by the bicinchoni-nic acid protein quantification method.
4. Statistical analysis
Statistical analysis was done using one way analysis of variance and inter group comparisons were made using Turkey’s multiple comparison test. The values are expres-sed as mean±SD. P value ⁄0.05 was considered as signi-ficant.
RESULTS
1. Effects of aging & γγ-irradiation on the SOD activity in the brain of mice
We investigated the changes of the SOD activity in the brain by aging and radiation exposure in mice (Fig. 1). A sig-nificant decrease ( p⁄0.05) in SOD activity was observed in the old group compared to the young group (12.2±1.4 vs 10.5±2.0 U mg-1protein). The SOD activity of 5 Gy group was the lowest (10.5±2.6 U mg-1protein) and that of 0.2 Gy×25 group was highest (13.5±1.7). There are no signifi-cant differences in SOD activity between all IR exposed groups and the age-matched control group.
2. Effects of aging and irradiation on the MDA contents in the brain
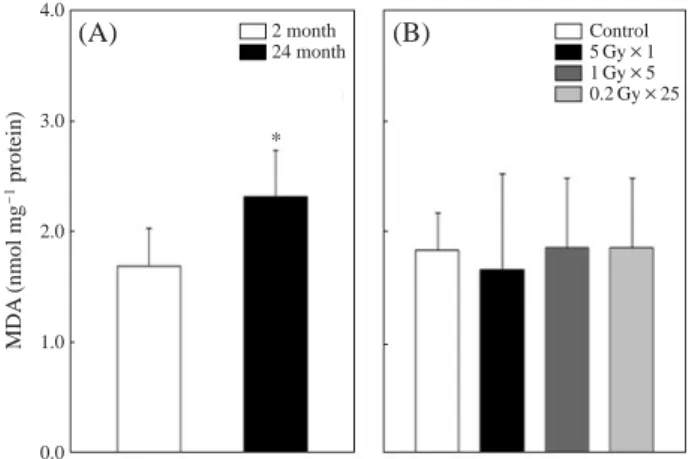
Fig. 2 shows an MDA level in the brain of normal and γ-irradiated mice. MDA level in the brains of the old group (24 month) was significantly ( p⁄0.05) higher than that of the young group (2 month) (1.7±0.3 vs 2.3±0.4 nmol mg-1 protein). The mean value of MDA levels of each irradiation group was similar to the control group within the range of between 1.7 and 1.9 nmol mg-1protein. There are no signifi-cant difference in MDA level of brain between all IR exposed groups and the age-matched control group.
DISCUSSION
Brains are generally liable to oxidative stress. In the litera-ture, a decrease of SOD and an increase of MDA with age is a consistent finding in most tissues (Navarro and Boveris 2004). And these changes are paralleled by increased DNA and protein oxidation and mitochondrial oxidative changes in several animals as well as in humans (Ando et al. 1990; Smith et al. 1991).
The chronic oxidative stress has been observed in the irra-Fig. 1. The effect of aging and irradiation on the Superoxide
dismu-tase (SOD) activity in the brain of the young and old (2-, 24-month) (A) and irradiated (4-month old) groups (B) of C57BL/ 6 mice. Mice were exposed to the ionizing radiation at single (5 Gy) or fractionated (1 Gy×5, 0.2 Gy×25) at 2 month old.
Values are expressed as mean±SD (n==10-12). Asterisk (*)
indicates p⁄0.05 vs young group.
Fig. 2. The effect of aging and irradiation on the Malondialdehyde
(MDA) contents in the brain of the young and old (2-, 24-month) (A) and irradiated (4-month old) groups (B) of C57BL/ 6 mice. Mice were exposed to the ionizing radiation at single (5 Gy) or fractionated (1 Gy×5, 0.2 Gy×25) at 2 month old.
Values are expressed as meanæSD (n==10-12). Asterisk (*)
indicates p⁄0.05 vs young group.
SOD (U mg -1protein) 20 15 10 5 0 2 month 24 month Control 5 Gy×1 1 Gy×5 0.2 Gy×25 * (A) (B) MDA (nmol mg -1protein) 4.0 3.0 2.0 1.0 0.0 * 2 month 24 month Control 5 Gy×1 1 Gy×5 0.2 Gy×25 (A) (B)
diated lungs, kidneys of mice 15 to 24 weeks after irradiation as a late effect (Kang et al. 2003; Moulder 2003). Limoli et al. observed significant increase in lipid peroxidation in the adult male mouse hippocampus 2 weeks after brain irradia-tion with a single high dose of 10 Gy (Limoli et al. 2004). To find out whether the late effect was as oxidative stress in the irradiated brain, we investigated the changes of SOD and MDA level in irradiated brains.
In the present study, SOD activity and MDA levels in the irradiated mice brains were not significantly changed at 2 months after irradiation, suggesting there were no late effects of irradiation on the oxidative status of irradiated brains regardless of irradiation protocols (neither single high dose nor multiple low dose). The effect of radiation on the oxida-tive status in the brain was not sustained 2 month later. IR effect on oxidative status in brain seems reversible or tem-porary. Additionally to confirm no late effect of irradiation on SOD and MDA in the brain, we investigated for an extend-ed period up to 6 months after irradiation. There were no significant differences between the control group and the irradiation groups (Data not shown). Since persistent oxida-tive changes in the brain may lead to the neuroinflammation and neurodegenerative progress, this finding is supportive data for radiation therapy.
In the present study significant decrease of SOD activity and increase of MDA level in the old mice brains were observed, suggesting an involvement of oxygen free radicals in brain aging. These results are in accordance with other studies (Mo et al. 1995; Tian et al. 1998; Kurokawa et al. 2001).
Aging is a complicated multifactorial process in which a progressive decline in the physiologic function of organisms occurs. Oxidative stress describes cell damage caused by an overabundance of oxidants. In balanced status, ROS are produced as a by-product of metabolic processes and the level of ROS can be controlled with antioxidants, such as small molecular weight dietary supplements, including vitamins C and E; small molecular weight peptides and cofactor, including glutathione and pyruvate; and enzymes including SOD, catalase, and glutathione peroxidase. Antiox-idant enzymes are capable of degrading ROS into inert compounds through a series of chemical reactions. But in a state of imbalance, the levels of oxidants outweigh the levels of antioxidants. The relationship between IR and aging is not clear yet, but there are some mechanistic theories that IR
can accelerate aging through free-radical damage and oxida-tive stress (Richardson 2009).
Many scientific communities have become increasingly interested in the brain aging and in pharmaceutical approaches to decelerate the aging process or aging-related change. Accordingly, many efforts are currently directed towards the identification of molecular targets or mechanisms for aging-related neuro-degenerative disease.
Currently irradiation was suggested as a good experimental tool to specifically suppress the adult neurogenesis (Wojtow-icz 2006). In addition, it was found that radiation exposure did not make any significant changes of the oxidative status in brain at 2 month after radiation exposure. These evidences suggest that radiation-induced oxidative status changing may not sustain by 2 month in the brain. The ionizing radia-tion may warrant useful experimental model to support the drug development that specifically decelerates brain aging.
In summary, there were oxidative changes of the brain in which the lipid peroxidation was increased and the anti-oxida-tive enzyme activity was decreased during aging but no late effect of IR on these parameters was observed.
ACKNOWLEDGMENT
This work was supported by grant no. 2010-0005282 from Nuclear R&D Program of Ministry of Education, Science and Technology, Republic of Korea.
REFERENCES
Ando S, Kon K and Totani Y. 1990. Increased levels of lipid peroxides in aged rat brain as revealed by direct assay of peroxide values. Neurosci. Lett. 113:199-204.
Athar M. 2002. Oxidative stress and experimental carcinogene-sis. Ind. J. Expt. Biol. 40:656-667.
Benzi G and Moretti A. 1995. Age- and peroxidative stress-relat-ed modifications of the cerebral enzymatic activities linkstress-relat-ed to mitochondria and the glutathione system. Free Radic. Biol. Med. 19:77-101.
Floyd RA. 1999. Antioxidants, oxidative stress, and denegera-tive neurological disorders. Pro. Soc. Exp. Biol. Med. 222: 236-245.
Hiramatsu M, Korno M, Edamatsu R, Mitsuta K and Mori A. 1992. Increased superoxide dismutase activity in aged human cerebrospinal fluid and rat brain determined by
elec-tron spin resonance spectrometry using the spin trap method. J. Neurochem. 58:1160-1164.
Kang SK, Rabbani ZN, Folz RJ, Golson ML, Huang H, Yu D, Samulski TS, Dewhirst MW, Anscher MS and Vujaskovic Z. 2003. Overexpression of extracellular superoxide dismu-tase protects mice from radiation-induced lung injury. Int. J. Radiat. Oncol. Biol. Phys. 57:1056-1066.
Kurokawa T, Asada S, Nishitani S and Hazeki O. 2001. Age-related changes in manganase superoxide dismutase activity in the cerebral cortex of senescence-accelerated prone and resistant mouse. Neurosci. Lett. 298:135-138.
Leutner S, Eckert A and Muller WE. 2001. ROS generation, lipid peroxidation and antioxidant enzyme activities in the aging brain. J. Neural. Transm. 108:955-967.
Limoli CL, Rola R, Giedzinski E, Mantha S, Huang TT and Fike JR. 2004. Cell density-dependent regulation of neural precursor cells function. Pro. Natl. Acad. Sci. USA. 101: 16052-16057.
Mewissen DJ, Comar CL, Trum BF and Rust JH. 1957. A for-mula for chronic radiation dosage versus shortening of life span: application to a large mammal. Radiat. Res. 6:450-459.
Mo JQ, Hom DG and Andersen JK. 1995. Decreases in protec-tive enzymes correlates with increased oxidaprotec-tive damage in the aging mouse brain. Mech. Ageing Dev. 81:73-78. Moulder JE. 2003. Pharmacological intervention to prevent or
ameliorate chronic radiation injuries. Semin. Radiat. Oncol.
13:73-84.
Navarro A and Boveris A. 2004. Rat brain and liver mitochon-dria develop oxidative stress ad lose enzymatic activities on aging. Am. J. Physiol. Regul. Integr. Comp. Physiol. 287: R1244-R1249.
Ohkawa H, Ohishi N and Yagi K. 1979. Assay for lipid perox-ides in animal tissues by thiobarbituric acid reaction. Anal.
Biochem. 95:351-358.
Peskin AV and Winteerbourn CC. 2000. A microtiter plate assay for superoxide dismutase using a water soluble tetrazolium salt (WST-1). Clinica. Chim. Acta 293:157-166.
Richardson RB. 2009. Ionizing radiation and aging: rejuvenating an old idea. Aging 293:157-166.
Riley PA. 1994. Free radicals in biology: oxidative stress and the effects of ionizing radiation. Int. J. Radiat. Biol. 65:27-33.
Robbins MEC and Zhao W. 2004. Chronic oxidative stress and radiation-induced late normal tissue injury: a review. Int. J. Radiat. Biol. 80:251-259.
Smith CD, Carney JM, Starke-Reed PE, Oliver CN, Stadtman ER, Floyd RA and Markesbery WR. 1991. Excess brain protein oxidation and enzyme dysfunction in stress in nor-mal aging and in Alzheimer disease. Proc. Natl. Acad. Sci. USA. 88:10540-10543.
Stadtman ER. 2006. Protein oxidation and aging. Free Radic. Res. 40:1250-1258.
Tian L, Cai Z and Wei H. 1998. Alterations of antioxidant enzymes and oxidative damage to macromolecules in dif-ferent organs of rats during aging. Free Radic. Biol. Med.
24:1477-1484.
Upton AC, Kimball AW, Furth J, Christenberry KW and Bene-dict WH. 1960. Some delayed effects of atom-bomb radia-tions in mice. Cancer Res. 20:1-60.
Wojtowicz JM. 2006. Irradiations as an experimental tool in studies of adult neurogenesis. Hippocampus. 16:261-266. Zhao W, Diz DI and Robbins ME. 2007. Oxidative damage
pathways in relation to normal tissue injury. Br. J. Radiol.
80:S23-S31.
Manuscript Received: August 16, 2010 Revision Accepted: August 27, 2010