저작자표시-비영리-변경금지 2.0 대한민국 이용자는 아래의 조건을 따르는 경우에 한하여 자유롭게 l 이 저작물을 복제, 배포, 전송, 전시, 공연 및 방송할 수 있습니다. 다음과 같은 조건을 따라야 합니다: l 귀하는, 이 저작물의 재이용이나 배포의 경우, 이 저작물에 적용된 이용허락조건 을 명확하게 나타내어야 합니다. l 저작권자로부터 별도의 허가를 받으면 이러한 조건들은 적용되지 않습니다. 저작권법에 따른 이용자의 권리는 위의 내용에 의하여 영향을 받지 않습니다. 이것은 이용허락규약(Legal Code)을 이해하기 쉽게 요약한 것입니다. Disclaimer 저작자표시. 귀하는 원저작자를 표시하여야 합니다. 비영리. 귀하는 이 저작물을 영리 목적으로 이용할 수 없습니다. 변경금지. 귀하는 이 저작물을 개작, 변형 또는 가공할 수 없습니다.
석사학위논문
제주도산 긴가락박쥐
(Miniopterus
schreibersii)의 외부형태학적 특성
제주대학교 교육대학원
생물교육전공
안 연 하
2016년 8월
제주도산 긴가락박쥐(Miniopterus
schreibersii)의 외부형태학적 특성
지도교수 오 홍 식
안 연 하
이 논문을 교육학 석사학위 논문으로 제출함
2016년 6월
안연하의 교육학 석사학위 논문을 인준함
심사위원장
㊞
위
원
㊞
위
원
㊞
제주대학교 교육대학원
External Morphological Characteristics of
Miniopterus schreibersii in Jeju Island, Korea
Yeon-ha An
(Supervised by Professor Hong-Shik Oh)
A thesis submitted in partial fulfillment of the requirement for the degree of Master of Education
2016. 6.
This thesis has been examined and approved.
____________________________________
Thesis director, Keun-Jae Ahn, Prof. Faculty of Science Education____________________________________
____________________________________
_______________
Department of Biology Education
GRADUATE SCHOOL OF EDUCATION
<국문초록>
제주도산 긴가락박쥐(Miniopterus
schreibersii)의 외부형태학적 특성
안 연 하 제주대학교 교육대학원 생물교육전공 지도교수 오 홍 식 이 연구는 제주도에 서식하고 있는 긴가락박쥐(Miniopterus schreibersii)의 외부 형태학적 특징을 밝혀 서식지 및 생물다양성 보호에 필요한 정보를 제공하기 위 하여 이루어졌다. 현장조사는 2014년 3월부터 2016년 4월까지 제주도 내 용암동 굴(13개 지역), 인공 진지동굴(18개 지역), 해식동굴(2개 지역), 궤(2개 지역) 등 35개 지역에서 이루어졌다. 조사결과, 문헌기록을 포함하여 적어도 11개 지역에 서 긴가락박쥐의 서식이 확인되었다. 긴가락박쥐는 대부분 100개체 이상 무리로 관찰되었고, 간혹 1개체씩 관찰되기도 하였다. 서식지역은 용암동굴과 인공 진지 동굴, 산림 지역 등이었으나, 대규모 집단이 제주도 동부와 서부지역에서 발견되 었다. 제주도 집단의 형태형질 측정치 중 머리-몸통의 길이를 제외한 대부분의 형질들이 제주도 집단과 한반도 집단 사이에서 유사한 양상을 보였다. 반면, 우 리나라의 두 집단(제주도, 한반도 집단)과 일본 집단 사이에서는 지리적 형태변 이가 뚜렷하게 나타났다. 제주도 집단 내에서 암컷과 수컷은 체중(BW), 귀/이주 길이의 비율(ETR), 상완의 길이(Hum), 종아리의 길이(Tib)에서 유의적인 차이 (P<0.05)를 보였다. 이 중 BW과 ETR은 암컷 우세적인 양상을 보였으나, BW는 임신기간 동안 증가된 체중에 기인한 것으로 추정되었다. 반면, Hum과 Tib은 수 컷 우세적인 양상을 보였다. 연구를 통해 긴가락박쥐의 서식은 제주도 내 각처에 서 용암동굴, 인공 진지동굴, 산림 등 다양한 서식지를 확인하였으며, 형태적으로 는 제주도 집단의 머리-몸통의 길이가 한반도 집단보다 크지만, 일본 집단보다는작은 특성을 보였으며, 제주도 집단 내에서 암-수의 형태적 차이를 나타내었다. 이러한 결과들은 향후 제주도 천연동굴이나 인공 동굴의 보전과 생태관광 프로 그램의 개발 등에서 반드시 제고되어야 할 중요한 정보들을 내포하고 있다. 또한 긴가락박쥐 서식지의 분포, 타 지역 집단들과의 지리적 형태 변이, 암-수 간의 성 적 크기 이형을 나타내는 형질들은 향후 한반도와 일본, 중국 등 동아시아의 여 러 지역에서 충분한 자료의 수집을 통한 종합적인 연구가 필요함을 시사하고 있 다. 본 연구결과들은 현재까지 생태적 특성이 거의 알려진 바 없는 우리나라 긴 가락박쥐를 집단 수준에서 다룬 최초의 연구결과라는 점에서, 긴가락박쥐의 종적 집단 특성을 이해하는 데 도움을 줄 수 있는 유용한 자료가 될 것이다.
목 차
국문초록 ··· ⅰ 목 차 ··· ⅲ List of Tables ··· ⅳ List of Figures ··· ⅴ Ⅰ. 서 론 ··· 1 Ⅱ. 재료 및 방법 ··· 4 1. 조사지역 ··· 4 2. 조사방법 ··· 6 3. 외부형태 형질 측정 ··· 6 4. 외부형태의 집단 간 비교 ··· 7 5. 암-수 사이의 외부형태 형질 비교 ··· 8 Ⅲ. 결과 및 고찰 ··· 9 1. 긴가락박쥐의 서식지 확인 및 외부형태 관찰 ··· 9 2. 제주도 긴가락박쥐 집단의 외부형태 측정 ··· 12 3. 제주도 집단과 타 지역 집단 비교 ··· 14 4. 암-수 사이의 외부형태 형질 비교 ··· 17 Ⅳ. 참고문헌 ··· 22 Abstract ··· 30List of Tables
Table 1. Location and type of survey sites in this study ··· 5
Table 2. Collection site and number of the M. schreibersii specimens used in this study ··· 12
Table 3. Mean and SD of external morphological characters of M. schreibersii measured in this study ··· 13
Table 4. Comparison of the external morphological characters of the M. schreibersii populations of Japan, Korean Peninsula and Jeju Island ··· 14
Table 5. Comparison of the external morphological characters between both sexes of
List of Figures
Figure 1. Distribution map of M. schreibersii in the world ··· 2
Figure 2. Survey and collection sites of M. schreibersii in this study ··· 4
Figure 3. The positions of each measurement of external morphological characters of M. schreibersii ··· 7
Figure 4. Finding and recording sites of hibernation of M. schreibersii ··· 9
Figure 5. A group of M. schreibersii found in Albeolari-gul in March 2016 ··· 10
Figure 6. External morphology of the M. schreibersii ··· 11
Figure 7. Comparison of SDI levels of external morphological characters among the M. schreibersii populations of Japan, Korean Peninsula and Jeju Island ··· 16
Figure 8. SSDI levels of external morphological characters between both sexes of M. schreibersii ··· 19
Figure 9. Averages (A) and monthly changes (B) of body weights between males and females of M. schreibersii. ··· 20
Ⅰ
. 서론
전 세계 포유류는 5,000여 종으로 그 중 박쥐류(Chiroptera)는 1/4 이상을 차지 하는 큰 분류군이다(Willson and Reeder, 2005). 매우 추운 지역(남극, 북극)과 몇 몇 대양의 섬들을 제외한 전 세계에 분포하며, 대부분 곤충을 섭식하나 무척추동 물, 양서류, 어류, 과일 꽃 등 다양한 먹이원을 섭취한다. 소익수아목(Microchiroptera)은 일반적으로 이주가 있으며, 대퇴부 사이에 막 (interfemoral membrane)이 잘 발달되어 있고, 꼬리는 비교적 긴 특성을 보이지만, 형태적으로 아주 다양한 변이를 나타내기도 한다(Hutson et al, 2001). 우리나라에 서식하는 박쥐류는 대부분 소익수아목에 속하며 자연동굴이나 인공동굴, 삼림, 건물의 천장 등에서 서식한다(Kunz, 1982; Altringham et al., 1998; Kunz and Lumsden, 2003; Patterson et al., 2003; 윤 등, 2004). 국내에 분포하는 박쥐류에 대 한 분류체계는 학자들간 서로 다른 견해를 보이고 있지만, 현재 3과 11속 23종으 로 분류되고 있다(한 등, 2011).
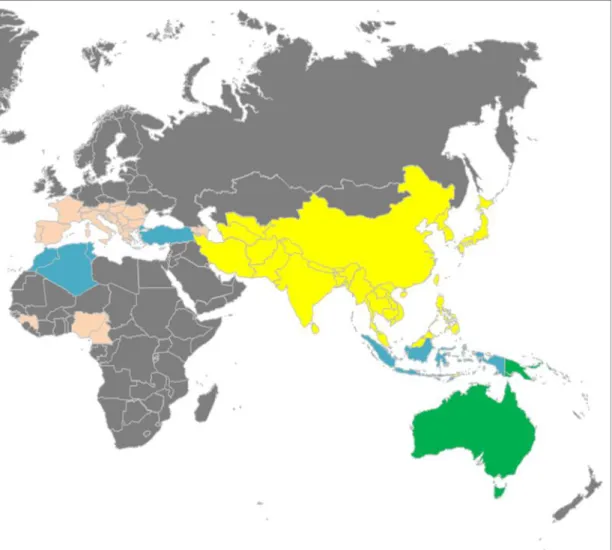
긴가락박쥐(Miniopterus schreibersii)는 소익수아목, 애기박쥐과(Vespertilionidae), 긴가락박쥐아과(Miniopterinae), 긴가락박쥐속(Miniopterus)에 속하며, 유럽 남부, 아 프리카 북부와 사하라 사막 이남, 동아시아, 남아시아 등 구대륙과 호주 등지에 서식하는 것으로 알려져 있다(Maeda, 1982; 윤 등, 2004; Tian et al., 2004; Abe et
al., 2005; 윤, 2010; Puechmaille et al., 2014). 우리나라에서는 제주도를 포함하여
강원도 강릉, 철원, 충북, 충남, 경남 통영 등 전국에 분포한다(윤과 손, 1989; 윤, 2010). 한반도로부터 지리적으로 격리된 이후 오랜 기간을 걸쳐 오는 동안 제주도 육 상 척추동물은 종의 분포나 형태는 물론 유전적으로도 독특한 생물학적 특성을 보여주고 있다. 예를 들어, 비바리뱀(Sibynophis chinensis)은 제주도가 생물지리적 으로 북방한계선이고, 무당개구리(Bombina orientalis)는 남방한계선이다(김과 오, 2005; Kim and Oh, 2006; 장 등, 2010). 포유류에서도 노루(Capreolus pygargus)는 한반도 집단에 비해 체형이 작고, 유전적으로도 서로 다르며(Koh et al., 2000; 윤
등, 2004), 제주등줄쥐(Apodemus chejuensis), 작은땃쥐(Crocidura shantungensis) 등 도 한반도 집단과 형태 및 유전적인 차이를 보이고 있다(Oh and Mori, 1998; Motokawa et al., 2003; Oh et al., 2013; 김 등, 2015). 박쥐류 연구에서, Mori (1933)는 김녕사굴에서 채집된 관박쥐가 일본의 R. f. nippon과 유사하지만 두동장 (head-body length)과 비막의 색 등 외부 형태적 차이가 있어 제주관박쥐(R. f.
quelpartis)로 명명하였다(Mori, 1933). 또한 김(2015)은 제주도 Myotis 속 4종에 관
한 연구를 통해 제주도 집단이 한반도 집단에 비해 체형이 큰 경향성이 있음을 강조하면서 분자유전학적으로도 차이가 있음을 보고한 바 있다.
Fig. 1. Distribution map of M. schreibersii in the world.
제주도에서 긴가락박쥐의 분포에 대해서는 남궁(1981)이 와흘굴과 만장굴 조사 에서 처음으로 보고하였고, 손(1981)은 대흘리 자연동굴에서 긴가락박쥐의 서식 을 기록하였다. 이후 양과 홍(1988), 채와 박(1988)은 제주도 용암동굴에서 긴가락 박쥐의 서식과 채집을 보고하였으며, 2000년대 이후에는 지속적으로 관찰기록이 이루어지고 있다. 오(2002)는 제주도에 긴가락박쥐뿐만 아니라 작은긴가락박쥐(M. fuscus)도 서식하는 것으로 보고하였으며, 오 등(2007)은 한라산국립공원 조사를 통해 관음사 지역에서 관박쥐(R. ferrumequinum), 큰발윗수염박쥐(M.
macrodactylus), 집박쥐(Pipistrellus abramus) 등과 함께 긴가락박쥐의 서식을 보고
하였다. 또한 Jo et al. (2012)은 제주도 포유류 현황에서 관박쥐, 흰배윗수염박쥐, 붉은박쥐 등 2과 5속 12종을 기술하면서 작은긴가락박쥐, 긴가락박쥐를 포함하였 다. 오(2013)는 제주도 자연동굴과 인공 진지동굴 5개 지역에서 약 7,500개체를 관찰하였고, 최근 박 등(2015)은 제주도 7개 지역에서 2,530개체를 기록하였다. 현재까지 제주도 긴가락박쥐에 대한 연구는 자연동굴과 진지동굴에서의 서식 을 확인하는 수준에 머물러 있어, 제주도 집단의 특성을 설명할 수 있는 자료는 마련된 바 없다. 이에 본 연구는 다른 지역 집단과의 비교를 통해 제주도에 서식 하고 있는 긴가락박쥐의 외부형태학적 특징을 밝혀 서식지 및 생물다양성 보호 를 마련하는 데 필요한 정보를 제공하기 위하여 이루어졌다.
Ⅱ
. 재료 및 방법
1. 조사 지역
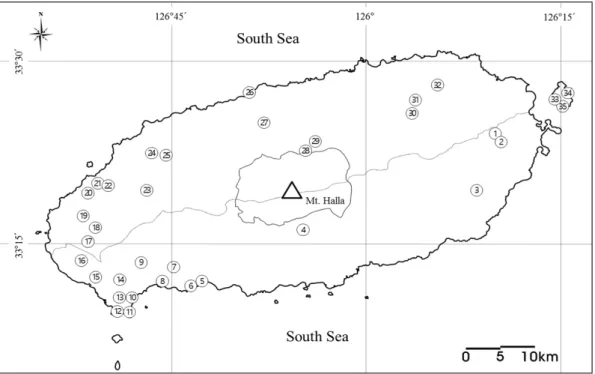
제주도 내에서 긴가락박쥐에 대한 조사는 2014년 3월부터 2016년 4월까지 제 주시 19개 지역, 서귀포시 16개 지역을 대상으로 수행되었다(Fig. 2, Table 1). 세 부적으로 보면, 용암동굴(lava tube, LT) 13개 지역, 진지동굴(cave encampment, CE) 18개 지역, 해식동굴(sea cave, SC) 2개 지역, 궤(natural cavelet, NC) 2개 지역 이다.
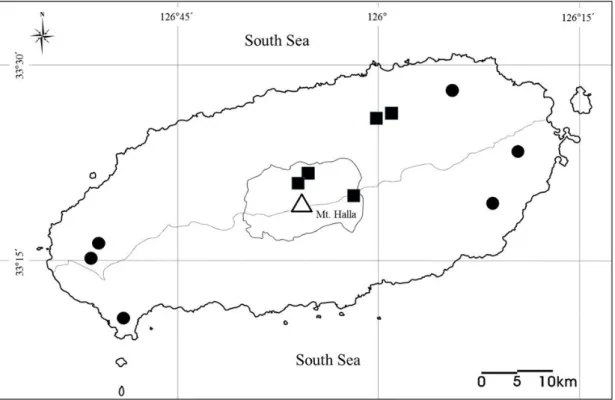
Fig. 2. Survey and collection sites of M. schreibersii in this study. Circles indicate survey site of M. schreibersii. The number, location and type of each site are given in Table 1.
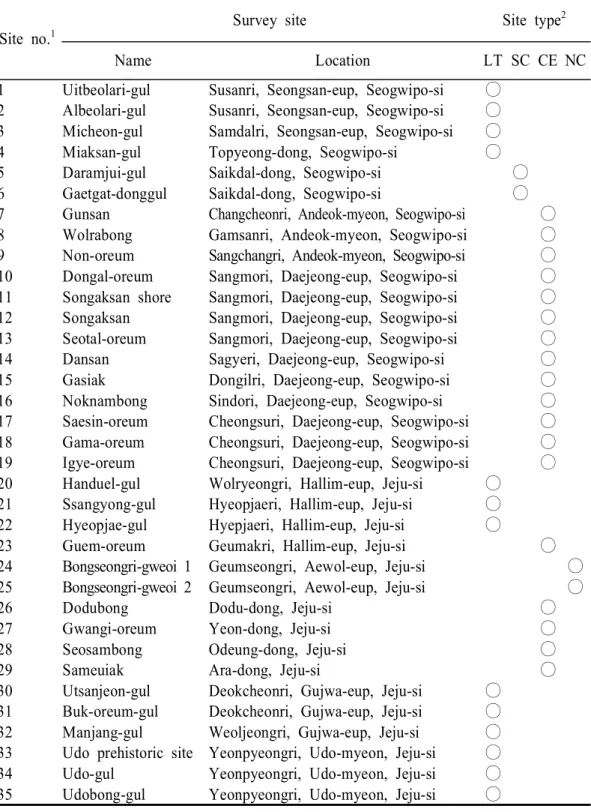
Table 1. Location and type of survey sites in this study
Site no.1 Survey site Site type 2
Name Location LT SC CE NC
1 Uitbeolari-gul Susanri, Seongsan-eup, Seogwipo-si ○ 2 Albeolari-gul Susanri, Seongsan-eup, Seogwipo-si ○ 3 Micheon-gul Samdalri, Seongsan-eup, Seogwipo-si ○ 4 Miaksan-gul Topyeong-dong, Seogwipo-si ○ 5 Daramjui-gul Saikdal-dong, Seogwipo-si ○ 6 Gaetgat-donggul Saikdal-dong, Seogwipo-si ○ 7 Gunsan Changcheonri, Andeok-myeon, Seogwipo-si ○ 8 Wolrabong Gamsanri, Andeok-myeon, Seogwipo-si ○ 9 Non-oreum Sangchangri, Andeok-myeon, Seogwipo-si ○ 10 Dongal-oreum Sangmori, Daejeong-eup, Seogwipo-si ○ 11 Songaksan shore Sangmori, Daejeong-eup, Seogwipo-si ○ 12 Songaksan Sangmori, Daejeong-eup, Seogwipo-si ○ 13 Seotal-oreum Sangmori, Daejeong-eup, Seogwipo-si ○ 14 Dansan Sagyeri, Daejeong-eup, Seogwipo-si ○ 15 Gasiak Dongilri, Daejeong-eup, Seogwipo-si ○ 16 Noknambong Sindori, Daejeong-eup, Seogwipo-si ○ 17 Saesin-oreum Cheongsuri, Daejeong-eup, Seogwipo-si ○ 18 Gama-oreum Cheongsuri, Daejeong-eup, Seogwipo-si ○ 19 Igye-oreum Cheongsuri, Daejeong-eup, Seogwipo-si ○ 20 Handuel-gul Wolryeongri, Hallim-eup, Jeju-si ○
21 Ssangyong-gul Hyeopjaeri, Hallim-eup, Jeju-si ○ 22 Hyeopjae-gul Hyepjaeri, Hallim-eup, Jeju-si ○
23 Guem-oreum Geumakri, Hallim-eup, Jeju-si ○ 24 Bongseongri-gweoi 1 Geumseongri, Aewol-eup, Jeju-si ○ 25 Bongseongri-gweoi 2 Geumseongri, Aewol-eup, Jeju-si ○ 26 Dodubong Dodu-dong, Jeju-si ○ 27 Gwangi-oreum Yeon-dong, Jeju-si ○ 28 Seosambong Odeung-dong, Jeju-si ○ 29 Sameuiak Ara-dong, Jeju-si ○ 30 Utsanjeon-gul Deokcheonri, Gujwa-eup, Jeju-si ○
31 Buk-oreum-gul Deokcheonri, Gujwa-eup, Jeju-si ○ 32 Manjang-gul Weoljeongri, Gujwa-eup, Jeju-si ○ 33 Udo prehistoric site Yeonpyeongri, Udo-myeon, Jeju-si ○ 34 Udo-gul Yeonpyeongri, Udo-myeon, Jeju-si ○ 35 Udobong-gul Yeonpyeongri, Udo-myeon, Jeju-si ○
1, the number of each site are marked in Fig. 2.
2. 조사방법
조사는 현장을 방문하여 긴가락박쥐의 서식여부와 개체수를 파악하였고, hand-net (Kunz and Kurta, 1988)을 이용하여 포획한 후 관찰하였다. 포획된 박쥐 는 관찰일자, 포획지점, 성별 등을 기록하였다. 포획하거나 관찰한 개체 중 일부 는 실험실로 옮겨 외부형태형질을 측정하였다. 측정을 마친 개체들은 ethanol (80-90%)을 이용하여 액침표본을 제작하였다.
3. 외부형태 형질 측정
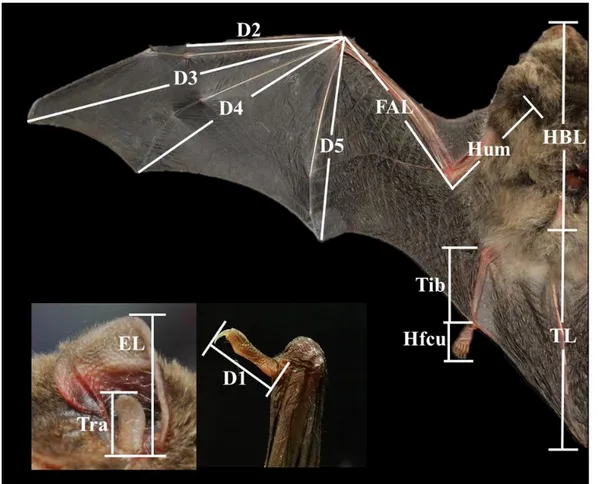
외부 형질은 체중(body weight, BW), 머리-몸통의 길이(head-body length, HBL), 꼬리의 길이(tail length, TL), 몸 전체의 길이(whole body length, WBL), 앞팔의 길 이(forearm length, FAL), 상완의 길이(humerus length, Hum), 귀의 길이(ear length, EL), 이주의 길이(tragus length, Tra), 종아리의 길이(tibia length, Tib), 첫 번째 손 가락의 길이(1st digit length, D1), 두 번째 손가락의 길이(2nd digit length, D2), 세 번째 손가락의 길이(3rd digit length, D3), 네 번째 손가락의 길이(4th digit length, D4), 다섯 번째 손가락의 길이(5th digit length, D5)를 측정하였다(Fig. 3). 단, 몸 전체의 길이(whole body length, WBL)는 HBL과 TL의 합산값을 이용하였다. 외부 형질들 중 길이의 비례적인 관계를 알아보기 위해 BTR (머리-몸통의 길 이에 대한 꼬리 길이의 비율), ETR (귀의 길이에 대한 이주길이의 비율), D35R (다섯 번째 손가락의 길이에 대한 세 번째 손가락 길이의 비율) 등을 산출하였 다. 외부 형태형질과 관련한 용어는 Yoshiyuki (1989)와 윤(2010)을 참고하였다. 체 중은 전자저울(MW11300, Cas, Korea)을 이용하여 0.1g 수준까지 측정하였고, 길 이는 디지털 캘리퍼스(500-181-20, Mitutoyo, Japan)를 이용하여 0.01mm 수준까지 측정하였다.
Fig. 3. The positions of each measurement of external morphological characters of M.
schreibersii. HBL, head-body length; FAL, forearm length; Hum, humerus length; TL,
tail length; EL, ear length; Tra, tragus length; Tib, tibia length; D1, 1st digit length; D2, 2nd digit length; D3, 3rd digit length; D4, 4th digit length; D5, 5th digit length.
4. 외부형태의 집단 간 비교
제주도에서 채집한 긴가락박쥐 집단과 기존에 보고된 타 지역 집단의 외부 형 태적 차이를 비교하기 위해 산출된 평균값의 비율을 백분율로 환산한 크기차이 지수(size difference index, SDI)를 이용하여 비교하였다.
여기서, k는 개별 형질, 는 a집단의 k형질의 평균값, 는 제주도 집단의 k 형질의 평균값을 의미한다. 즉, 제주도 집단의 형질 평균값은 100으로 환산되며, 제주도 집단의 평균값보다 큰 경우는 100 이상의 값을, 작은 경우는 100이하의 값을 나타낸다. 5. 암-수 사이의 외부형태 형질 비교 제주도 긴가락박쥐 집단의 암컷과 수컷 사이의 성적 크기 이형(sexual size dimorphism, SSD)을 확인하기 위하여 암-수 성별에서 측정된 평균값을 비교하였 다. 평균값 차이에 대한 유의성은 Student’s t-test를 사용하였고(SPSS Statistics, ver. 19.0), 유의성 검사는 95% 신뢰수준에서 이루어졌다. 그리고 성별에 따른 크 기차이의 정도를 정량화하기 위하여 Levenson (1990)의 방법에 따라 각 형질에 대한 성적 크기이형지수(sexual size dimorphism Index, SSDI) (Zidarova, 2015)를 산 출하여 비교하였다. × 여기서 은 암컷 형질의 평균, 은 수컷 형질의 평균을 나타내며, 암컷의 평균이 수컷보다 크면 양의 값(), 수컷보다 작으면 음의 값()으로 나타내었다.
Ⅲ
. 결과 및 고찰
1. 긴가락박쥐의 서식지 확인 및 외부형태 관찰
제주도 내 35개 지역에 대한 현장조사를 통해 확인된 긴가락박쥐의 서식 지역 은 총 5개 지역이었으며, 확인된 장소는 용암동굴 2개 지역(벌라릿굴, 미천굴)과 인공 진지동굴 3개 지역(가마오름, 새신오름, 섯알오름)이었다(Fig. 4).
Fig. 4. Finding and recording sites of hibernation of M. schreibersii. Circles and squares indicate the finding sites in this study and those previously recorded, respectively.
기존 연구결과에 의하면, 제주도에서 긴가락박쥐가 관찰 또는 채집된 지역은 9 개 지역(벌라릿굴, 만장굴, 구린굴, 와흘굴, 대흘리 자연동굴, 섯알오름 진지동굴, 가마오름 진지동굴, 한라산 산림지역인 물장올교와 관음사 등산로)이었다. 따라서
문헌기록과 본 연구결과를 종합했을 때, 제주도에서 긴가락박쥐는 적어도 11개 장소에서 관찰되거나, 채집되었다. 반면, 조천읍 대흘리 자연동굴은 정확한 소재 를 파악할 수 없어 재확인하지 못하였고, 와흘굴과 만장굴 미공개구간은 출입이 통제되어 재확인할 수 없었다. 또한 구린굴과 물장올교, 관음사 등산로 등에 대 해서는 추가적인 확인이 필요하다. 본 연구 동안 미천굴 공개구간에서 긴가락박 쥐의 서식이 처음으로 확인되었으나, 미천굴 미공개구간에 대한 조사가 이루어지 지 않아, 향후 박쥐류 서식과 관련한 종의 분포, 집단의 크기나 이용시기 등에 대한 면밀한 조사가 필요하다고 하겠다. 서식지에서 발견되는 형태는 1개체씩 발견되는 경우도 있었고, 100개체 이상, 많게는 대략 1,500개체가 무리를 형성하는 경우도 관찰되었다. 한 동굴 안에서 발견되는 모든 개체들이 하나의 무리를 형성하는 경우도 있었으나, 대부분 동굴 내에서 2-3개의 무리로 나누어진 형태로 관찰되기도 하였다.
제주도 내 5개 지역에 서식하는 긴가락박쥐의 외부 형태를 관찰한 결과(Fig. 6), 몸의 털은 윤택이 있었으며, 암갈색, 담갈색 등으로 나타났으며, 아랫면 털은 윗면보다 엷었다. 양쪽 귀의 좌우 간격은 넓었으며, 귀와 이주의 끝이 둥글며, 비막은 종아리의 아랫부분에 부착되어 있었다. 꼬리는 꼬리 비막 밖으로 돌출해 있지 않았으며, 꼬리 비막 위쪽에 짧은 털이 있었다. 전반적으로 긴가락박쥐의 외부 형태에 대한 기존 기술 내용(Wallin, 1969; Yoshiyuki, 1989; 윤 등, 2004; Dietz and von Helversen, 2004; 윤, 2010)과 대체로 유사하였으며, 제주도 집단에 서만 특이적으로 관찰된 외형상의 특징은 없었다.
Fig. 6. External morphology of M. schreibersii. (A) M. schreibersii found in natural roost of cave encampment in March 2016; (B), ear and tragus; (C), hind foot; (D), adult female, captured in June 2015.
2. 제주도 긴가락박쥐 집단의 외부형태 측정
조사과정에서 긴가락박쥐의 집단 서식이 확인된 4개 지역에서 성체인 암컷(43 개체)과 수컷(38개체) 총 81개체에 대한 측정 결과는 Table 2에 제시한 바와 같 다.
Table 2. Collection site and number of the M. schreibersii specimens used in this study
Collection site No. of individuals Site name Site no.1 Site type Female Male Sum
Albeolari-gul 2 Lava tube 17 5 22 Seotal-oreum 13 Cave encampment 2 22 24 Saesin-oreum 17 Cave encampment 23 10 33
Gama-oreum 18 Cave encampment 1 1 2
Sum 43 38 81
1, site numbers are same with those given in Fig. 2 and Table 2.
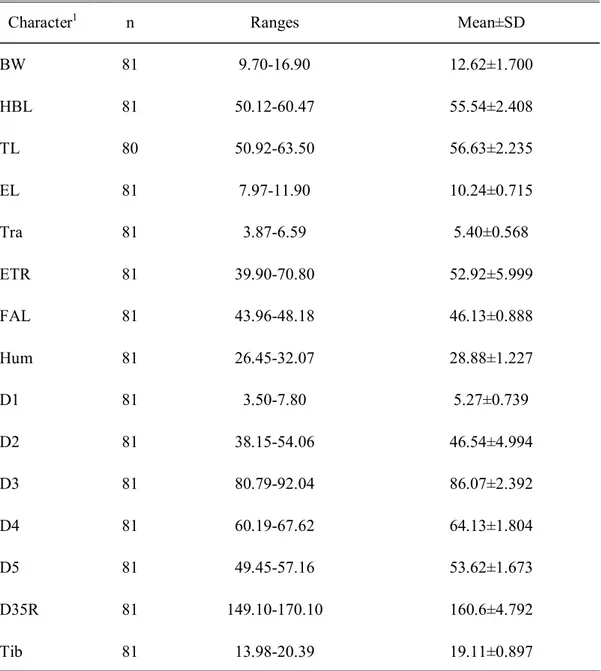
측정 결과, BW는 9.7-16.9g, 평균 12.62±1.700g으로 중형종임을 알 수 있었고, Walker (1975)가 보고한 범주 내에 있었다. HBL는 50.12-60.47mm, 평균 55.54±2.408mm, TL은 50.92-63.50mm, 평균 56.63±2.235mm를 나타내어, 윤 등 (2004)이 제시한 HBL (47-57mm)와 TL (34-51mm)보다 다소 큰 범위를 보였다. EL은 7.97-11.90mm, 10.24±0.715mm를 보였고, Tra는 3.87-6.59mm의 범위에서 출 현하고 평균 5.40±0.568mm였다. 이주 길이가 귀 길이의 52.92±5.999을 나타내었 다. FAL의 길이는 43.96-48.18mm로 측정되어, Walker (1975)와 윤 등(2004)의 범 위 내에 있었다. D35R (D3와 D5의 비율)은 140-170%, 평균 160.6±4.792%로 좁고 긴 협장형 날개를 갖는 긴가락박쥐의 특성(Macdonald and Barrett, 1993; 윤 등,
을 결정하는 주된 형질인 것으로 제안되었다(Struhsaker, 1961; Findley, 1972; Norberg and Rayner, 1987).
Table 3. Mean and SD of external morphological characters of M. schreibersii measured in this study
Character1 n Ranges Mean±SD
BW 81 9.70-16.90 12.62±1.700 HBL 81 50.12-60.47 55.54±2.408 TL 80 50.92-63.50 56.63±2.235 EL 81 7.97-11.90 10.24±0.715 Tra 81 3.87-6.59 5.40±0.568 ETR 81 39.90-70.80 52.92±5.999 FAL 81 43.96-48.18 46.13±0.888 Hum 81 26.45-32.07 28.88±1.227 D1 81 3.50-7.80 5.27±0.739 D2 81 38.15-54.06 46.54±4.994 D3 81 80.79-92.04 86.07±2.392 D4 81 60.19-67.62 64.13±1.804 D5 81 49.45-57.16 53.62±1.673 D35R 81 149.10-170.10 160.6±4.792 Tib 81 13.98-20.39 19.11±0.897
3. 제주도 집단과 타 지역 집단 비교
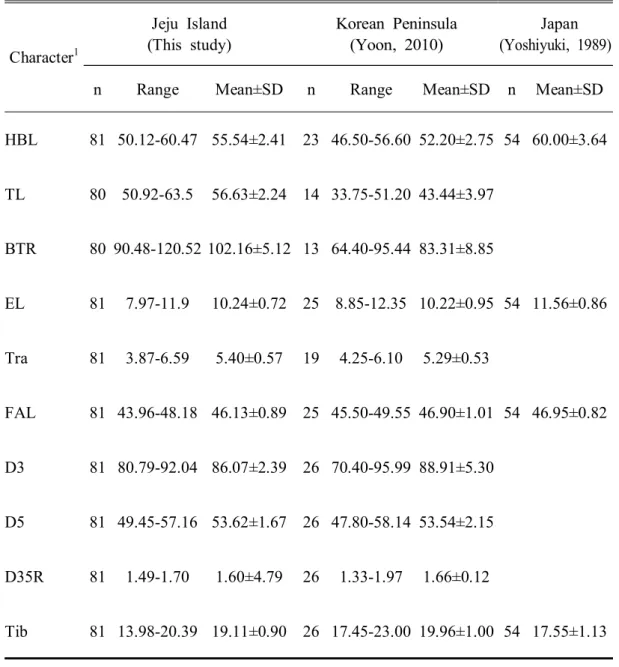
긴가락박쥐 제주도 집단과 한반도 집단(윤, 2010), 일본 집단(Yoshiyuki, 1989) 을 비교한 결과는 Table 4에 제시한 바와 같다.
Table 4. Comparison of the external morphological characters of the M. schreibersii populations of Japan, Korean Peninsula and Jeju Island
Character1 Jeju Island (This study) Korean Peninsula (Yoon, 2010) Japan (Yoshiyuki, 1989)
n Range Mean±SD n Range Mean±SD n Mean±SD
HBL 81 50.12-60.47 55.54±2.41 23 46.50-56.60 52.20±2.75 54 60.00±3.64 TL 80 50.92-63.5 56.63±2.24 14 33.75-51.20 43.44±3.97 BTR 80 90.48-120.52 102.16±5.12 13 64.40-95.44 83.31±8.85 EL 81 7.97-11.9 10.24±0.72 25 8.85-12.35 10.22±0.95 54 11.56±0.86 Tra 81 3.87-6.59 5.40±0.57 19 4.25-6.10 5.29±0.53 FAL 81 43.96-48.18 46.13±0.89 25 45.50-49.55 46.90±1.01 54 46.95±0.82 D3 81 80.79-92.04 86.07±2.39 26 70.40-95.99 88.91±5.30 D5 81 49.45-57.16 53.62±1.67 26 47.80-58.14 53.54±2.15 D35R 81 1.49-1.70 1.60±4.79 26 1.33-1.97 1.66±0.12 Tib 81 13.98-20.39 19.11±0.90 26 17.45-23.00 19.96±1.00 54 17.55±1.13
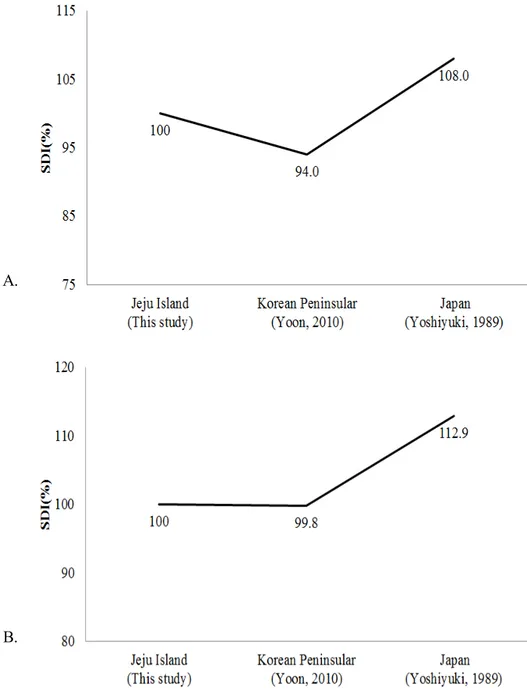
제주도 집단과 한반도 집단(윤, 2010)을 비교하였을 때, HBL은 제주도 집단 (55.54±2.41mm)이 한반도 집단(52.2±2.75mm)보다 3.34mm 정도 더 길었고, TL은 제주도 집단(56.63±2.24)이 한반도 집단(43.44±3.97mm)보다 13.19mm 정도 더 길었 고, BTR은 제주도 집단(102.16±5.12%)이 한반도 집단(83.31±8.85%)보다 18.85% 정도 더 크게 나타났다. EL, Tra, D5은 제주도 집단이 한반도 집단보다 0.02mm, 0.11mm, 0.08mm 정도 더 길었다. 반면 D3은 한반도 집단(88.91±5.30mm)이 제주 도 집단(86.07±2.39mm)보다 2.84mm정도 더 길었고, D35R은 한반도 집단 (1.66±0.12)이 제주도 집단(1.60±4.79)보다 0.06 더 크게 나타났으며, Hfcu은 한반 도 집단(10.56±1.05mm)이 제주도 집단(9.12±1.12mm)보다 1.44mm 정도 더 길었고, Tib는 한반도 집단(19.96±1.00mm)이 제주도 집단(19.11±0.90mm)보다 0.85mm 정도 더 길었다. 집단 간 크기 차이를 비교한 SDI의 수준을 비교해 볼 때(Fig. 7), 제주도 긴가 락박쥐 집단의 HB는 한반도 집단에 비해 약 6% 정도 크게 나타났으며(Fig. 7A), EL은 제주도 집단이 한반도 집단보다 0.2% 정도 컸다(Fig. 7B). FAL은 제주도 집 단보다 한반도 집단이 1.7% 정도 더 큰 양상을 보였고(Fig. 7C), 반면, Tib의 경 우 제주도 집단보다 한반도 집단이 4% 정도 더 길었다(Fig. 7D). 일본 집단과 비 교했을 때, HB는 한반도와 제주도 집단에 비해 일본 집단이 8% 이상 더 컸다. 일본 집단의 EL은 제주도-한반도 집단보다 12% 이상 더 크게 나타났다(Fig. 7B). 일본 집단의 FAL은 제주도-한반도 집단과 거의 유사한 양상을 보였다(Fig. 7C). 일본 집단의 Tib는 제주도-한반도 집단에 비해 8% 이상 작았다(Fig. 7D). 결과적 으로 전체적인 몸의 크기는 일본 집단이 우리나라 집단보다 더 크고, 우리나라에 서는 제주도 집단이 한반도 집단보다 더 큰 양상을 보였다. 긴가락박쥐는 세계적으로 17개의 아종으로 구분될 정도로, 지리적 형태 변이 (geographical morphological variation)를 나타내는 대표적인 생물종으로 알려져 있 고(Dwyer, 1963; Richardson, 1977; Wallace, 1978; Albayrak and Coskun 2000), 학자 들에 따라서는 지리적 변이와 유전적 차이를 기반으로 종 분류체계를 재정비해 야한다고 제안되기도 하였다(Maeda, 1982; Tian et al., 2004; Bilgin et al., 2006). 특히, 태평양의 섬과 호주 등에 분포하는 집단은 M. australis를 학명으로 채택하 고 있으며(Appleton et al., 2004; Puechmaille et al., 2014), 이외에도 우리나라를 포
함한 동아시아의 M. s. fuliginosus를 완전한 종으로 분류해야한다는 의견도 제기
되고 있다(Maeda, 1982; Tian et al., 2004; Mahmood-ul-Hassan and Salim, 2015).
A.
B.
Fig. 7. Comparison of SDI levels of external morphological characters among the M.
schreibersii populations of Japan, Korean Peninsula and Jeju Island. A, HBL; B, EL;
C, FAL; D, Tib. All abbreviations of each trait are given in the Materials and Methods section.
C.
D.
Fig. 7. Continued.
4. 암-수 사이의 외부형태 형질 비교
수집된 긴가락박쥐의 암컷과 수컷에서 측정된 외부형태를 비교하였다(Table 5). 측정된 자료 중 BW, ETR, Hum, Tib 등은 암-수 사이에서 유의적인 차이를 보였
다(P<0.05). 반면 HBL, TL, Body, BTR, EL, Tra, FAL, D1, D2, D3, D4, D5, D35R, Hfcu는 암-수 사이에서 유의한 차이는 없었다(P>0.05).
Table 5. Comparison of the external morphological characters between both sexes of
M. schreibersii
Characters1 Females Males P-value Significance2
n Mean±SD n Mean±SD BW 43 13.32±1.898 38 11.83±0.973 2.68×10-5 *** HBL 43 55.79±2.272 38 55.27±2.557 0.338 n.s. TL 42 56.67±1.976 38 56.60±2.518 0.889 n.s. Body 42 112.40±3.187 38 111.86±4.235 0.527 n.s. BTR 42 50.42±1.272 38 50.60±1.240 0.523 n.s. EL 43 10.15±0.792 38 10.35±0.608 0.199 n.s. Tra 43 5.510±0.495 38 5.28±0.627 0.077 n.s. ETR 43 54.52±5.614 38 51.11±5.976 0.010 * FAL 43 46.12±0.911 38 46.14±0.873 0.943 n.s. Hum 43 28.63±1.212 38 29.17±1.195 0.048 * D1 43 5.21±0.740 38 5.34±0.742 0.428 n.s. D2 43 46.77±5.064 38 46.29±4.970 0.671 n.s. D3 43 86.42±2.404 38 85.67±2.346 0.162 n.s. D4 43 64.21±1.639 38 64.05±1.994 0.702 n.s. D5 43 53.63±1.361 38 53.61±1.987 0.972 n.s. D35R 43 161.19±4.204 38 159.94±5.361 0.244 n.s. Hfcu 43 9.26±1.131 38 8.97±1.096 0.250 n.s. Tib 43 18.85±1.021 38 19.39±0.627 0.006 **
1, abbreviations of each trait are given in the Materials and Methods section.
2, mean±SD values in the same row with different letters are significantly different at
BW에서 암컷(13.32±1.898g)이 수컷(11.83±0.973g)에 비해 약 1.49g 더 무거웠고 (P=2.68×10-5), 귀의 길이에 대한 이주 길이의 비율인 ETR은 암컷(54.42±5.614%) 이 수컷(51.11±5.976%)에 비해 약 3.31% 정도 더 컸다(P=0.010). 반면, 상완의 길 이(Hum)은 수컷(29.17±1.195mm)이 암컷(28.63±1.212mm)에 비해 0.54mm 정도 더 길었으며(P=0.048), 종아리의 길이(Tib)에서도 수컷(19.39±0.627mm)이 암컷 (18.85±1.021mm)에 비해 0.54mm 정도 더 길었다(P=0.006).
Fig. 8. SSDI levels of external morphological characters between both sexes of M.
schreibersii. Positive and negative values indicate female-dominant and male-dominant
characters, respectively.
암컷과 수컷의 평균값 사이에서 암-수 편향성을 살펴보기 위하여 SSDI를 산출 하여 비교하였다(Fig. 8). 조사된 총 18개의 형질들 중에서 암컷 우세적인(female dominant) 양상을 나타내는 형질은 총 11개의 형질이었으나, 통계적인 유의차를 나타내는 형질은 BW와 ETR이었다. 반면, 수컷 우세적인(male dominant) 형질은 총 5개의 형질(BTR, EL, Hum, D1, Tib)이었는데, 이 중 Hum과 Tib에서만 통계적 으로 유의한 차이를 보였다.
대해 월별변화를 확인한 결과(Fig. 9B), 6월에 급격하게 감소하는 경향이 있었고, 6월부터 암컷 무리 내에서 새끼들이 관찰되어, 긴가락박쥐가 6월경에 출산하면서 암컷들의 체중이 감소하는 것으로 보인다. 긴가락박쥐 속 동물들은 대부분 이전 해에 교미하여, 지연착상된 이후 4-6월 사이에 배발생을 거쳐 출산하는 것으로 알려져 있다(Richardson, 1977; Wallace, 1978; Mori and Uchida, 1981; 윤 등, 2004; Hockman et al., 2009). 흥미로운 현상은 7월 이후 암컷의 체중이 수컷보다 가벼 운 수준으로 떨어진다는 점으로, 아마도 출산 후 체중이 감소하게 되는 것으로 보이며, 이에 따라 암컷의 체중은 암컷-우세적인 형질에서 제외되어야 할 것이다.
A.
B.
Fig. 9. Averages (A) and monthly changes (B) of body weights between males and females of M. schreibersii.
일반적으로 성적 크기 이형(sexual size dimorphism)을 나타내는 암-수의 형질 차이는 번식활동에서의 경쟁, 생식력의 향상을 위한 것으로 제안되고 있다(Ralls, 1976; Nowak, 1994; Lee and Dominguez, 2000; Fenton and Simmons, 2014). 대부분 의 박쥐들은 암컷이 수컷에 비해 더 큰 경향을 보이며, 간혹 암컷과 수컷 사이에 서 극단적인 크기 차이를 나타내기도 한다(Peterson, 1965; Ralls, 1976; Nowak, 1994). 긴가락박쥐의 암컷들처럼 번식기 동안 대규모 공동육아집단을 형성하는 경우에는 후손 양육은 전적으로 암컷이 책임지게 된다. 이에 따라 30일 정도에 성체의 크기로 성장하는 애기박쥐과 박쥐류의 특성상, 암컷 우세적인 신체의 발 달이 Rousettus, Myotis를 비롯한 애기박쥐과 동물들에서 보고된 바 있다(Myers,
1978; Williams and Findley, 1979; Hayssen and Kunz, 1996; Elangovan et al., 2004; Fenton and Simmons, 2014). 하지만, 제주도 긴가락박쥐 집단에서는 ETR을 제외 한 다른 형질들에서 암컷 우세적인 형질들이 발견되지 않는다는 점에서, 향후 여 러 집단들을 대상으로 한 지속적인 연구가 필요하다 하겠다.
Ⅳ
. 참고문헌
김병수, 오홍식. 2005. 제주산 비바리뱀(Sibynophis collaris Gray)의 분포와 서식지. 한국환경생태학회지, 19(4): 342-347. 김유경. 2015. 제주도산 Myotis 속 박쥐류 4종의 계통학적 유연성 연구. 제주대학 교대학원. 박사학위논문. 김태욱, 박수곤, 김유경, 박준호, Pradeep Adhikari, 김가람, 박선미, 이준원, 한상 현, 오홍식. 2015. 작은땃쥐(Crocidura shantungensis)의 외부 형태 및 두 개골 형질의 특성. 환경생물, 33(4): 441-449. 남궁준. 1981. 제주도 용암동굴의 동물상과 그 환경. 한국동굴학회지, 6(7): 17-27. 박수곤, 김유경, 김태욱, 박준호, Pradeep Adhikari, 김가람, 박선미, 이준원, 한상 현, 오홍식. 2015. 제주도에서 박쥐류의 분포에 관한 연구. 환경생물, 33(4): 394-402. 손성원. 1981. 제주도의 익수류상. 경남대학교 논문집, 8: 161-169. 양영민, 홍석욱. 1988. 제주도 용암동굴에 서식하는 익수목의 분포 및 생태에 관 한 연구. 제34회 과학전람회. 생물(기초과학), pp. 1-58. 오대식. 2013. 한국산 주요 박쥐류의 분포현황 및 두개골 형태분석. 인천대학교대 학원. 석사학위논문. 오홍식. 2002. 한라산 국립공원 자연자원조사보고서(포유류, 조류, 양서·파충류편). 국립공원관리공단. 오홍식, 장민호, 김병수. 2007. 한라산 국립공원의 포유류 현황. 한국환경생태학회 지, 21(3): 235-242. 윤명희. 2010. 한국의 척추동물. 대한민국 생물지. 5(1), 박쥐류. 환경부 국립생물 자원관, pp. 36-92. 윤명희, 손성원. 1989. 한국산 박쥐류의 계통분류학적 연구 1. Rhinolophidae의 1 종과 Vespertilionidae의 6종에 대한 분류학적인 재검토 및 한국산 익수류 상의 천이. 한국동물학회지, 32(4): 374-392.
윤명희, 한상훈, 오홍식, 김장근. 2004. 한국의 포유동물. 동방미디어, pp. 36-40. 장민호, 김병수, 박수곤, 김태욱, 오홍식. 2010. 비바리뱀(Sibynophis chinensis)의 뱀 류 섭식에 관한 최초 보고. 한국양서파충류학회지, 2(1): 59-61. 채종보, 박영효. 1988. 제주도산 박쥐류의 채집목록 및 분포조사. 제34회 과학전 람회. 생물(기초과학), pp. 1-36. 한상훈, Dar Fukui, 정철운, 최용근, 김선숙, 전주민. 2011. 산림성 박쥐류의 종다 양성 및 계통연구(I). 국립생물자원관.
Abe, H., N. Ishii, T. Ito, Y. Kaneko, K. Maeda, S. Miura and M. Yoneda. 2005. A Guide to the Mammals of Japan. Tokai University Press, Kanagawa, Japan. Albayrak, I. and S. Coskun. 2000. Geographic variations and taxonomic status of
Miniopterus schreibersii (Kuhl, 1819) in Turkey (Chiroptera: Vespertilionidae). Turkish Journal of Zoology, 24: 125-133.
Altringham, J. D., T. McOwat and L. Hammond. 1998. Bats: Biology and Behavior. Oxford University Press, Oxford, 272pp.
Appleton, B. R., J. A. Mckenzie and L. Christidis. 2004. Molecular systematics and biogeography of the bent-wing bat complex Miniopterus schreibersii (Kuhl, 1817) (Chiroptera: Vespertilionidae). Molecular Phylogenetics and Evolution, 31: 431-439.
Bilgin, R., A. Karataş, E. Çoraman, I. Pandurski, E. Papadatou and J. C. Morales. 2006. Molecular taxonomy and phylogeography of Miniopterus schreibersii (Kuhl, 1817) (Chiroptera: Vespertilionidae), in the Eurasian transition. Biological Journal of the Linnean Society, 87: 577-582.
Dietz, C. and O. von Helversen. 2004. Illustrated identification key to the bats of Europe. Electronic publication ver. 1.0. released 15. 12. Tuebingen and Erlangen, Germany, pp. 40-45.
Dwyer, P. D. 1963. Reproduction and distribution in Miniopterus (Chiroptera). The Australian Journal of Science, 25: 435-436.
Elangovan, V., H. Raghuram, E. Y. S. Priya and G. Marimuthu. 2004. Wing morphology and flight performance in Rousettus leschenaulti. Journal of
Mammalogy, 85(4): 806-812.
Fenton, M. B. and N. B. Simmons. 2014. Bats: A World of Science and Mystery. The University of Chicago Press, Nevramont Publishing Company, New York, pp. 171-197.
Findley, J. S. 1972. Phenetic relationships among bats of the genus Myotis. Systematic Biology, 21(1): 31-52.
Hayssen, V. and T. H. Kunz. 1996. Allometry of litter Mass in bats: maternal size, wing morphology, and phylogeny. Journal of Mammalogy, 77(2): 476-490. Hockman, D., M. K. Mason, D. S. Jacobs and N. Illing. 2009. The Role of Early
Development in Mammalian Limb Diversification: A Descriptive Comparison of Early Limb Development Between the Natal Long-Fingered Bat (Miniopterus natalensis) and the Mouse (Mus musculus). Dev Dyn., 238(4): 965-979.
Hutson, A. M., S. P. Mickleburgh and P. A. Racey. 2001. Microchiropteran Bats: Global Status Survey and Conservation Action Plan. IUCN/SSC Chiroptera Specialist Group. IUCN, Gland, Switzerland and Cambridge, 258pp.
Jo, Y. S., T. W. Kim, B. J. Choi and H. S. Oh. 2012. Current status of terrestrial mammals on Jeju Island. Journal of Species Research, 1(2): 249-256.
Kim, B. S. and H. S. Oh. 2006. A Taxonomic reinvestigation of the Collared Many toothed snake Sibynophis collaris Gray (Reptiles: Serpentes: Colubridae) from Jeju Island, Korea. Integrative Biosciences, 10(3): 121-123.
Koh, H. S., B. G. Yang, H. S. Yoo and T. Y. Chun. 2000. Diversity of mitochondrial DNA cytochrome b gene in roe deer (Capreolus pygargus
tianschanicus) from Jejudo Island, Korea. The Korean Journal of Systematic
Zoology, 16: 169-176.
Kunz, T. H. 1982. Roosting Ecology of Hats. In: Ecology of Bats (T. H. Kunz, ed.). Plenum Press, New York.
Smithsonian Institution Press, Washington, D. C., 533pp.
Kunz, T. H. and L. F. Lumsden. 2003. Ecology of Cavity and Foliage Roosting Bats. In: Bat Ecology (Kunz , T. H. and M. B. Fenton, eds.). The University of Chicago Press, Chicago.
Lee, Jr. T. E. and D. J. Dominguez. 2000. Ametrida Centurio. Mammalian Species, 640: 1-4.
Levenson, H. 1990. Sexual size dimorphism in chipmunks. Journal of Mammalogy, 71: 161-170.
Macdonald, D. W. and P. Barrett. 1993. Mammals of Britain and Europe. Harper Collins, London.
Maeda, K. 1982. Studies on the classification of Miniopterus in Eurasia, Australia and Melanesia. Honyurui Kagaku. Supplement 1. Mammalian Science, pp 1-176.
Mahmood-ul-Hassan, M. and M. Salim. 2015. Two new bat species (Chiroptera: Mammalia) for Pakistan: Miniopterus fuliginosus and Myotis formosus. Mammalia, 79(1): 125-129.
Mori, T. and Uchida, T. A. 1981. Ultrastructural observations of ovulation in the Japanese long-fingered bat, Miniopterus schreibersii fuliginosus. Journal of the Society for Reproduction and Fertility, 63: 391-395.
Mori, T. 1933. On two bats from Korea. J. Chosen N. H. Soc. 16: 1-5.
Motokawa, M., L. K. Lin, M. Harada and S. Hattori. 2003. Morphometric geographic variation in the Asian Lesser White-Toothed Shrew Crocidura shantungensis (Mammalia, Insectivora) in East Asia. Zoological Science, 20(6): 789-795. Myers, P. 1978. Sexual dimorphism in size of vespertilionid bats. The American
Natulaist, 112(986): 701-711.
Nowak, M. R. 1994. Walker's Bats of the World. Johns Hopkins University Press, pp. 63–64.
Norberg, U. M. and J. M. V. Rayner. 1987. Ecological morphology and flight in bats (Mammalia; Chiroptera): wing adaptations, flight performance, foraging
strategy and echolocation. Philosophical Transactions of the Royal Society B, 316(1179): 335-427.
Oh, D. J., T. W. Kim, M. H. Chang, S. H. Han, H. S. Oh and S. J. Kim. 2013. Migration route estimation of the Jeju striped field mouse Apodemus agrarius
chejuensis (Rodentia, Muridae). Mitochondrial DNA, 24(2): 137-144.
Oh, H. S. and T. Mori. 1998. Taxonomic re-examination of the striped field mouse,
Apodemus agrarius coreae and A. a. chejuensis: Evidence from crossbreeding
experiments (Mammalia: Rodentia). Journal of the Faculty of Agriculture, Kyushu University, 43(1-2): 143-151.
Oh, H. S., Y. Yoshinaga, T. Kaneko, H. Iida and T. Mori. 2003. Taxonomic re-examination of the Apodemus agrarius chejuensis, comparing external and cranial morphological characters among four Asian Apodemus species. Journal of the Faculty of Agriculture, Kyushu University, 47(2): 373-386.
Patterson, B. D., M. R. Willig and R. D. Stevens. 2003. Trophic Strategies, Niche Partitioning, and Patterns of Ecological Organization. In: Bat Ecology (Kunz, T. H. and M. B. Fenton, eds.). The University of Chicago Press, Chicago, pp. 536-579.
Peterson, R. L. 1965. A review of the genus Ametrida, family Phyllostomidae. Contributions of the Royal Ontario Museum, Life Sciences, 65: 1-13.
Puechmaille. S. J., B. Allegrini, P. Benda, K. Gürün, J. Srámek, C. Ibañez, J. Juste and R. Bilgin. 2014. A new species of the Miniopterus schreibersii species complex (Chiroptera: Miniopteridae) from the Maghreb region, North Africa. Zootaxa, 3794(1): 108-124.
Ralls, K. 1976. Mammals in which females are larger than males. The Quarterly Review of Biololgy, 51: 245-276.
Richardson, E. G. 1977. The biology and evolution of the reproductive cycle of
Miniopterus schreibersii and M. australis (Chiroptera: Vespertilionidae).
Mammalogy, 42: 152-159.
Tian, L. X., B. Liang, K. Maeda, W. Metzner and S. Y. Zhang. 2004. Molecular studies on the classification of Miniopterus schreibersii (Chiroptera: Vespertilionidae) inferred from mitochondrial cytochrome b sequences. Folia Zoologica, 53: 303-311.
Walker, E. P. 1975. Mammals of the World. Vol I. John Hopkins Universotu Press, Baltimore.
Wallace, G. I. 1978. A histological study of the early stages of pregnancy in the bent-winged bat (Miniopterus schreibersii) in north-eastern New South Wales, Australia (30°27'S). Journal of Zoology, 185: 519-537.
Wallin, L. 1969. The Japanese Bat Fauna: A Comparative Study of Chorology, Species Diversity and Ecological Differentiation. Almqvist and Wiksells Boktryckeri AB.
Williams, D. F. and J. S. Findley. 1979. Sexual size dimorphism in vespertilionid bats. The American Midland Naturalist, 102(1): 113-126.
Wilson, D. E. and D. M. Reeder. 2005. Mammal Species of the World: A Taxonomic and Geographic Reference. Third Edition. The Johns Hopkins University Press, Baltimore.
Yoshiyuki, M. 1989. A Systematic Study of the Japanese Chiroptera. National Science Museum, Tokyo, Japan.
Zidarova, S. 2015. Is there sexual size dimorphism in shrews? A case study of six european species of the family Soricidae. Acta Zoologica Bularica, 67(1): 19-34.
<Abstract>
External Morphological Characteristics of
Miniopterus schreibersii in Jeju Island, Korea
Yeon-Ha An
Major in Biology Education, Graduate School of Education, Jeju National University (Supervised by Professor Hong-Shik Oh)
This study was undertaken with the purpose of identifying external morphological characteristics of Miniopterus schreibersii on the Jeju Island and providing information necessary for the protection of habitats and biological diversity. The field research was conducted from March 2014 to April 2016 in a total of 35 areas on the Jeju Island, including lava tubes (n=13), cave encampments (n=18), sea caves (n=2), and natural cavelets (n=2). According to the research results and the literature review, the inhabitation of M. schreibersii was confirmed in at least 11 areas. M.
schreibersii bats were mostly in the form of populations of over 100 individuals;
however, separate individuals were occasionally observed as well. While their habitats were lava tubes, cave encampments, and forests, large-scale populations were found in the eastern and western parts of the Jeju Island. Among the measured morphological characters of the Jeju population, most of them, except for the head-body length, were similar to those of the Korean Peninsula population. By contrast, there were remarkable differences in geographical morphological variation between the two Korean populations (the Jeju population and the Korean Peninsula population) and the Japanese population. The sexual size dimorphism in the Jeju population exhibited the significant differences (P<0.05) in body weight (BW), ear-length/tragus-length ratio (ETR), humerus length (Hum), and tibia length (Tib). Among these, BW and ETR were female-biased, but this expression of BW was
considered to occur due to the increased body weight during the gestation period. On the contrary, Hum and Tib characteristics were male-biased. Through the research results, M. schreibersii’s various habitats on the Jeju Island, including lava tubes, cave encampments, and forests, were confirmed. In terms of external morphological characters, the head-body length of the Jeju population was greater than that of the Korean Peninsula population, but smaller than that of the Japanese population. The sexual size dimorphism in the Jeju population was also found as well. These results provide important information that should necessarily be considered in the planning of the preservation of natural or artificial caves and the development of eco-tourism programs on the Jeju Island. In addition, our results imply that comprehensive studies on habitat distribution, geographical morphological variation among populations, and sexual size dimorphism of M. schreibersii need to be conducted with the collection of sufficient data from various regions in East Asia, including the Korean Peninsula, Japan, and China. Considering the fact that this study is the first to demonstrate M.
schreibersii in South Korea at the level of populations, whose ecological
characteristics are little known yet, our results will contribute to better understanding of population characteristics of M. schreibersii.