Oleanolic acid 및 그 유도체가 MC3T3-E1 조골세포주의 분화에 미치는 효과
김세원*·이창호**·정희경***·조성신***·이홍기***·박용순*†
*한양대학교 식품영양학과, **한양대학교 의과대학 약리학교실, ***한양대학교 의생명공학과
Effects of Oleanolic Acid and its Derivatives on the Differentiation of MC3T3-E1 Osteoblastic Cell
Se Won Kim*, Chang Ho Lee**, Hee Kung Jung***, Sung Sin Jo***, Hong Ki Lee*** and Yong Soon Park*†
*Department of Food and Nutrition, Hanyang University, Seoul 133-791, Korea.
**Department of Pharmacology, Hanyang University, Seoul 133-791, Korea.
***School of Biomedical Science & Engineering, Hanyang University, Seoul 133-791, Korea.
ABSTRACT : Ursolic acid, triterpenoid compound has been shown to stimulate osteoblast differentiation and enhance bone formation. In the present study, we examined the effects of similar triterpenoid compounds, oleanolic acid (OA) and its derivatives, such as oleanolic acid acetate (OAA) and oleanolic acetate methyl ester (OAM) on the bone formation in MC3T3-E1 osteoblast cells. We determined cellular proliferation, alkaline phosphatase (ALP) activity, mineralization, and expression of osteoblast specific genes and mitogen activated protein kinase phosphorylation. Treatment of 0.1-10µM OA, OAA, and OAM increased cellular proliferation, but not significantly increased as compared with dimethyl sulfoxide (DMSO). OA, OAA, and OAM at 5uM concentration enhanced ALP expression, mineralization, and osteocalcin (OCN) mRNA level. In conclusion, OA and its derivatives stimulated the osteoblast differentiation by increasing ALP, mineraliza- tion, and OCN mRNA expression. However, there were no significantly difference on osteoblast differentiation among treat- ment of OA, OAA, and OAM.
Key Words : Oleanolic Acid, Oleanolic Acid Derivatives, Osteoblast Specific Ggene, Differentiation, Proliferation, MC3T3-E1 Cell
서 언
인체의 뼈는 골 형성과 재형성 과정을 통하여 그 항상성이 유지된다 (Parfitt, 1994). 조골세포에서 type I collagen (COLI), alkaline phosphatase (ALP), osteopontin (OPN), Runx2, osteocalcin (OCN) 등과 같은 골 관련 유전자의 연속 적인 발현이 골형성 과정에 관여한다 (Owen et al., 1990).
ERK, JUNK, p38를 포함하는 mitogen-activated protein kinase (MAPKs)는 조골세포 분화과정에서 BMP2에 의해 활 성화되는 것으로 알려진 주요 non-Smad 신호전달 경로이다 (Kozawa et al., 2002). MAPKs 경로의 활성화는 OPN 과 COLI 유전자의 발현 유도를 조절하고 (Hipskind and Bilbe, 1998), ERK와 JNK의 활성화는 ALP 활성과 골 석회화를 유 도한다고 보고되었다 (Hanai et al., 2006).
조골세포 활성의 약화로 골형성이 감소되거나 파골세포의 활성도가 강화되어 골 흡수가 증가되는 경우, 또는 두 가지
요소가 동시에 작용되는 경우에 골다공증이 발생한다 (Lee et al., 2008). 특히 골다공증은 폐경기 여성에 있어서 주된 건강 문제로 인식되어 왔으며 (Zhang et al., 2005), 이는 폐경 후 에스트로겐 감소로 골 소실이 증가되기 때문이다 (Ebeling et al., 1996). 그 외의 골다공증 원인으로 불충분한 칼슘 섭취, 장내 칼슘 흡수력의 저하, 비타민 D부족으로 인한 이차적 부 갑상선 호르몬 항진과 골 소실 증가 (Lips, 2001), 유전적 요 인 (Christian et al., 1989) 등이 있다.
골다공증 치료요법으로 골 동화 요법과 골 흡수 억제 요법 이 사용되고 있다. 골 동화 요법은 불화나트륨 (Kleerekoper and Mendlovic, 1993), 간헐적인 parathyroid hormone (PTH) 및 그 유사체 투여요법 (Marumo and Satio, 2011) 등이 보 고되었고, 골 흡수 억제 요법은 에스트로겐 요법, 칼시토닌 요 법, 칼슘 보충제 및 비타민 D 유도체 요법 등이 있다. 골 폐 경기 여성의 골다공증 치료에는 에스트로겐 투여요법이 효과 적인 치료법으로 인식되고 있으나, 유방암 발생 및 혈전 형성
†Corresponding author: (Phone) +82-2-2220-12055 (E-mail) [email protected]
Received 2011 November 22 / 1st Revised 2011 December 13 / 2nd Revised 2011 December 20 / 3rd Revised 2011 December 22 / Accepted 2011 December 22
등의 부작용 유발, 투여시기 조절의 어려움과 유방암, 자궁암, 간질환 등을 가진 특정 환자에 대한 적용상의 어려움 등으로 인하여 그 사용에 있어 제약이 따른다 (Anderson and Grancer, 1998). 골 흡수 억제제는 이미 진행된 골 소실을 완 전히 회복할 수 없어 골다공증을 완전히 예방할 수 없다 (Delmas, 2002). 따라서 골다공증의 예방과 치료를 위하여 골 형성을 촉진시키는 방법에 관한 연구가 활발하게 이루어지고 있다. 특히, 생약제에 대한 효능 및 효과를 근거로 하여 골조직 재생능력에 미치는 영향 등에 대한 접근이 다양하게 시도되고 있다 (Cho et al., 1996; Boonen et al., 1997; Kwak et al., 2006; Kwon et al., 2011).
Oleanolic acid (OA)는 triterpenoid계 화합물로서 유리산 또 는 배당체의 형태로, 올리브 잎 (Olea europaea)에서 최초로 분리되었고, 쓴풀 (Swertia japonica), 자바애플 (Eugenia jambos), 용담 (Gentiana lutea) 등과 같은 약용식물에서 발견 되는, 자연계에 널리 존재하는 생리활성 물질이다 (Jie, 1995).
OA는 상대적으로 독성이 없으며, 간 보호효과 (Liu et al., 2008), 항염증효과 (Dinkova-Kostova et al., 2005), 항암효과 (Zhou et al., 2011), 저혈압효과 (Bachhav et al., 2011), 저 혈당효과 (Musabayane et al., 2010) 등의 약리적 효과가 보 고되었다. oleanolic acid acetate (OAA), oleanolic acetate methyl ester (OAM)는 OA의 유도체로서 같은 triterpenoid계 화합물이다 (Fig. 1).
OA의 조골세포 분화 작용에 대한 연구는 없었으나, OA와 유사한 고리구조와, 약리적 효과를 가지고 있는 triterpenoid계 화합물인 ursolic acid에 대한 연구는 보고된 바 있다 (Jie, 1995). Lee 등 (2008)에 의하면, MC3T3-E1 세포에서 ursolic acid가 MAP kinase와 그 외 전사인자를 활성화시킴으로써 조 골세포의 분화를 촉진시키고, 골 형성을 유도하였다고 한다.
또한 OA와 유사한 고리구조를 가지고 있는 monoter- penoid계 천연 화합물인 3-Carene도 조골세포의 무기질화를 촉 진시킨다고 하였다 (Jeong et al., 2008). MC3T3-E1 조골세 포는 생쥐 두개골에서 유래한 것으로 뼈 형성 과정 중 세포의 증식, 분화, 석회화 과정이 골아세포와 유사한 대사적 특징을 가지고 있어, 조골세포 분화실험에 많이 사용되고 있다 (Sudo et al., 1983). 따라서 본 연구에서 OA 및 그 유도체인 OAA, OAM이 MC3T3-E1 조골세포의 활성과 분화에 미치는 효과를 평가하였다.
재료 및 방법
1. 세포배양
MC3T3-E1 세포 (CRL-2953, American Type Culture Collection, Manassas, VA, USA)는 증식유도 배지 (growth medium, GM; α-minimal essential medium (LM 00853, WelGENE, Daegu, Korea); 10% fetal bovine serum (FBS;
S001-01, WelGENE, Daegu, Korea); 100 unit/㎖, 100 g/㎖
penicillin/streptomycin (LS202-02, WelGENE, Daegu, Korea)) 에서 배양하였다. 세포 분화를 유도하기 위하여, confluent 상 태로 배양된 세포를 분화유도 배지 (differentiation medium, DM; α-minimal essential medium (LM 00801, WelGENE, Daegu, Korea); 10% FBS; penicillin/streptomycin 100 unit/
ml, 100㎍/㎖; ascorbic acid 50 ㎍/㎖ (Sigma Co, St Louis, MO, USA); 10 mM β-glycerophosphoric acid diso- dium salt (Sigma Co, St Louis, MO, USA))에서 배양하였 다. 약물의 분화 효과를 검정하기 위해 세포에 dimethyl sulfoxide (DMSO), OA, OAA, OAM를 각각 처리 하였다.
세포는 37℃의 5% CO2 배양기에서 배양 하였으며, 배지는 3 Fig. 1. Structure of oleanolic acid, oleanolic acid 3-acetate and oleanolic acetate methyl ester.
일 간격으로 교환해 주었다. OA, OAA, OAM 등은 포도씨 (grapestone)에서 추출하였고, 서울대학교 천연물과학연구소 한 용남 교수님으로부터 제공받았으며, DMSO에 녹여 사용하였다.
2. 조골세포 증식률 측정
DMSO, OA, OAA, OAM의 농도 별 처리에 따른 조골세 포의 성장 속도는 Suda 등 (1999)의 방법에 따라 MTT colorimetric assay를 이용하여 측정하였다. 96 well plate에 50,000 cell/㎠의 농도로 분주한 후, DMSO에 용해된 각 시료 를 최종농도 0.1, 1, 2.5, 5, 10 µM로 처리하여 3일 동안 배 양하였다. MTT시약 (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolim bromide, M2128, Sigma Co, St Louis, MO, USA)을 각 well에 20 µL (5 ㎎/mL in PBS)씩 첨가한 후 4 시간 더 배양하였다. 그 결과 생성된 불용성의 formazan 결정 을 용해시키기 위하여 배지를 제거하고 DMSO를 100 µL 첨 가하여 실온에서 20분 동안 흔든 후, ELISA reader (Bio Rad iMark Microplate Reader, Bio-Rad, Seoul, Korea)를 이 용하여 570 ㎚에서 흡광도를 측정하였다.
3. ALP 염색에 의한 조골세포 활성도 분석
ALP 염색 정도를 확인하기 위하여 MC3T3-E1 세포를 50,000 cell/㎠의 농도로 96 well plate에 분주하여 4-5일간 배 양한 다음, 증식유도 (GM), 분화유도 (DM) 배지에서 DMSO 및 0.1, 1, 2.5, 5, 10 µM 농도의 OA, OAA, OAM을 처리 하여 14일간 배양하였다. 염색은 ALP kit (Sigma Co, St Louis, MO, USA)를 이용하여 측정하였다. 배지를 제거한 후 증류수로 한번 세척한 후 고정액 (citrate-acetone-formal- dehyde)을 첨가하여 약 30초간 실온에서 고정시켰다. Diazo- nium solution (sodium nitrite: FRV-alakline: 증류수: naphthol AS-BI alkaline solution = 1 : 1 : 45 : 1)을 첨가하여 약 15분간 실온에서 방치한 다음, hematoxylin solution으로 2분간 염색 한 뒤 현미경으로 관찰하였다.
4. Alizarin-red 염색법에 의한 석회화 형성도 검사
ALP염색법과 동일하게 MC3T3-E1 세포를 50,000 cell/㎠
농도로 96 well plate에 분주하여 4-5일간 배양한 다음, GM, DM배지에서 DMSO 및 0.1, 1, 2.5, 5, 10 µM 농도의 OA, OAA, OAM을 처리하여 14일간 배양하였다. PBS로 2번 세척 한 뒤 Alizerin red solution (40 mM, pH 4.2)을 처리하여 실온에서 1시간 동안 고정시킨 후, 현미경으로 관찰하였다 (Byers and García, 2004).
5. ALP 효소활성 측정
MC3T3-E1 세포를 50,000 cell/㎠ 농도로 24 well plate에 분주하여 4-5일간 배양한 다음, GM과 DM 배지에서 DMSO
및 0.1, 1, 2.5, 5, 10 µM 농도의 OA, OAA, OAM을 처리 하여 7일 및 14일간 배양하였다 (Cheng et al., 2000). 배양 이 끝난 세포는 배양액을 제거한 후 1x Tris buffered saline (50 mM Tris, pH 7.4, 150 mM NaCl)용액으로 3회 세척한 후, 각 well에 100 µL의 lysis용액 (10 mM Tris, pH 7.4, 0.5 mM MgCl2, 0.1% Triton X-100)을 가하고, 세포를 scraper로 수집한 후, 초음파 분산기 (Fisher Scientific, Pittsburgh, PA, USA)를 이용하여 40% 강도로 음파처리 하였 다. 세포를 4℃, 12,000xg에서 10분간 원심분리 하였으며, 96 well plate에 상층액을 20 µL씩 각각 분주한 후, 100 µL의 working assay solution (p-nitrophenyl phosphate 6.7 mM, 2.0 mM MgC12, 100 mM carbonate Buffer, pH 9.8)을 넣고 37℃에서 15분간 반응시켰다. 반응 후 NaOH 용액 (200 mM)을 80 µL씩 넣고 405 ㎚에서 흡광도를 측정하였다. 단백 질량은 bovine serum albumin을 이용한 Bradford법 (1976)을 이용하여 정량하였고, ALP 활성은 unit/mg protein으로 표시 하였다.
6. RNA 추출과 역전사 중합 효소 연쇄반응 (Reverse Transcription Polymerase Chain Reaction, RT-PCR)
RNA는 RNeasy mini kit (74104, Qiagen, Valencia, CA, USA)를 이용하여 추출하였으며, 분광광도계 (NanoDrop 1000 Spectrophotometer V3.7, Thermo Fisher Scientific, Wil- mington, DE, USA)을 이용하여 정량하였다. cDNA는 1 ㎍의 RNA와, oligo (dT), M-MLV Reverse Transcriptase (EBT- 1208, Elpis Bio, Daejeon, Korea)를 이용하여 최종 20 µL 에서 합성하였으며, 역전사 과정은 70℃에서 10분, 42℃에서 1시간 30분, 94℃에서 10분 동안 수행하였다. PCR 과정은 합 성한 1 µL의 cDNA와 HiPi Plus Thermostable DNA polymerase (EBT-1010; Elpis-Bio, Daejeon, Korea)을 사용하 였다. PCR에 사용한 모든 염기서열 (Genotach, Daejeon, Korea)은 Table 1에 정리하였다. PCR 조건은 94℃, 30초; 61
℃, 30초; 72℃, 30초로 설정하여, beta-actin의 경우 27 cycle, OCN은 30 cycle, 그 외 OPN, Runx2, CoL1은 모두 25 cycle로 수행하였다. 반응 산물은 2% agarose gel에서 전 기영동 후 ethidum bromide로 염색하여 UV lamp 하에서 확 인하였다.
7. Western blot analysis
세포를 RIPA완충액 (50 mM Tris-HCl pH 7.4, 150 mM NaCl, 1 mM EDTA, 1% NP-40, 0.25% sodium-deoxy- cholate)에 protease inhinitor cocktail (Roche, Mannheim, Germany)과 phosphatase inhibitor 1 (p2580; Sigma, St Louis, MO, USA)과 phosphatase inhibitor 2 (p5726; Sigma, St Louis, MO, USA)를 혼합한 용액으로 용해시킨 후,
14,000× g로 4℃에서 20분간 원심분리 하였다. 단백질 정량 후, 20-40 ㎍의 단백질을 10% SDS polyacrylamide gel에서 전기영동을 시행하였고, transfer 완충액 (25 mM Tris, 192 mM glycine, 20% methanol)을 넣고, 300 mA에서 2시간 동 안 nitrocellulose membrane으로 전이시켰다. Membrane을 5% skin milk가 포함된 TBST (Tris-buffered saline: 20 mM Tris-HCl, pH 7.5, 137 mM NaCl; 0.1% Tween 20) 용액에서 1시간 bloking 시킨 후, TBST용액에 각각의 1차 항 체 (beta-actin, 1 : 10,000; JNK, 1 : 5000; pJNK, ERK, pERK, p38, p-p38, 1 : 1000)를 첨가하여 4℃에서 12시간 이상 반응 시켰다. Membrane에 5% skin milk 용액과 2차 항체를 1차 항체와 같은 농도로 첨가하여 상온에서 1시간 동안 반응시켰 다. Membrain을 PicoEPD Western Reagent (EBP-1073;
Elpis Bio, Daejeon, Korea)용액으로 감광시켜 ChemiDoc XRS System (Bio-rad, Seoul, Korea)를 이용하여 밴드를 확 인하였다.
ERK, JNK, p-JNK, p38, p-p38, anti-goat IgG 항체는 Santa Cruz Biotechnology (Santa Cruz, CA, USA)에서,
beta-actin 항체는 Sigma (St Louis, MO, USA)에서, p-ERK 항체는 Cell Signaling technology (Danvers, MA, USA)에서, goat-anti-mouse와 goat-anti-rabbit 항체는 Jackson Immune- Research (Westgrove, PA, USA)에서 구입하였다.
8. 통계처리
본 실험결과는 최소한 3회 이상의 실험을 걸쳐 평균 ± 표준 편차로 표시하였다. 자료분석은 SPSS 18.0 software (SPSS;
Chicago, IL, USA)를 이용하여 통계분석을 수행하였다. 유의 적 차이를 확인하기 위해 one-way ANOVA를 수행하였고, 군 별 차이는 Duncan의 다중검정법을 이용하여 시료간 유의적 차이를 p < 0.05에서 검증하였다.
결과 및 고찰
1. OA와 그 유도체가 조골세포 증식에 미치는 효과 조골세포주의 증식은 OA, OAA, OAM을 0.1, 1, 2.5, 5, 10 µM로 처리하였을 때, GM대조군에 비하여 유의하게 높았 Table 1. Primer sequences used in this study.
Target gene Forward (5'-3') Reverse (5'-3')
β-Actin GTGGGGCGCCCCAGGCACCA TCCTTAATGTCACGCACGATTTC
Runx2 GCCGGGAATGATGAGAACTA GGACCGTCCACTGTCACTTT
OPN ATTTGCTTTTGCCTGTTTGG CTCCATCGTCATCATCATCG
CoL I GCGAAGGCAACAGTCGCT CTTGGTGGTTTTGTATTCGATGAC
OCN CTCACAGATCCCAAGCCCA CCAAGGTAGCGCCGGAGTCT
Fig. 2. Effect of oleanolic acid oleanolic acid (OA) and its derivatives (OA), oleanolic acid acetate (OAA) and oleanolic acetate methyl ester (OAM) on the proliferation of MC3T3-E1 cells by the MTT assay. Data are expressed as a percentage of growth medium (GM). Bars represent means±SD, and different letters are significantly different at P<0.05.
으나, DMSO 대조군에 비해서는 유의하게 증가하지 않았다.
OA, OAA, OAM 약물처리군 간에도 유의한 차이가 없었다
(Fig. 2). Lee 등 (2008)은 0-20 µM의 ursolic acid를 조골세 포주에 처리한 결과, 약물을 처리하지 않았을 때 보다 증식률 Fig. 3. Effect of 0.1-10 µM oleanolic acid and its derivatives on osteoblast differentiation by Alkaline phosphatase (ALP) and
mineralization by Alizerin red staining. Cells were treated with dimethyl sulfoxide (DMSO), oleanolic acid (OA; A), oleanolic acid acetate (OAA; B), and oleanolic acetate methyl ester (OAM; C) in growth medium (GM), differentiation medium (DM) for 14 day (× 100).
이 낮았으며, 10 µM 이상에서 세포성장이 급격히 억제되었다 고 보고하였다. 본 연구에서는 DMSO대조군과 비교했을 때 OA와 그 유도체의 증식유도 효과를 확인할 수 없었으나, GM 대조군과 비교하여 증식률이 유의하게 높은 것으로 보아 ursolic acid 보다 세포독성이 적은 것을 확인할 수 있었다.
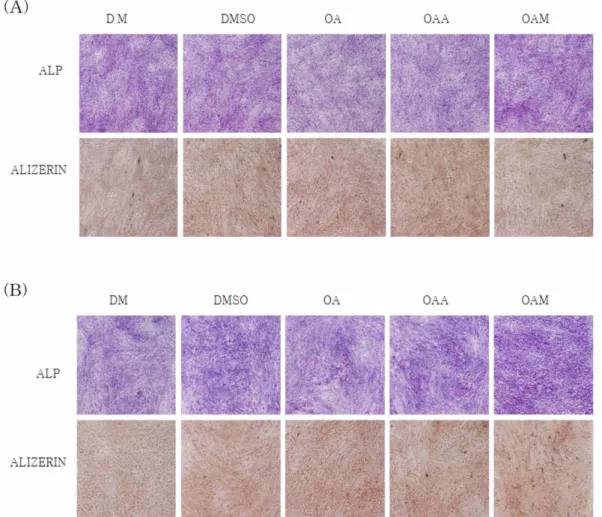
2. ALP와 Alizerin 염색법에 의한 효소발현과 석회화 형성 1) ALP 염색법에 의한 효소발현
OA, OAA, OAM을 0.1, 1, 2.5, 5, 10 µM로 14일간 처리 하여 ALP효소의 발현을 확인한 결과, 10 µM의 OA를 처리하 였을 때, GM과 DM 조건에서 모두 세포가 사멸하였다 (Fig.
3). GM 조건과는 다르게, DM 조건에서는 DMSO 대조군, OA, OAA, OAM 약물처리군 모두에서 붉은색으로 염색된 ALP 효소발현이 증가하였으나, 약물처리군 간에 ALP 발현의 차이는 없었다. 따라서, 세표가 사멸하지 않는 최대농도인 5 µM의 OA, OAA, OAM으로 실험을 진행하였다.
2) Alizerin-red 염색법에 의한 석회화 형성
OA, OAA, OAM을 0.1, 1, 2.5, 5, 10 µM 농도로 14일간 배양한 다음 Alizerin-red염색법에 의한 석회화 형성 정도를 확인한 결과, ALP 염색 결과와 같이 10 µM의 OA를 처리하 였을 때, GM과 DM 조건에서 모두 세포가 사멸하였다 (Fig.
3). GM 조건에서는 모든 군에서 석회화 형성을 확인할 수 없 었으나, DM 조건에서는 석회화가 형성되었다. 그러나 약물처 리군 간에 석회화 형성의 차이는 없었다.
3) 배양 7일 및 14일째 ALP 효소발현과 석회화 형성 ALP 염색결과, 배양 7일째에는 DM과 DMSO대조군에 비 해 모든군에서 ALP의 효소가 발현되지 않았으나, 14일째에는 OA, OAA, OAM 약물처리군에서 ALP 효소발현이 증가하였 다 (Fig. 4). 특히 OAA와 OAM 에서 ALP 효소발현이 높게 나타났다. Alizerin 염색결과, 배양 7일부터 석회화 결정이 형 성되기 시작하였으나 약물처리군 간의 차이는 없었으며, 14일
Fig. 4. Effect of 5 µM oleanolic acid and its derivatives on osteoblast differentiation by Alkaline phosphatase (ALP) and mineralization by Alizerin red staining. Cells were treated with dimethyl sulfoxide (DMSO), oleanolic acid (OA), oleanolic acid acetate (OAA), and oleanolic acetate methyl ester (OAM) in differentiation medium (DM) for 7 day (A) and 14 day (B) (× 100).
째에 OA, OAA, OAM 약물처리군에서 DM과 DMSO대조군 보다 석회화 형성이 증가하였다 (Fig. 4).
Jeong 등 (2008)의 연구에서 3-carene을 0-10 µM 농도로 처리한 후 ALP와 Alizerin으로 염색하였을 때, 5 µM 이하에 서는 조골세포의 무기질화가 촉진되었으나, 10 µM에서 조골 세포가 사멸하였다. 따라서 OA와 유사한 고리구조를 가진 화 합물은 높은 농도에서 세포독성이 있는 것으로 보여 진다.
Lee 등 (2008)의 연구에서는 ursolic acid 처리 시 배양 9일 째에 ALP효소발현이 증가하였고, 15일째에 무기질화가 관찰 되었다. Jeong 등 (2008)의 연구에서도 3-carene 처리 시 배 양 9일째에 ALP 효소발현이 증가하였고, 15일째에 석회화 형 성이 증가하였다.
3. ALP 효소활성 분석
배양 7일째에는 ALP 활성이 OA, OAA, OAM 처리군에서 DM 대조군 보다 유의하게 (p = 0.001) 높았으며, OAM 처리 군에서는 DMSO 대조군보다 유의하게 높았다 (Fig. 5). 배양 14일째에는 ALP 활성이 OA, OAA, OAM 처리군에서 DM 대조군보다 유의하게 높았으나, DMSO 대조군과 비교하였을 때 유의한 차이는 없었다. 약물처리군 간에도 유의한 차이는 없었다. 배양 7일째에 ALP 활성이 DMSO 대조군에서 DM 대조군보다 유의하게 높았으며, 14일째에도 유의하지는 않았 으나 증가하는 경향을 보였다. Cheung 등 (2006)의 연구에 의하면 MC3T3-E1 세포에 1%의 DMSO 처리 시 분화를 촉 진시킨다고 하였다. 그러나 본 연구에서는 0.25%의 DMSO 를 사용했기 때문에 위 연구의 결과와 차이가 있었다.
Lee 등 (2008)의 연구에서는 ursolic acid 5 µM 처리 시 9 일째에 대조군보다 ALP활성이 높았으며, 본 연구에서의 7일
째 결과와 유사하였다. 반면에 Park 등 (2005)의 연구에서 미 역취 뿌리 메탄올 추출물을 3-27일간 처리 시 18일째부터 ALP 활성이 급격히 유도되었으며, Maeda 등 (2001)의 연구에 서도 simvastatin을 26일간 처리 시 18일째에 ALP활성이 가 장 높았다. 본 연구에서도 7일째보다 14일째에 ALP활성이 증 가하였으며, 14일 이상의 배양기간이 ALP 활성에 영향을 주 는 요소라 사료된다.
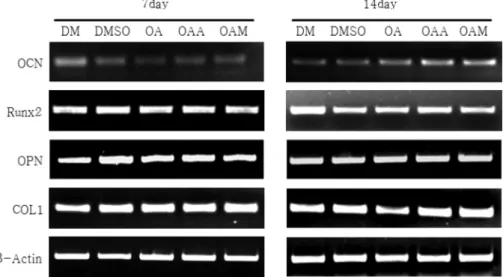
4. RT-PCR을 이용한 유전자 발현 분석
배양 7일째에는 DM 및 DMSO대조군과 OA, OAA, OAM 약물처리군 간에 CoLI, OPN, Runx2, OCN 등에서 유 전자 발현의 차이가 나타나지 않았다. 배양 14일째에는 OA, OAA, OAM처리군에서 OCN 유전자 발현이 증가하였고, 유 전자 발현의 정도는 OA > OAA > OAM 순으로 높았다 (Fig.
6). 그러나 OPN, Runx2, COLI 유전자 발현은 약물처리군 간 에 차이가 없었다.
OCN의 발현은 골이 석회화되어 결절을 형성하는 시기와 일 치하며, 이는 골기질의 석회화를 촉진시키는 요인으로 작용하 는 것으로 알려져 있다 (Hauschka et al., 1989). OPN은 OCN과 달리 조골세포 분화의 초기와 석회화기에 나타나며 (Owen et al., 1990), 조골세포에서 Runx2 발현이 증가할때 COLI과 OCN의 발현이 유도되어 뼈의 무기질화를 촉진한다 (Byers and Garcia, 2004). Lee 등 (2008)의 연구에서 5 µM ursolic acid 처리 시 OPN, OCN, COLI 유전자 발현이 대조 군보다 높았다.
5. Western blot을 이용한 단백질 발현 분석
배양 14일째에 모든군에서 p38의 인산화는 관찰되지 않았 Fig. 5. Effect of oleanolic acid and its derivatives on osteoblast differentiation by Alkaline phosphatase (ALP) activity. Cells were treated with dimethyl sulfoxide (DMSO), 5 µM oleanolic acid (OA), oleanolic acid acetate (OAA), and oleanolic acetate methyl ester (OAM) in differentiation medium (DM) for 7 day and 14 day. Bars represent means ± SD, and different letters are significantly different at P < 0.05.
으나, ERK는 OAA와 OAM 처리 시, JNK는 OAM 처리 시 인산화가 감소하였다 (Fig. 7). Lee 등 (2008)의 연구에서
2.5-5 µM ursolic acid가 JNK와 p38의 인산화를 유도하였으며, 1 µM ursolic acid가 ERK의 인산화를 유도하였다. 그러나 Jeong 등 (2008)의 연구에서는 3-carene 처리 시 ERK, JUNK, p38 MAPKs의 인산화가 관찰되지 않았다. 조골세포의 분화를 유도하는 데 있어 MAPKs외에도 protein kinase C 및 D, phosphatidyl inositol 3 kinase 등 여러 non-Smad 신호전 달 경로를 활성화 시켜 그 효과를 나타낸다 (Jun et al., 2009). 따라서 MAPKs외에도 조골세포 분화에 관여하는 BMP 신호전달 경로에 대한 연구 또는 OA 및 그 유도체가 골의 석회화와 ALP활성에 미치는 영향을 알아보기 위하여 처리기 간을 늘리는 연구가 수행되어야 할 것으로 사료된다.
결론적으로, OA 및 그 유도체는 ALP효소발현과 석회화 형 성, OCN의 유전자 발현을 유도하여 조골세포의 분화를 촉진 하였으나, OA와 그 유도체간에 분화효과의 차이는 확인할 수 없었다.
감사의 글
본 연구는 농촌진흥청 (Agricultural R&D 15th Agenda, 200901OFT061740279와 PJ006983201005) 지원으로 수행되었 습니다.
LITERATURED CITED
Anderson JJ and Garner SC. (1998). Phytoestrogen and bone.
Bailliere's Clinical Endocrinology and Metabolism. 12:543-557.
Bachhav SS, Patil SD, Bhutada MS and Surana SJ. (2011).
Oleanolic acid prevents glucocorticoid-induced hypertension in Fig. 6. Effect of oleanolic acid and its derivatives on mRNA expression of Runx2, osteopontin, osteocalcin and type-1 collagen in MC3T3-E1 cells. Cells were treated with dimethyl sulfoxide (DMSO), 5 µM of oleanolic acid (OA), oleanolic acid acetate (OAA), and oleanolic acetate methyl ester (OAM) in differentiation medium (DM) for 7 day and 14 day.
Fig. 7. Effect of oleanolic acid and its derivatives on activation of mitogen-activated protein kinase (JNK, ERK and p38). Cells were treated with dimethyl sulfoxide (DMSO), 5 µM of oleanolic acid (OA), oleanolic acid acetate (OAA), and oleanolic acetate methyl ester (OAM) in differentiation medium (DM) for 14 day.
rats. Phytotherapy Research. 25:1435-1439.
Byers BA and García AJ. (2004). Exogenous Runx2 expression enhances in vitro osteoblastic differentiation and mineralization in primary bone marrow stromal cells. Tissue Engineering.
10:1623-1632.
Boonen A, Broos P and Deqeker J. (1997). The prevention of treatment of age related osteoporosis in the elderly by systemic recombinant growth factor therapy (rhIGF-I or rhRGF-β): a perspective. Journal of Internal Medicine. 242:285-290.
Bradford MM. (1976). A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry.
72:248-254.
Byers BA and García AJ. (2004). Exogenous Runx2 expression enhances in vitro osteoblastic differentiation and mineralization in primary bone marrow stromal cells. Tissue Engineering.
10:1623-32.
Christian JC, Yu PL, Slemenda CW and Johnston CC Jr.
(1989). Heritability of bone mass: a longitudinal study in aging male twins. The American Journal of Human Genetics. 44:429- 433.
Cheng SL, Shin CS, Towler DA and Civitelli R. (2000). A dominant negative cadherin inhibits osteoblast differentiation.
Journal of Bone and Mineral Research. 15:2362-2370.
Cheung WM, Ng WW and Kung AW. (2006). Dimethyl sulfoxide as an inducer of differentiation in preosteoblast MC3T3-E1 cells. FEBS Letters. 580:121-126.
Cho SH, Kim KG, Kim SR, Lee jA, Moom H and Hwang YY.
(1996). The effects of 17-β estradiol, medroxyprogesterone acetate and parathyroid hormone on the differentiation of osteoblast cell. Korean Journal of Obstetrics & Gynecology.
39:1497-1506.
Delmas PD. (2002). Treatment of postmenopausal osteoporosis.
Lancet 359:2018-2026.
Dinkova-Kostova AT, Liby KT, Stephenson KK, Holtzclaw WD, Gao X, Suh N, Williams C, Risingsong R, Honda T, Gribble GW, Sporn MB and Talalay P. (2005). Extremely potent triterpenoid inducers of the phase 2 response:
correlations of protection against oxidant and inflammatory stress. Proceedings of the National Academy of Sciences of the United States of America. 12:4584-4589.
Ebeling PR, Atley LM and Guthrie JR. (1996). Bone turnover markers and bone density across the menopausal transition. The Journal of Clinical Endocrinology & Metabolism. 81:3366- 3371.
Hauschka PV, Lian JB, Cole DEC and Gundberg CM. (1989).
Osteocalcin and matrix Gla protein: vitamin K-dependent proteins in bone. Physiological Reviews. 69: 990-1047.
Hipskind RA and Bilbe G. (1998). MAP kinase signaling cascades and gene expression in osteoblasts. Frontiers in Bioscience. 3: 804-816.
Hanai Y, Tokuda H and Ishisaki A. (2006). The involvement of p44/p42 MAP kinase in insulin-like growth factor-I-induced alkaline phosphatase activity in osteoblast like MC3T3-E1 cells.
Molecular and Cellular Endocrinology. 251: 42-48.
Ji SH, Ahn DH and Jun M. (2010). Effects of Petasites japonicus and Momordica charantia L. extracts on MC3T3-E1
osteoblastic cells. Journal of Food Science and Nutrition.
39:203-209.
Jie L. (1995). Pharmacology of oleanolic acid and ursolic acid.
Journal of Ethnopharmacology. 49:57-68
Jeong JG, Kim YS, Min YK and Kim SH. (2008). Low concentration of 3-carene stimulates the differentiation of mouse osteoblastic MC3T3-E1 subclone 4 cells. Phytotherapy Research. 22:18-22.
Jun JH, Ryoo HM, Woo KM, Kim GS and Baek JH. (2009).
Bone morphogenetic protein 2-induced MAPKs activation is independent of the Smad1/5 activation. International Journal of Oral Biology. 34:115-121.
Kwak HB, Tang D, Ha H, Lee JH, Kim HN, Woo ER and Hee H. (2006). Tanshinone IIA isolated from Salvia miltiorrhiza Bunge inhibits osteoclast differentiation and bone resorption.
Korean Journal of Medicinal Crop Science. 14:146-147.
Kwon JW, Lee HK, Park HJ, Kwon TO, Choi HR and Song JY. (2011). Screening of biologocal activities to different ethanol extracts of Rubus coreanus Miq. Korean Journal of Medicinal Crop Science. 19:325-333.
Kleerekoper M and Mendlovic DB. (1993). Sodium fluoride therapy of postmenopausal osteoporosis. Endocrine Reviews.
14:312-323.
Kozawa O, Hatakeyama D and Uematsu T. (2002). Divergent regulation by p44/p42 MAP kinase and p38 MAP kinase of bone morphogenetic protein-4-stimulated osteocalcin synthesis in osteoblasts. Journal of Cellular Biochemistry. 84:583-589.
Lee SU, Park SJ, Kwak HB, Oh J, Min YK and Kim SH.
(2008). Anabolic activity of ursolic acid in bone: Stimulating osteoblast differentiation in vitro and inducing new bone formation in vivo. Pharmacological Research. 58:290-296 Lips P. (2001). Vitamin D deficiency and secondary hyperpar
athyroidism in the elderly: consequences for bone loss and fractures and therapeutic implications. Endocrine Reviews.
22:477-501.
Liu J, Wu Q, Lu YF and Pi J. (2008). New insights into generalized hepatoprotective effects of oleanolic acid: key roles of metallothionein and Nrf2 induction. Biochemical Pharmacology. 76:922-928.
Maeda T, Matsunuma A, Kawane T and Horiuchi N. (2001).
Simvastatin promotes osteoblast differentiation and mineralization in MC3T3-E1 cells. Biochemical and Biophysical Research Communications. 280:874-877.
Musabayane CT, Tufts MA and Mapanga RF. (2010).
Synergistic antihyperglycemic effects between plant-derived oleanolic acid and insulin in streptozotocin-induced diabetic rats. Renal Failure. 32:832-839.
Marumo K and Saito M. (2011). Effects of parathyroid hormone (teriparatide) on bone mass and quality in osteoporosis.
Japanese Journal of Clinical Medicine. 69:1264-1269.
Owen TA, Aronow M, Shalhoub V, Barone LM, Wilming L and Tassinari MS. (1990). Progressive development of the rat osteoblast phenotype in vitro: Reciprocal relationships in expression of genes associated with osteoblast proliferation and differentiation during formation of the bone extracellular matrix.
Journal of Cellular Physiology. 143:420-430.
Parfitt AM. (1994). Osteonal and hemi-osteonal remodeling: The
spatial and temporal framework for signal traffic in adult human bone. Journal of Cellular Biochemistry. 55:273-286.
Park HP, Lee JW, Kim HJ and Lee IS. (2005). Effects of Solidago uirga-aurea var. gigantea Miq. root extracts on the activity and differentiation of MC3T3-E1 osteoblastic cell.
Journal of Food Science and Nutrition. 34:929-936.
Pittenger JJ, Mackay AM, Beck SC, Jaiswal RK, Douglas R and Mosca JD. (1999). Multilineage potential of adult human mesenchymal stem cells. Science. 284:143-147.
Sudo H, Kodama H, Amagai Y, Yamamoto S and Kasai S.
(1983). In vitro differentiation and calcification in new clonal osteogenic cell line derived from newbone mouse calvaria. The Journal of Cell Biology. 96:191-198.
Suda T, Takahashi N, Udagawa N, Jimi E, Gillespie MT and Martin TJ. (1999). Modulation of osteoclast differentiation and function by the new members of the tumor necrosis factor receptor and ligand families. Endocrine Reviews. 20:345-357.
Zhou R, Zhang Z, Zhao L, Jia C, Xu S, Mai Q, Lu M, Huang M, Wang L, Wang X, Jin D and Bai X. (2011). Inhibition of mTOR signaling by oleanolic acid contributes to its anti-tumor activity in osteosarcoma cells. Journal of Orthopaedic Research.
29:846-852.
Zhang Y, Li JX, Zhao J, Wang SZ, Pan Y, Tanaka K and Kadota S. (2005). Synthesis and activity of oleanolic acid derivatives, a novel class of inhibitors of osteoclast formation.
Bioorganic & Medicinal Chemistry Letters. 15:1629-1632.