109
Copyright © 2016 The Korean Society of Fisheries and Aquatic Science pISSN:0374-8111, eISSN:2287-8815
서 론
이매패류
(
이하패류)
는분변-
경구감염환을통한세균성및 바이러스성장관계질환의주요전파매개체로간주되고있다(Butt et al., 2004).
이는지역사회에서장관계질환이유행하게 되면해당병원체가하수및오수를통하여담수및기수환경 으로배출되게되고(Da Silva et al., 2007; Iwai et al., 2009),
패 류는여과섭식작용을통해서식해역에존재하는미생물을체 내에농축하여사람이섭취하는경우재감염을유발하기때문 이다(Lees, 2000).
또한전통적으로패류를생식하거나살짝익 혀먹는식습관때문에오염된패류에의한재감염의위험성이 사라지지않고있다(Lees, 2000).
더욱이70
년대이후패류냉 동가공품이국제적으로유통되면서오염된제품에의한감염 의시공간적범위가확대되고있다(Pintó et al., 2009; Polo et al., 2010).
이러한패류에의한감염증발생을예방하고자유럽및북미 대부분국가와남미및아시아일부국가에서는엄격한패류 위생규정을마련하였으며유럽과북미의경우그역사가백
여년에이르고있다
(Lees, 2000).
동규정에는패류생산해역의분변계대장균
(fecal coliform)
농도또는패육중의대장균(Escherichia coli)
농도를기준으로패류생산해역의등급을구 분하고그등급에따라패류의수확전후관리조건을명시하고 있다(European Commission, 2004; US FDA, 2014).
그러나 이러한노력에도불구하고패류생식과관련된감염증이지속 적으로발생하고있으며(Butt et al., 2004),
특히노로바이러스 에오염된굴섭취에따른장관계질환이세계적으로유행하고 있다(Kroneman et al., 2008; Scallan et al., 2011).
오염된활패류중미생물의농도를저감화할수있는방법중 시설내정화및자연정화가현재상업적이용되고있는대표적 인것들이다
(Lees, 2000; Dore et al., 2010).
시설내정화는패자연정화에 의한 양식굴(Crassostrea gigas) 중 노로바이러스 저감화
유홍식*·박용수
1·안세라·박큰바위
1·심길보
1·송기철·이태식
1국립수산과학원 서해수산연구소, 1국립수산과학원 식품위생가공과
Defecation of Norovirus from the Oyster Crassostrea gigas by Depuration Following Translocation of the Growing Area
Hongsik Yu*, Yong Su Park
1
, Sera An, Kunbawui Park1
, Kil Bo Shim1
, Ki Cheol Song and Tae Seek Lee1
West Sea Fisheries Research Institute, National Institute of Fisheries Science, Incheon 22383, Korea
1
Food Safety and Processing Research Division, National Institute of Fisheries Science, Busan 46083, Korea
The efficacy of depuration following growing area translocation for the defecation of norovirus was evaluated under experimental conditions using oysters Crassostrea gigas previously subjected to bioaccumulation of this virus at a waste treatment plant discharge site. Three trials were assayed in an open experimental system with a commercial oyster farm located in a shellfish growing area approved by the Korean Shellfish Sanitation Program. Real-time reverse-transcription polymerase chain reaction (RT-PCR) was used to quantify viruses in the digestive glands of oysters. The final viral loads in oysters after 12 days remained under the detection limit (10 copies/g digestive gland) of the real-time RT-PCR. This reduction trend showed two-phase removal kinetics, with an initial slow reduction or slight increase in viruses during the first 2 days of depuration and subsequent stabilization with 0.12 to 2.64 log unit norovirus copies/g digestive gland per 2 days of depuration for the remaining time.
Key words: Norovirus, Crassostrea gigas , Oyster, Translocation, Depuration
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial Licens (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
http://dx.doi.org/10.5657/KFAS.2016.0109 Korean J Fish Aquat Sci 49(2) 109-115, April 2016
Received 29 February 2016; Revised 29 March 2016; Accepted 30 March 2016
*Corresponding author: Tel: +82. 32. 745. 0751 Fax: +82. 32. 745. 0619
E-mail address: [email protected]
류를탱크에담고위생처리용수에최대
48
시간노출시키는것 을말하며자연정화는오염된해역에서양성된패류를청정해 역으로이동시켜수주에서수개월양성하는것을말한다.
이방 법들은인위적또는자연환경하에서패류의여과작용을유도 하여장내의내용물과함께병원성미생물이배출되도록하는 동일한원리를이용하고있다(Lees, 2000; Nappier et al., 2008;
Oh et al., 2012; Polo et al., 2014a).
시설내정화는유럽
,
미국및일본등에서널리상업적으로 이용되고있으며대장균농도가다소높은패류를생식용으로 출하하기전반드시거쳐야하는강제적규제항목중하나이다(Lees, 2000; European Commission, 2004; US FDA, 2014).
한편시설내정화는인공적으로조성된환경에서패류에먹이 를공급하지않고이루어지기때문에수일이상실시할경우수 질악화로폐사가발생하고자가소화에인한중량감소등패류 의상업적가치가하락되기싶다
(Lees, 200; Dore et al., 2010).
그래서시설내정화는대개
24
시간전후로이루어지며노로바 이러스나A
형간염바이러스등과같은장관계바이러스를충분 히제거할수없어바이러스저감화대책으로활용하기위해서 는개선의여지가많은것으로알려져있다(Lees, 2000; Rich- ards et al., 2010; Polo et al., 2014a; Polo et al., 2014b).
유럽에 서시설내정화후생식용기준에부합되는패류에의해장관 계바이러스감염증이발생한여러사례들이이를뒷받침하고 있다(Chalmers and McMillan, 1995; Le Guyader et al., 2003, 2006; Pintó et al., 2009).
자연정화는대상해역의위생학적상태에따라릴레잉
(relay- ing)
과트랜스로케이션(translocation)
으로그개념이대별된다(European Commission, 2004; US FDA, 2014; DAF, 2011).
릴레잉은등급이지정되고수확이허가된패류생산해역간의 이동을말하며트랜스로케이션은등급미지정또는수확이금 지된해역에서 생산된 패류를등급이지정되고 수확이 허가 된해역으로이동하는것을의미한다
(European Commission, 2004; US FDA, 2014; DAF, 2011).
릴레잉은유럽,
미국및호 주등에서공식적으로활용되고있으며해역이동수주후상업 적목적의패류수확이허용된다.
트랜스로케이션은호주등일 부국가에서채택되고있는위생처리기법으로상대적으로오 염도가심한패류를대상으로하고있어해역이동최소2
개월경과후상업적목적의패류수확이허용된다
(DAF, 2011).
자연정화는시설내정화에비하여적지선정의어려움등의제한 이있으나폐사율이낮고장기간정화작용을통한바이러스제 거가능성이높아주목을받고있다
(Dore et al., 2010).
패류중바이러스의생물축적과배출은패류와바이러스의생 물학적특성에영향을받는복잡한생리과정이다
(Lees, 2000;
Nappier et al., 2009; Oh et al., 2012; Polo et al., 2014b).
최근 연구결과에따르면특정바이러스의생물축적에는패류조직 중특이바이러스수용체가관여하며,
이로인하여패류와바이 러스의종류에따라배출속도도달라진다고한다(Le Guyader
et al., 2006; Tian et al., 2006, 2007; Maalouf et al., 2011).
그러 므로새로운패류위생관리방안의강구및기존정화공정의개 선과관련규정의마련에는바이러스-
패류상호작용과정화요 구시간과같은기초정보가필요하다.
이연구에서는최근패류 위생분야의주요위해요소인노로바이러스에오염된굴을청 정해역으로이동시켜양성하는자연정화(translocation)
를통한 굴중노로바이러스저감화가능성을확인하고자하였다.
재료 및 방법
대상 패류 및 노로바이러스 생물축적
자연정화 대상 패류는 생식을 위주로 소비되는 양식 굴
(Crassostrea gigas)
을선정하였으며,
남해안(
경남통영시자란 만·
사량도일원)
에서2014
년4
월에채취하였다.
채취해역해수 로패각외부를세척하고폐사하였거나파손된굴을제외하였 다.
선별된패류를채취당일에노로바이러스생물축적을위하 여부산시환경공단남부사업소처리하수최종방류지점(
부산 시남구대연동소재)
으로운반하였다.
운반된패류를현장에서30
개의패류양성망(
원추형, ø 40 cm×h 40 cm)
에약30
개체 식나누어담고수심약2-4 m
중에계류시켜1
개월간양성하 면서노로바이러스축적을유도하였다.
노로바이러스축적여 부및그농도확인을위하여매주20
개체씩무작위로채취하여 분석을실시하였다.
굴 중 노로바이러스 분리 및 RNA 추출
굴에서노로바이러스분리는
Jothikumar et al. (2005)
의방 법을일부변형하여사용하였다.
탈각한굴20
개이상으로부 터 분리된소화맹랑(digestive gland)
을 조직파쇄기로균질화 한후3 g
을취하여동량의300 μg/mL Proteinase K solution (Pro mega, USA)
을첨가하고완전히혼합하였다.
혼합된시료 를37℃, 320 rpm
의조건으로1
시간진탕반응시킨후Protein- ase K
의불활성화시키기위하여65℃
에서15
분간방치한후3,000 g
에서5
분간원심분리하여상징액을취하여노로바이러스
RNA
추출시료로사용하였다.
RNA
추출에는Viral RNA mini kit (QIAgen, USA)
을사용 하였다.
상징액300 μL
에AVL buffer 1,120 μL
를첨가하여혼 합한후실온에서10
분간반응시켰다.
반응액에95-100%
에 탄올1,120 μL
를 첨가하여 혼합하였으며,
혼합액을630 μL
를spin column tube
로옮겨6,000 g
에서1
분간원심분리하 였다.
남은혼합액을동일한방법으로처리한후,
동일한spin
column tube
에AW1
완충액500 μL
를첨가하여6,000 g
에서1
분간원심분리하였다.
또한AW2
완충액500 μL
를각각첨가 하여20,000 g
에서3
분간원심분리하였으며,
원심분리후spin
column
에걸러진용액은제거하였다.
다음으로spin column
을새로운tube
로옮긴후AVE
완충액(sodium azide
함유) 60
μ L
를넣고1
분간반응시켰다.
마지막으로6,000 g
로1
분간원심분리하여
real time RT-PCR
을수행하기위한시료로사용 하였다.
Real time RT-PCR
Real time RT-PCR
반응을 위하여OneStep RT-PCR kit (QIAgen, USA)
및RNase inhibi tor (Ambion, USA)
시약을 사용하였다.
폴리오바이러스의RNA
를internal control RNA
(US FDA
제공)
로첨가하여반응이적절히이루어지는지확인하였다
.
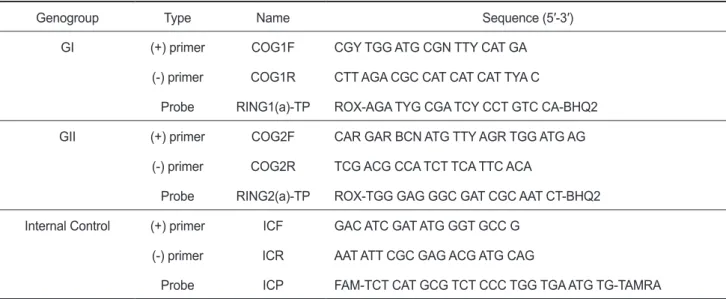
노로바이러스 유전자 검출을 위하여
Table 1
의primer
와probe
를이용하여25×enzyme mix 0.5 μL, 5×buffer 5 μL, 10×dNTPs 1 μL, RNase inhibitor (5 units/ μL) 0.25 μL, 10 μM primer (Forward
및Reverse) 1 μL, 10 μM IC primer (For ward
및Reverse) 0.5 μL, 10 μM probe (
노로바이러스및internal control RNA) 0.5 μL, IC RNA 1 μL,
추출한RNA 5 μ L
로반응액을조성한후,
멸균증류수를첨가하여최종적으로
25 μL
의반응액을조성하였으며,
유전자증폭을위해서는Thermal cycler dice TP800 (Takara, Japan)
를이용하여50℃
에서
50
분간역전사를수행하고, 95℃
에서15
분간DNA
를변 성시켰다.
이후95℃
에서10
초, 53℃
에서25
초, 62℃
에서70
초로
45 cycles
를반복하였다.
양성대조군으로노로바이러스RNA (Takara, Japan)
를사용하고,
음성대조군으로멸균증류 수를사용하였다. Real time RT-PCR
반응은시료마다3
회반 복하였다.
표준곡선 산출 및 노로바이러스 정량
표준곡선산출을위한
realtime RT-PCR
에는pET30a vector
에해당유전자의sequence (GI
형96 bp, GII
형98 bp)
를삽 입하여제작된plasmid DNA (Takara, Korea)
를이용하였다. Shin et al. (2014)
에따라plasmid DNA GI
형과GII
형각각을10
6-10
2copies/reaction
농도로단계 희석한후realtime RT- PCR
을실시하고Thermal cycler dice realtime software (Ta- kara, Japan)
를이용하여표준곡선을산출하였다.
이표준곡선 을근거로반복구3
개의노로바이러스농도(copies/g)
를각각 계산하고이를평균하여시료당노로바이러스농도로하였다. 자연정화
하수처리장최종방류지점에서양성되어노로바이러스가축 적된굴을재수확하여청정해역인한국패류위생계획수출용패 류생산지정해역
2
호(
경남통영시자란만·
사량도해역)
로당일 운반하였다.
양성및운반중폐사한개체를제외한노로바이러 스축적굴을24
개의새로운패류양성망(
원추형, ø 40 cm×h 40 cm)
에30
개체씩나누어담고각양성망을4
단으로연결하였 다. 4
단연결된양성망2
세트(
양성망8
개/
굴약240
개체)
를한 개의시험구로설정하고기존수하식양식시설(
굴수하식수산 업협동조합제공)
에총3
개의시험구(T1-T3)
를20 m
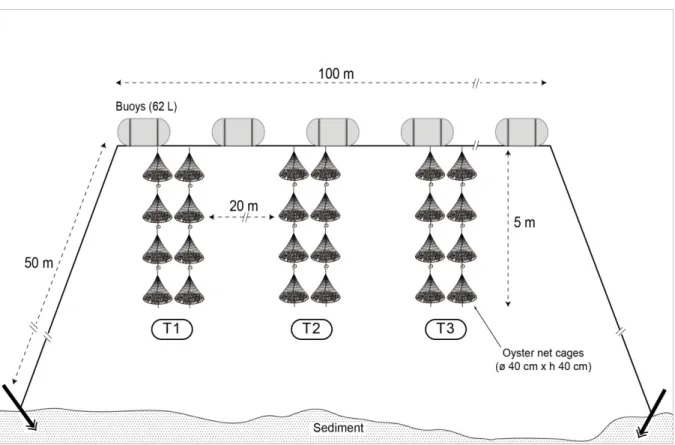
간격으로 설치하여자연정화를유도하였다(Fig. 1).
자연정화기간중2
일간격으로시험구별로굴을20
개체씩수심을고려하지않고 무작위채취하여노로바이러스농도변화를확인하였으며,
패 류의정화작용에영향을미칠수있는해수의수온변화를Tid- biT v2 water temp data logger (Onset Computer Co., U.S.A)
를이용하여1
시간간격으로모니터링하였다.
통계처리
굴 중노로바이러스농도 감소에대한자연정화 효율을평 가하기위한통계처리는
SAS
프로그램을이용한분산분석표 를 작성하였으며, Duncan
의 다중범위검정(Duncan's multi- plerange test)
으로P<0.05
에서결과간의유의성을검정하였다(Steel and Torries, 1980).
또한노로바이러스감소경향은회귀 방정식으로구현하였다.
Table 1. Primers and probes for realtime RT-PCR to quantitate norovirus
Genogroup Type Name Sequence (5′-3′)
GI (+) primer COG1F CGY TGG ATG CGN TTY CAT GA
(-) primer COG1R CTT AGA CGC CAT CAT CAT TYA C
Probe RING1(a)-TP ROX-AGA TYG CGA TCY CCT GTC CA-BHQ2
GII (+) primer COG2F CAR GAR BCN ATG TTY AGR TGG ATG AG
(-) primer COG2R TCG ACG CCA TCT TCA TTC ACA
Probe RING2(a)-TP ROX-TGG GAG GGC GAT CGC AAT CT-BHQ2 Internal Control (+) primer ICF GAC ATC GAT ATG GGT GCC G
(-) primer ICR AAT ATT CGC GAG ACG ATG CAG
Probe ICP FAM-TCT CAT GCG TCT CCC TGG TGA ATG TG-TAMRA
결 과
자연정화가실시된해역의수질은시험기간중특이한변동
은 관찰되지 않았다
.
평균 수온은18.8℃
이었고수온변화는±0.8℃
이었다.
염분농도, pH
및용존산소는시료채취시마다(2
일간격)
측정되었으며그범위는각각31.4-33.7 psu, 7.11- 8.1
및6.11-9.17 mg/L
이었다.
RT-PCR
의 신뢰성이 확인을 위해 첨가한internal control RNA
의사용농도와RT-PCR
에서얻어진cycle threshold (CT)
값을표준곡선에대입하여산출된바이러스농도를비교한결 과, 117
건의모든RT-PCR
반응(39
개시료×3
회반복)
에서얻어진바이러스회수율이모두
90%
이상으로나타나바이러스추출과
RT-PCR
반응이적절히수행되었음을알수있었다.
양식굴을채취하여오염된해역에양성한지
3
주후굴중노 로바이러스농도는평균3.2×10
3copies/g
이었으며그범위는2.0×10
3-4.3×10
3copies/g
이었다.
그리고유전형은모두GII
로밝혀졌다.
노로바이러스가축적된동양식굴을청정해역으 로이동시켜노로바이러스배출을유도한2
차일에3
개의시험 구중2
개의시험구(T1
및T3)
에서노로바이러스농도가0.11
및0.14 log units
정도로소폭증가하였다가이후감소하는경 향을보였고,
시험구T2
는2
일차에거의노로바이러스농도가거의일정하였으며
(0.06 log units
감소),
이후빠른감소경향 을나타내었다(Table 2, Fig. 2).
이후자연정화12
일차에모든 시험구에서노로바이러스농도가검출한계(10 copies/g)
이하 로낮아져축적된노로바이러스가거의모두배출된것으로판 단되었다.
자연정화기간중2
일간격으로분석된시험구별노 로바이러스농도감소는T1
에서0.13-1.56 log units, T2
에서Table 2. Results for the 3 depuration trials with norovirus ex- pressed as number of RNA copies/g digestive gland
Sampling
time Trial
T1 T2 T3
t01 3.3ⅹ103 2.9ⅹ103 4.3ⅹ103
t22 4.5ⅹ103 2.5ⅹ103 5.6ⅹ103
t4 2.5ⅹ103 1.0ⅹ103 3.3ⅹ103
t6 6.2ⅹ102 7.5ⅹ102 2.5ⅹ103
t8 3.7ⅹ102 1.2ⅹ102 1.2ⅹ103
t10 ND3 ND 4.4ⅹ102
t12 ND ND ND
1t0 is sampling at translocation of oyster from bioaccumulation site to depuration site. 2t2-t12 is sampling every 2 days during depu- ration process; 3not detected (below detection limit, 10 copies/g).
Fig. 1. Schematic diagram of the experimental depuration facility for defecation of norovirus from oyster Crassostrea gigas following trans- location of the growing area. T1-T3, experimental depuration trial.
0.13-1.05 log units
그리고T3
에서0.12-2.64 log units
의범위 로나타났으며,
감소추세선의R
2값은0.74-0.93
이었다(Table
2, Fig. 2).
노로바이러스농도감소가가장급격히일어난구간은
T1
및T2
시험구에서정화8-10
일차(
각각1.56 log units
감 소및1.05 log units
감소)
이었고T3
시험구에서는정화10-12
일차(2.64 log units
감소)
이었다(Table 2, Fig. 2).
고 찰
이연구의주목적은
RT-PCR
정량기법을기반으로실험적으로구성된굴자연정화과정의효율성을평가하여양식해역이 동
(translocation)
을통한굴자연정화유도가노로바이러스저 감화기법으로활용가능성이있는지확인하는데있다.
이를위 하여대조구바이러스를이용한노로바이러스회수율검증을 실시하고복잡한패류매트릭스에의한PCR
반응억제여부를 확인할수있도록하여적절하고일관성있는굴중노로바이러 스농도가산출되도록하였다.
그리고굴중노로바이러스배출 과수심과의상관성은적지선정과시설물설계를위한예비시 험에서수심2 m, 7 m
및12 m
로구분하여조사하였는데,
노로 바이러스배출과수심과의유의할만한상관성은발견되지않 았다(data not shown).
자연정화
2
일차에3
개의시험구중2
개의시험구에서노로바 이러스농도가소폭증가하였으며이후안정된감소경향을보 였고,
나머지1
개시험구에서는2
일차까지거의노로바이러스 농도가거의일정하게유지되다가4
일차이후빨리감소하였 다.
그리고12
일차에모든시험구에서노로바이러스농도가검 출한계이하로낮아졌다.
일반적으로대장균
(Escherichia coli)
을대상으로한시설내 정화및자연정화에서는이러한초기농도증가경향이보고된 바없으며단일상정화동태(one-phase depuration kinetics)
가 관찰된다(Dore et al., 2010; Oh et al., 2012; Polo et al., 2014a;
Lees, 2000; Nappier et al., 2009).
이는패류소화맹랑에서대장균농도감소는소화맹랑으로부터배출되는속도에만의존적이 기때문인것으로알려져있다
(Polo et al., 2014a).
그러나굴중 노로바이러스정화에서관찰된복합상정화동태(multi-phase depuration kinetics)
는단순한배출외에다른인자가영향을미 치고있음을나타낸다.
많은연구자들이패류중대부분바이러 스입자가소화맹랑내강에존재하며장관세포에존재하는특이수용체에결합되어있다고보고하고있으며
(Le Guyader et
al., 2006; Mcleod et al., 2009a, 2009b; Tian et al., 2006; Wang et al., 2008),
바이러스정화효율이세균에비하여매우낮은 것은바이러스가이들특이수용체에결합하여세포내대사에 저항력이높기때문으로보고하고있다(Maalouf et al., 2010;
Nappier et al., 2008).
또한Provost et al. (2011)
은패류소화과 정의최종단계에관여하여세균과바이러스의식균작용을담당 하는혈구(hematocytes)
의산소화(acidic digestion)
에대한저 항성이노로바이러스가세균뿐만아니라다른장관계바이러 스에비해서도상대적으로높다고보고한바있다.
이러한이유 로인공배양이불가능한노로바이러스대신feline calicivirus
나murine norovirus
등을대체바이러스로이용하여정화실 험을실시한경우패류장관계세포와의친화력차이로정화효 율이과대평가될수있다는주장도있다(Mcleod et al., 2009b;
Nappier et al., 2008; Provost et al., 2011; Ueki et al., 2007).
이 상의여러연구사례로미루어보아자연정화초기에나타난노 로바이러스지연배출은굴의장관세포수용체에특이적으로 부착된노로바이러스의이탈과소화에일정시간이소요되었기 때문으로 추정된다.
그리고초기에일시적으로노로바이러스 농도가높아지는것은소화맹랑전단계(pre-gland system)
에 서존재하던일부노로바이러스가소화맹랑단계로넘어온후RT-PCR
에의해검출되었기때문으로사료된다. McMenemy
(2014)
에의하면굴에서노로바이러스정화효율은노로바이러스가소화맹랑전단계에서소화맹랑으로들어가는대사속도 와소화맹랑에서배출되는정화속도에지배된다고한다
.
한편대체바이러스를이용하지않고노로바이러스를직접이 Fig. 2. Depuration kinetics of norovirus and its corresponding trend line for each experimental depuration trial (T1-T3). Results are ex- pressed as copies/g digestive gland. Day 0, sampling at translocation of oyster Crassostrea gigas from bioaccumulation site to depuration site; Day 2-12, sampling during depuration process.
Norovirus (copies/g)
0 2 4 6 8 10 12
0 1x103 2x103 3x1033 4x103 5x103 6x103
T1 e-0.1976*(x+42.2612)
R2=0.7359
0 2 4 6 8 10 12
0 1x103 2x103 3x103 4x103 5x103 6x103
T2e-0.2533*(x+31.8454)
R2=0.9291
0 2 4 6 8 10 12
0 1x103 2x103 3x103 4x103 5x103 6x103
T3e-0.1550*(x+55.5636)
R2=0.7874
Day
용한릴레잉에관한보고는매우드물지만
, Dore et al. (2010)
은릴레잉과시설내정화를순차적으로실시하여그효과를보 고한바있다.
동연구에서노로바이러스에오염된굴(
초기농 도1.1×10
3-2.9×10
3copies/g)
을수확하여수온3-4℃
의조건 에서릴레잉을실시한결과,
노로바이러스농도가1 log unit
감 소하는데17
일이소요되었어굴을수확한후수온을상승시켜(15-17 ℃)
시설내정화를추가로실시하였는데4
일만에노로바이러스농도가
1 log unit
추가로감소되었다고하였다. Dore et al. (2010)
은굴의생육한계를벗어나지않는범위내에서수 온이높을수록굴의여과섭식활동과장내분해세균의활성이 증가하여노로바이러스배출이촉진된것으로추측하였다.
이상의연구결과를종합하면노로바이러스에오염된해역에 서양식되어생식에부적합굴이라도청정해역에옮겨
12
일이 상양성되는경우생식에따른노로바이러스감염증발생가능 성을상당히낮출수있을것이라고판단되며동결과는굴에서 노로바이러스저감화대책마련과정화효율개선연구에기초 자료로활용될수있을것으로생각된다.
그러나수온이패류중 바이러스의배출에영향을미치는것으로보고된바있어향후 수온등환경인자와굴성장주기등생리적요소를포함한다 양한잠재영향인자를종합적으로해석한추가연구가필요할 것이며,
나아가패류수확,
해역이동과분망작업등에소요되는 제반비용에대한경제성평가도있어야할것으로사료된다.
사 사
이 논문은
2016
년도 국립수산과학원 수산과학연구사업(R2016059)
의지원으로수행된연구이며연구비지원에감사드립니다
.
References
Butt AA, Aldridge KE and Sanders CV. 2004. Infections re- lated to the ingestion of seafood. Part I: viral and bacte- rial infections. Lancet Infect Dis 4, 201-212. http://dx.doi.
org/10.1016/S1473-3099(04)00969-7.
Chalmers JWT and McMillan JH. 1995. An outbreak of viral gastroenteritis associated with adequately prepared oysters.
Epidemiol Infect 115, 163-167. http://dx.doi.org/10.1017/
S095026 8800058222.
DAF. 2011. Health protocol for the translocation and movement of live bivalve molluscus. Retrieve form https://www.daf.
qld.gov.au/fisheries/aquaculture/management-and-policies/
fampr003-move ment-of-live-bivalve-molluscs on January 2016.
Da Silva AK, Le Saux JC, Parnaudeau S, Pommepuy M, Eimel- ech M and Le Guyader FS. 2007. Evaluation of removal of noroviruses during wastewater treatment, using real time re- verse transcription-PCR: different behaviors of genogroups I and II. Appl Environ Microbiol 24, 7891-7897. http://
dx.doi.org/10.1128/AEM.01428-07.
Dore B, Keaveney1 S, Flannery1 J and Rajko-Nenow1 P. 2010.
Management of health risks associated with oysters harvest- ed from a norovirus contaminated area, Ireland, February–
March 2010. Euro Surveill 15, 1-4.
European Commission. 2004. Regulation (EC) no 854/2004 of the Euro pean Parliament and of the Council of 29 April 2004 laying down specific rules for the organisation of of- ficial controls on products of animal origin intended for hu- man consumption. L226. Off J Eur Communities, 83-127.
Iwai M, Hasegawa S, Obara M, Nakamura K, Horimoto E, Takizawa T, Kurata T, Sogen S and Shiraki K. 2009. Con- tinuous presence of noroviruses and sapoviruses in raw sew- age reflects infections among inhabitants of Toyoma, Japan (2006 to 2008). Appl Environ Microbiol 75, 1264-1270.
http://dx.doi.org/10.1128/AEM.01166-08.
Jothikumar N, Lowther JA, Henshilwood K, Lees DN, Hill VR and Vinj J. 2005. Rapid and sensitive detection of noro- viruses by using TaqMan-based one-step reverse transcrip- tion-PCR assays and application to naturally contaminated shellfish samples. Appl Environ Microbiol 71, 1870-1875.
http://dx.doi.org /doi/10.1128/AEM.71.4.1870-1875.2005.
Kroneman A, Verhoef L, Harris J, Vennema H, Duzier E, van Duynhoven Y, Gray J, Iturriza M, Böttiger B, Falken- horst G, Johnsen C, von Bonsdorff CH,Maunula L, Kuusi M, Pothier P, Gallay A, Schreier E, Höhne M, Koch J, Szücs G, Reuter G, Krisztalovics K, Lynch M, McKeown P, Foley B, Coughlan S, Ruggeri FM, Di Bartolo I, Vainio K, Isak- baeva E, Poljsak-Prijatelj M, Grom AH, Mijovski JZ, Bosch A, Buesa J, Fauquier AS, Hernandéz-Pezzi G, Hedlund KO and Koopmans M. 2008. Analysis of integrated virological and epidemiological reports of norovirus outbreaks collect- ed within the foodborne viruses in Europe Network from 1 July 2001 to 30 June 2006. J Clinical Microbiol 46, 2959- 2965. http://dx.doi.org/10.1128/JCM. 00499-08.
Le Guyader FS, Loisy F, Atmar RL, Hutson AM, Estes MK, Ruvoen-Clouet N, Pommepuy M and Le Pendu J. 2006.
Norwalk virus-specific binding to oyster digestive tissues.
Emerg Infect Dis 12, 931-936. http://dx.doi.org/10.3201/
eid1206.051519.
Le Guyader FS, Neil FH, Dubois E, Bon F, Loisy F, Kohli E, Pommepuy M and Atmar RL. 2003. A semiquantitative ap- proach to estimate Norwalk-like virus contamination of oys- ters implicated in an outbreak. Int J Food Microbiol 87, 107- 112. http://dx.doi.org/10.1016/S0168-1605(03)00058-8.
Lees DN. 2000. Viruses and bivalve shellfish. Int J Food Mi- crobiol 59, 81-116. http://dx.doi.org/ 10.1016/S0168- 1605(00)00248-8.
Maalouf H, Schaeffer J, Parnaudeau S, Le Pendu J, Atmar RL, Crawford SE and Le Guyader FS. 2011. Strain-dependent norovirus bioaccumulation in oysters. Appl Environ Micro- biol 77, 3189-3196. http://dx.doi.org/10.1128/AEM.03010-
Maalouf H, Zakhour M, Le Pendu J, Le Saux JC, Atmar RL 10.
and Le Guyader FS. 2010. Norovirus genogroup I and II ligands in oysters: tissue distribution and seasonal varia- tions. Appl Environ Microbiol 76, 5621-5630. http://dx.doi.
org/10.1128/AEM.00148-10.
McMenemy P. 2014. Depuration dynamics of norovirus in oys- ters. Ph.D. Thesis, University of Stirling, Stirling, U.K.
Mcleod C, Hay B, Grant C, Greening G and Day D. 2009a.
Localization of norovirus and poliovirus in Pacific oys- ters. J Appl Microbiol 106, 1220-1230. http://dx.doi.
org/10.1111/j.1365-2672.2008. 04091.x.
Mcleod C, Hay B, Grant C, Greening G and Day D. 2009b.
Inactivation and elimination of human enteric viruses by Pacific oysters. J Appl Microbiol 107, 1809-1818. http://
dx.doi.org/10.1111/ j.1365-2672.2009.04373.x.
Nappier SP, Graczyk TK and Schwab KJ. 2008. Bioaccu- mulation, retention, and depuration of enteric viruses by
Crassostrea virginica and Crassostrea ariakensis oysters.
Appl Environ Microbiol 74, 6825-6831. http://dx.doi.
org/10.1128/AEM.01000-08.
Nappier SP, Graczyk TK, Tamang L and Schwab KJ. 2010. Co- localized Crassostrea virginica and Crassostrea ariakensis Oysters differ in bioaccumulation, retention and depuration of microbial indicators and human enteropathogens. J Appl Microbiol 108, 736-744. http://dx.doi.org/10.1111/ j.1365- 2672.2009.04480.x.
Oh EG, Yoo HD, Yu HS, Ha KS, Shin SB, Lee TS, Lee HJ, Kim JH and Son KT. 2012. Removal of fecal indicator bacteria from bivalves under natural and electrolyzed water. Ko- rean J Fish Aquat Sci 45, 11-16. http://dx.doi.org/10.5657/
KFAS.2012.0011.
Pint RM, Costafreda MI and Bosch A. 2009. Risk assessment in shellfish-borne outbreaks of hepatitis A. Appl Envi- ron Microbiol 75, 7350-7355. http://dx.doi.org/10.1128/
AEM.01177-09.
Polo D, Álvarez C, Longa A and Romalde JL. 2014a. Depura- tion effectiveness for hepatitis A virus removal from mussels (Mytilus galloprovincialis). Int J Food Microbiol 180, 24- 29. http://dx.doi. org/10.1016/j.ijfoodmicro.2014.04.001.
Polo D, Álvarez C, Vilariño ML, Longa A and Romalde JL.
2014b. Depuration kinetics of hepatitis A virus in clams.
Food Microbiol 39, 103-107. http://dx.doi.org/10.1016/j.
fm.2013.11.011.
Polo D, Vilariño ML, Manso CF and Romalde JL. 2010. Im- ported mollusks and dissemination of human enteric viruses.
Emerg Infect Dis 16, 1036-1038. http://dx.doi.org/10.3201/
eid1606. 091748.
Provost K, Ozbay G, Anderson R, Richards GP and Kingsley DH. 2011. Hemocytes are sites of enteric virus persistence within oysters. Appl Environ Microbiol 77, 8360-8369.
http://dx.doi.org/10.1128/AEM.06887-11.
Richards GP, McLeod C and Le Guyader FS. 2010. Processing strategies to inactivate viruses in shellfish. Food Environ Vi- rol 2, 183-193. http://dx.doi.org/10.1007/s12560-010-9045- Scallan E, Hoekstra RM, Angulo FJ and Tauxe RV. 2011. Food-2.
borne illness acquired in the United States—Major patho- gens. Emerg Infect Dis 17, 7-15. http://dx.doi.org/10.3201/
eid1701.P11101.
Shin SB, Oh EG, Lee HJ, Kim YK, Lee TS and Kim JH. 2014.
Norovirus quantification in oysters Crassostrea gigas col- lected from Tongyeoung, Korea. Korean J Fish Aquat Sci 47, 501-507. http://dx.doi.org/10.5657/KFAS.2014.0501.
Steel RGD and Torrie JH. 1980. Principle and procedure of statistics; a biometrical approach (2nd ed.). Mitchell GJ and Mitchell JH, eds. MacGraw-Hill Book Company, New York, U.S.A., 121-165.
Tian P, Bates AH, Jensen HM and Mandrell RE.2006. Norovi- rus binds to blood group A-like antigens in oyster gastroin- testinal cells. Lett Appl Microbiol 43, 645-651. http://dx.doi.
org/ 10.1111/j.1472-765X.2006.02010.x.
Ueki Y, Shoji M, Suto A, Tanabe T, Okimura Y, Kikuchi Y, Saito N, Sano D and Omura T. 2007. Persistence of cali- civiruses in artificially contaminated oysters during depura- tion. Appl Environ Microbiol 73, 5698-5701. http://dx.doi.
org/10.1128/AEM.00290-07.
US FDA (Food and Drug Adminstration). 2013. National Shell- fish Sanitation Program, Guide for the Control of Molluscan Shellfish, Model ordinance. Retrieved from http://www.fda.
gov/Food/Food Safety/Product-Specific Information/Sea- food/Federal State Programs/National Shellfish Sanitation Program/default.htm on January 18, 2016.
Wang D, Wu Q, Kou X, Yao L and Zhang J. 2008. Distribution of norovirus in oyster tissues. J Appl Microbiol 105, 1966- 1972. http://dx.doi.org/10.1111/j.1365-2672.2008.03970.x.