Identification and Biochemical Characterization of a New Xylan-degrading Streptomyces atrovirens
Subspecies WJ-2 Isolated from Soil of Jeju Island in Korea
Da Som Kim, Chang Hwan Bae, Joo Hong Yeo, and Won-Jae Chi *
Biological and Genetic Resources Assessment Division, National Institute of Biological Resources, Incheon 22689, Republic of Korea
Received: September 12, 2016 / Revised: October 17, 2016 / Accepted: October 17, 2016
Introduction
Plant biomass is the most abundant and commonly used material for the manufacture of bioproducts, and for biofuel production. Xylan, a major structural compo- nent of plant cell wall polysaccharides, is a heteroge- neous polysaccharide with a backbone of β-1,4-linked xylose units that are substituted with acetyl, arabinosyl, and methylglucuronosyl residues [32]. Due to the com- plex heterogeneity of this structure, the synergistic activity of many different xylanolytic enzymes is required.
However, β-1,4-endoxylanases (EC. 3.2.1.8) and β-xylosi-
dases (EC 3.2.1.37) are the principle enzymes involved in xylan backbone hydrolysis. Xylanases are important in a wide variety of biotechnological and industrial appli- cations including pulp bleaching, production of food and animal feed, and dough improvement, and the biocon- version of hemicellulosic biomass to fuels [6, 11, 27].
Recently, antiallergic, antioxidant, antimicrobial and anti-inflammatory activities, and the prebiotic effect of xylooligosaccharides (XOS) have been reported [1, 20].
Hydrolysis of lignocellulosic biomass by microbial enzymes is considered an environmentally friendly alternative to chemical hydrolysis, which generates hazardous byprod- ucts [7]. Thus, several efforts to isolate new microorgan- isms producing more relevant xylanases have been reported [8, 9, 19, 24].
The genus Streptomyces produces numerous chemical A bacterial strain was isolated from a soil sample collected on Jeju Island, Korea. The strain, designated WJ-2, exhibited a high xylanase activity, whereas cellulase activity was not detected. The 16S rRNA gene sequence of WJ-2 was highly similar to type strains of the genus Streptomyces. A neighbor-joining phyloge- netic tree based on 16S rRNA gene sequences showed that strain WJ-2 is phylogenetically related to Strep- tomyces atrovirens. Furthermore, DNA-DNA hybridization analysis confirmed that strain WJ-2 is a novel subspecies of Streptomyces atrovirens. The genomic DNA G+C content was 73.98 mol% and the major fatty acid present was anteiso-C15:0 (36.19%). The growth and xylanase production of strain WJ-2 were signifi- cantly enhanced by using soytone and xylan as nitrogen and carbon sources, respectively. Crude enzyme preparations from the culture broth of strain WJ-2 exhibited maximal total xylanase activities at pH 7.0 and 55 ℃. Thin-layer chromatography analysis revealed that the crude enzyme degrades beechwood xylan to yield xylobiose and xylotriose as the principal hydrolyzed end products.
Keywords: Xylanase, Streptomyces atrovirens, identification, xylanase production, characterization
*Corresponding author
Tel : +82-32-590-7113, Fax: +82-32-590-7472 E-mail: [email protected]
© 2016, The Korean Society for Microbiology and Biotechnology
compounds, including antitumor agents and antibiotics.
Streptomyces is also exploited for industrial applications, as it can be used in the production of important proteins, including proteolytic enzymes and glycosidic hydrolases such as cellulase, xylanase, and agarase [13, 31, 34].
Many of the taxonomical studies of Streptomyces have focused on screening bacterial strains that produce chemical compounds [2]. In this study, we isolated a xylanase producing Streptomyces strain during screen- ing for enzymatic hydrolysis of lignocellulosic polysac- charides from soil samples of Jeju Island, and determined optimum conditions of culture medium for fermentation of this strain.
Materials and Methods
Isolation of bacterial strain
Samples of soil taken from Song-ak mountain on Jeju Island were serially diluted in distilled water (10
-1−10
-5), and 100 μl of each dilution was smeared on an LB agar plate supplemented with 0.1% xylan azure (Sigma, USA). The plates were cultivated at 40 ℃ for 48 h to iso- late thermostable xylanase-producing bacteria. Colonies showing xylanase activity were selected and transferred to fresh LB agar plates. Of these, a colony that we named strain WJ-2 was selected and used for further experiments in this study.
16S rRNA gene sequence and phylogenetic analysis Strain WJ-2 was cultured in LB liquid medium sup- plemented with 0.2% xylan at 40 ℃ for 48 h, after which cells were harvested by centrifugation at 10,840 × g for 20 min. Genomic DNA was extracted and used as the template for amplification of the 16S rRNA gene by PCR with bacterial universal primers, 27F and 1492R [5].
The amplified DNA fragment was directly sequenced using an Applied Biosystems 3730xl DNA Analyzer. The 16S rRNA gene sequence of strain WJ-2 was submitted to the GenBank database using the Basic Local Align- ment Search Tool (BLAST) program [3] at NCBI to search for related strains. The 16S rRNA gene sequences of the related type strains were collected from EzTaxon server [10]. A Neighbor-Joining (NJ) phylogenetic tree was constructed in Mega 6 program using the 16S rRNA gene sequences of strain WJ-2 and related type strains.
Bootstrap analysis was used to evaluate the tree topol-
ogy of the NJ data performing 1,000 resamplings. The evolutionary distance matrix was estimated according to the Kimura two-parameter model [22].
DNA-DNA hybridization
Genomic DNAs from strain WJ-2 and its reference strains, S. atrovirens NRRL B-16357, S. flavoviridis NBRC12772, S. pilosus NBRC12807, S. longispororuber NBRC13488 were prepared from the cells grown on LB plate by using a Genomic DNA Extraction Kit (DyneBio, Korea). E. coli KCCM12119 was used as a negative con- trol. The DNA-DNA hybridization (DDH) experiment was performed with a DIG High Prime DNA Labeling and Detection Starter Kit II according to the supplier’s recommendation (Roche Applied Science, Switzerland) and hybridization signals were calculated with the Quantity One program (Bio-Rad, USA). The signal derived from self-hybridization was taken as 100%.
Phenotypic and physiological characteristics
Gram staining was performed with a Gram stain kit (BD, USA) according to the supplier’s recommendation;
specimens were observed with an optical microscope.
The production of enzymes and carbon utilization of strain WJ-2 were observed using API 20NE and API ZYM strips (Biomérieux, France) according to the manu- facturer’s instructions, with the exception that the bac- terial suspension was prepared in AUX medium supplemented with 1.0% NaCl and trace elements. Sus- ceptibility to various antibiotics including ribostamycin (100 μg/ml), paromomycin (100 μg/ml), thiostrepton (100 μg/ml), kanamycin (100 μg/ml), neomycin (100 μg/
ml), ampicillin (100 μg/ml), apramycin (100 μg/ml), and chloramphenicol (100 μg/ml) was determined on LB plates using the paper disk diffusion method. Growths at different pH values (4.0 −11.0 at intervals of pH 1.0) and at various concentrations of NaCl (0 −10% at inter- vals of 1%) were also determined using LB agar plates.
Characteristics of morphological and physiological differ- entiation
Morphological changes and pigment production of
strain WJ-2 on various agar plates including LB, LBX
(LB containing 0.2% xylan), ISP-2 (BD, USA) and ISP-
4 (BD, USA), were observed after cultivation for 5 d at
40 ℃.
Chemotaxonomic characteristics
Cellular fatty acids were analyzed as methyl esters by gas chromatography (GC) using the Microbial Identifica- tion System (MIDI). Genomic DNA G+C content was determined by reverse phase HPLC as described previ- ously [25].
Optimization of medium compositions
To identify the optimal conditions for enhancing xylanase production, we modified the nitrogen and car- bon sources in the medium. The original medium con- taining 0.2% (NH
4)
2SO
4, 0.05% K
2HPO
4, 0.02% MgSO
4· 7H
2O, 0.001% FeSO
4· 7H
2O (pH 7.2) was prepared according to the composition of the minimal medium (MM) previously described by Hopwood et al. [18].
Instead of (NH
4)
2SO
4, various nitrogen sources, such as bacto peptone, bacto tryptone, soytone, yeast extract, meat extract, and asparagines, were added to original medium at a final concentration of 0.2%. Eight distinct carbon sources, such as maltose, carboxyl methyl cellu- lose (CMC), glucose, xylan, xylose, sucrose, starch, and fructose, were added at a final concentration of 0.1% to medium containing 0.2% soytone as the sole nitrogen source. Additionally, the effect that different carbon source concentrations (0.1 −0.5%) had on xylanase pro- duction was also determined. Culture broths were sam- pled at regular intervals and centrifuged at 10,840 × g for 20 min to obtain cell-free culture supernatant.
Determination of xylanase activity
Xylanase activity in cell-free culture broth was mea- sured using the DNS method with 0.2% (w/v) xylan as a substrate [26]. The reaction was incubated at 40 ℃ for 30 min, and xylanase activity was quantified as described above. One unit (U) of xylanase was defined as the amount of enzyme that produced 1 μmol of xylose per min under the assay conditions. Xylose was used as a reference reducing sugar for preparing standard curve.
Cell growth and xylanase production
Strain WJ-2 was cultured in the optimized medium consisting of 0.2% soytone, 0.05% K
2HPO
4, 0.02%
MgSO
4· 7H
2O, 0.001% FeSO
4· 7H
2O, and 0.3% beech- wood xylan (pH 7.2) and sampled at regular intervals.
Cell mass (wet cell weight) from the culture broth was measured to determine growth curves, and cell-free cul-
ture broth was collected by centrifugation at 18,320 × g for 20 min to measure xylanase activity.
Preparation of crude enzyme
Strain WJ-2 was cultured in the optimized medium at 40 ℃ for 2 d, after which cell-free culture broth was pre- pared by centrifugation. The cell-free culture broth (850 ml) was concentrated overnight by ammonium sul- fate fractionation (75%). After centrifugation at 21,250 × g for 60 min, the precipitate was resuspended in 20 mM Tris-Cl buffer (pH 7.0) and dialyzed against the same buffer for 16 h. The resulting protein sample (85 ml) was prepared to determine optimum conditions for xylanase activity.
Determination of optimum conditions for xylanase activity The optimum pH condition for xylanase activity of crude enzyme was measured at 40 ℃ under various pH conditions using 20 mM MOPS buffer (pH 6.0 −7.0) and 20 mM Tris-Cl buffer (pH 7.0 −9.0). The optimum tem- perature for xylanase activity was measured at across a range of 45 −70℃ (at intervals of 5℃), in 20 mM Tris-Cl buffer (pH 7.0).
Determination of thermostability
The crude enzyme was pre-incubated at different tem- peratures ranging from 55 −65℃ (at intervals of 5℃) for 2 h and sampled at regular intervals. The residual activ- ities were measured at 55 ℃ for 30 min.
Thin layer chromatography (TLC) analysis
The crude enzyme was mixed in 20 mM Tris-Cl (pH 7.0) containing 0.5% beechwood xylan at a final volume of 80 μl. The reaction mixture was incubated at 55℃ for 24 h and sampled at regular intervals. Aliquots of reac- tion mixture (15 μl) were spotted onto silica gel 60 plates (Merck, Germany) and doubly ascended in the solvent system of n-butanol: acetic acid: water (2:1:2, v/v/v), then visualized by heating the plate at 120 ℃ after spraying 10% (w/v) sulfuric acid in 100% (w/v) EtOH.
Results
Phenotypic and chemotaxonomic analysis
Strain WJ-2 was isolated as a xylan-hydrolyzing bac-
terium from a soil sample mixed with decaying wood
(Fig. 1A). The isolate was deposited in the Korean Agri- cultural Culture Collection (KACC) under accession number KACC92087. The isolate was Gram-positive and aerobic. Diffusible pigment was not produced on agar plates such as LB, ISP-2, ISP-4, and R2YE [21].
Gray-colored spores were produced by aerial mycelium
cultured on agar plates (Table 1). Growth was observed between 15 ℃ (very poor) and 50℃, but not at 12℃ and 55 ℃. Growth was observed from pH 6.0 to pH 10.0 (poor at pH 10.0), but not at pH 5.0 and pH 11.0. Cells showed susceptibility to thiostrepton (weak), neomycin (weak), ribostamycin (strong), and paromomycin (strong) but Fig. 1. Isolation of strain WJ-2. (A) Xylanase activity and phenotype of strain WJ-2: Strain WJ-2 was cultured on an ASW-YP agar plate containing 0.1% (w/v) xylan azure at 40 ℃ for 3 d. (B) Construction of a Neighbor-Joining phylogenetic tree based on 16S rRNA gene sequences: The tree is drawn to scale, with branch lengths in the same units as those of the evolutionary distances used to infer the phylogenetic tree. Distances were determined according to the Kimura-two model and bootstrap values (>50%) based on 1,000 replicates are listed as percentages at nodes. Nucleotide sequence accession numbers are given in parentheses. Scale bar, 0.001 substitutions per 100 nucleotides. Evolutionary analyses were conducted in MEGA6. (C) DNA-DNA hybridization analysis: The values obtained from self hybridization of strain WJ-2 were set at 100%. (1), strain WJ-2; (2), S. atrovirens NRRL B-16357; (3), S.
flavoviridis NBRC12772; (4), S. pilosus NBRC12807; (5), S. longispororuber NBRC13488; (6), E. coli KCCM12119.
were resistant to ampicillin, kanamycin, apramycin, and chloramphenicol. Strain WJ-2 was positive for nitrate reduction, agarase, amylase, gelatinase, alkaline phos- phatase, esterase (C4), esterase lipase (C8), leucine aryl- amidase, valine arylamidase, acid phosphatase, naphthol- AS-BI-phosphohydrolase, β-galactosidase, α-glucosi- dase, and α-mannosidase. It was negative for indole pro- duction, arginine dihydrolase, urease, lipase (C14), cystine arylamidase, trypsin, α-chymotrypsin, α-galacto- sidase, β-glucuronidase, N-acetyl-β-glucosaminidase, and α-fucosidase. β-glucosidase was negative for 6-Br-2- naphythyl- β D-glucopyranoside hydrolysis (API ZYM strip) but positive for esculin hydrolysis (API 20NE strip). The strain utilized D-glucose, L-arabinose, D- mannose, D-mannitol, N-acetyl-glucosamine, D-maltose, potassium gluconate, adipic acid (weak) and malic acid, but not trisodium citrate, capric acid and phenylacetic acid. D-Glucose fermentation was negative. The major fatty acids (>4%) were C
15:0Anteiso (36.19%), C
15:0Iso
(10.58%), C
16:0(10.20%), C
16:0Iso (9.84%), and C
14:0Iso (4.18%). The total cellular fatty acid composition is shown in Table 2. The G+C content of the DNA was 73.98 mol%.
Genotypic analysis
Almost the entire 16S rRNA gene sequence (1,399 bp) was amplified from strain WJ-2 by PCR and sequenced.
The 16S rRNA gene sequence analysis of strain WJ-2 revealed that the organism is a member of the genus Streptomyces. Strain WJ-2 had 16S rRNA gene sequence similarity to the type strains Streptomyces longispororuber NBRC13488, Streptomyces atrovirens NRRL B-16357 (99.42%), Streptomyces pilosus NBRC12807 (99.35%), Streptomyces flavoviridis NBRC12772 (99.14%), Strepto- myces griseoflavus LMG19344 (99.14%), Streptomyces speibonae PK-Blue (98.92%), Streptomyces viridodiastat- icus NBRC13106 (98.92%), and Streptomyces albogriseo- lus NRRL B-1305 (98.92%). In a Neighbor-Joining phylogenetic tree based on 16S rRNA gene sequences, strain WJ-2 formed a clade with Streptomyces atrovirens NRRL B-16357 (Fig. 1B). Based on the phylogenetic tree result, Streptomyces atrovirens NRRL B-16357 was used as the reference strain for DNA-DNA hybridization analysis [14]. Several studies indicate that even strains with over 99% 16S rRNA gene sequence similarity may not belong to the same species [23, 30]. Therefore, we performed DNA-DNA hybridization between strain WJ- 2 and the genetically closest reference strains (Fig. 1C).
The values obtained following DNA-DNA hybridization of strain WJ-2 to type strains such as Streptomyces longispororuber NBRC13488 (68.06%), Streptomyces pilosus NBRC12807 (72.53%), Streptomyces flavoviridis NBRC12772 (70.82%) were approximately 70%. How- ever, the DNA relatedness between strain WJ-2 and Streptomyces atrovirens NRRL B-16357 was 87%, indi- cating that these strains represent a single species [35].
Based on DNA-DNA hybridization level (<90%, but
>70%), strain WJ-2 was classified as a subspecies of S.
atrovirens. In summary, three lines of investigation, 16S rRNA gene sequence similarity search, phylogenetic analysis and DNA-DNA hybridization analysis, strongly indicate that WJ-2 should be classified as a novel sub- species of Streptomyces atrovirens, and we therefore pro- pose the name as Streptomyces atrovirens subsp. WJ-2.
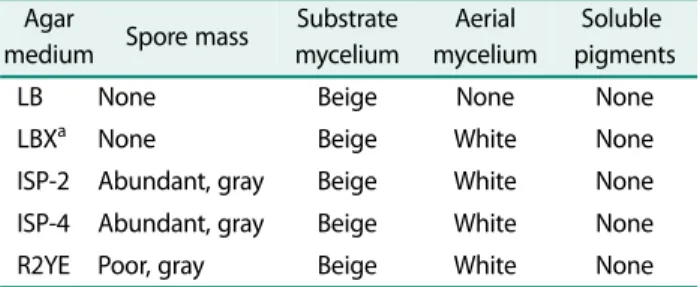
Additional support for this classification comes from the Table 1. Morphological characteristics of strain WJ-2 grown
on different media.
Agar
medium Spore mass Substrate mycelium
Aerial mycelium
Soluble pigments
LB None Beige None None
LBX
aNone Beige White None
ISP-2 Abundant, gray Beige White None ISP-4 Abundant, gray Beige White None
R2YE Poor, gray Beige White None
a
LBX is LB medium containing 0.2% xylan.
Table 2. The cellular fatty acid composition (≥1%) of strain WJ-2.
Fatty acid WJ-2 (%)
C
14:0Iso 4.18
C
15:0Iso 10.58
C
15:0Anteiso 36.19
C
15:02.96
C
16:1Iso H 2.04
C
16:0Iso 9.84
C
16:010.20
C
16:09 Methyl 1.31
C
17:1Anteiso C 1.99
C
17:0Iso 2.52
C
17:0Anteiso 7.54
phylogenetic analysis based on 16S rRNA gene sequence, since the trees constructed by both the NJ and MP methods had almost identical topologies.
Production of xylanase
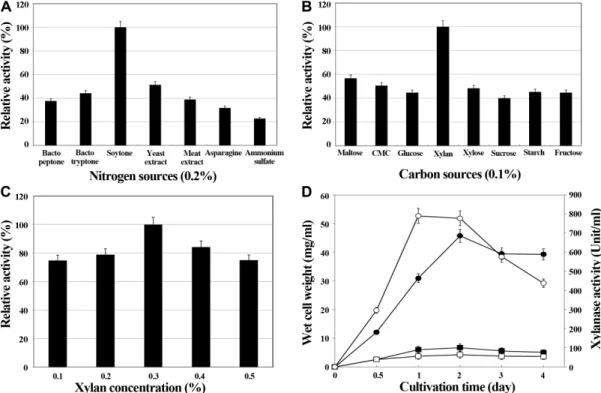
To determine to optimal medium composition for fer- mentation, the effect of various nitrogen and carbon sources on xylanase production was investigated. Soy- tone (0.2%) exhibited the highest xylanase production, which was 4-fold (595 U) that obtained with ammonium sulfate; thus soytone was chosen as the optimal nitrogen source (Fig. 2A). Xylan enhanced xylanase production by approximately 2-fold (602 U) when compared with the other carbon sources, and this was most evident (792 U) at 0.3% xylan concentration (Fig. 2B and 2C). Thus, the optimized medium consisting of 0.2% soytone, 0.05%
K
2HPO
4, 0.02% MgSO
4· 7H
2O, 0.001% FeSO
4· 7H
2O,
and 0.3% beechwood xylan (pH 7.2) was used for fermen- tation of strain WJ-2.
The strain was then inoculated in optimized medium and cultured at 40 ℃ for 4 d with vigorous shaking (Fig.
2D). The cell mass increased sharply during the first 2 d of cultivation, and then decreased slowly; the production of xylanase followed the same pattern. On the other hand, cell growth in the original medium was relatively slow, and xylanase production was very low. Together, these observations indicate that xylan is a important carbon source for xylanase production in this strain, and that enhanced xylan degradation can stimulate growth.
The other carbon sources we tested (including maltose, CMC, glucose, sucrose, xylose, starch, and fructose) were relatively weak stimulators of xylan production. Modifi- cations of the media composition lead to a considerable increase in the production of extracellular xylanase.
Fig. 2. Optimization of medium compositions for xylanase production. (A) Nitrogen source: Different nitrogen sources at 0.2%
concentration were added to medium containing 0.1% beechwood xylan as the sole carbon source. The cells were cultured at 40 ℃
for 2 d. The highest activity observed was considered 100% when calculating the relative activities. (B) Carbon source: Different car-
bon sources at 0.1% concentration were added to medium containing 0.2% soytone as the sole nitrogen source. The cells were cul-
tured at 40 ℃ for 2 d. The highest activity observed was considered 100% when calculating the relative activities. (C) Concentration
of xylan: The cells were inoculated in medium containing 0.2% soytone and different concentrations of xylan, then cultured at
40 ℃ for 2 d. The highest activity observed was considered 100% when calculating the relative activities. (D) Cell growth and
xylanase production in modified medium: The cells were cultured in modified medium at 40 ℃ for 4 d.
■, cell growth from the orig-
inal medium;
●, cell growth from the optimized medium;
□, xylanase activity from the original medium;
○, xylanase activity from
the optimized medium.
Optimum conditions for xylanase activity
The crude enzyme preparations from strain WJ-2 showed the highest total xylanase activity at pH 7.0 when incubated at 40 ℃. The relative activities at pH 6.0 and 8.0 were approximately 70% of maximum activity (Fig. 3A). Maximal activity was also observed at the higher temperature of 55 ℃ at pH 7.0 (Fig. 3B. At this pH, the activities were approximately 80% and 50% at 60 ℃ and 65℃, respectively. The total xylanase activities was maintained at approximately 50% for 20 min at 55 ℃ and thereafter reduced to approximately 30% after a 120-min incubation. However, activity was virtually abolished after incubation for 30 min at 60 ℃ or 20 min at 65 ℃ (Fig. 3C).
Mode of action
The action of the crude enzyme on beechwood xylan was analyzed by TLC using a silica gel 60 plate (Fig.
3D). Spots corresponding to xylotriose (X3) and xylotet- roase (X4) were detected on the plate during the depo- lymerization of xylan. Xylobiose (X2) was also detected by prolonged incubation. No xylose was detected among the products. TLC analysis of xylan hydrolysis products revealed that strain WJ-2 produces endo-type xylanases that degrade xylan into X2 and X3 as the principal hydrolyzed end products.
Discussion
Isolation and characterization of microorganisms that Fig. 3. Optimum conditions for xylanase activity. (A) Effect of pH: The reaction was carried out at 40 ℃ at different pH conditions.
The values obtained at pH 7.0 were set at 100%. All data shown are mean values from at least three replicate experiments.
●, 20 mM MOPS buffer;
■, 20 mM Tris-Cl buffer. (B) Effect of temperature: The reaction was carried out at pH 7.0 at different temperatures.
The values obtained at 55 ℃ were taken to be 100%. The relative activities are the averages from three independent experiments.
(C) Thermostability: The reactions were carried out at different temperatures, 55 ℃, 60℃, and 65℃. The values obtained at zero- incubation time were taken to be 100%. The relative activities are the averages from three independent experiments.
●, 55 ℃;
■, 60 ℃;
◆
, 65 ℃. (D) Thin layer chromatography analysis: The reaction was performed in 20 mM Tris-Cl buffer (pH 7.0) at 55℃ for 48 h. X1,
xylose; X2, xylobiose; X3, xylotriose; X4, xylotetraose. Xylooligosaccharides are indicated by arrows.
produce industrially suitable xylanases is critical for their successful application to biotechnological pro- cesses. In particular, xylanases that are active and sta- ble at high temperature are desirable in various industries. In this study, we found that a moderately thermotolerant Streptomyces atrovirens subspecies iso- lated from soil (strain WJ-2) was an excellent producer of extracellular xylanases. For high-level expression of highly active industrial enzymes, optimizing the culture and reaction conditions is the most critical step [4, 17].
Strain WJ-2 utilized soytone as the most efficient nitro- gen source. We found that xylan was the optimal carbon source for production of xylanase by strain WJ-2. Modifi- cations to the media composition lead to a considerable increase in the production of extracellular xylanase. Cell mass and total xylanase activity derived from cultures of strain WJ-2 in modified medium were almost 9-fold (790 U) higher than those observed in original medium.
Strain WJ-2 exhibited maximum total xylanase activi- ties at pH 7.0 and 55 ℃, which is well within the range of conditions (pH 5.0 −7.0 and at 50−65℃) that yield highest xylanase activity for most of the other known xylanases found in Streptomyces species [4, 9, 15, 33, 37, 38]. The half-life of strain WJ-2 xylanase at 55 ℃ was 20 min, which is similar to xylanases from Streptomyces sp. 7b and Streptomyces halstedii JM8 [4, 29]. These conditions are suitable for hemicellulolytic activities, which facili- tate the utilization of naturally available lignocellulosic materials. Strain WJ-2 xylanase hydrolyzed xylan prin- cipally to X2 and X3, while many others produce XOS with higher degrees of polymerization (DPs) [12, 16, 28].
XOS with DPs of 2 −4 are preferable for use as food ingre- dients, as they are associated with beneficial health effects [36]. These properties of strain WJ-2, as well the high levels of xylanase it produces, suggest that it may be suitable for use in the food and bakery industries, and in industrial processes including utilization of lignocellu- losic biomass for biofuel production, and the biological bleaching of pulp. In this study, we report the isolation of a xylanase-producing soil bacterium from Jeju Island of Korea, and that modifying the culture medium compo- sition enhances xylanase expression. Although several xylanase-producing microorganisms have been reported previously, this is the first description of xylanase pro- duction in an S. atrovirens strain.
Acknowledgments
This work was supported by a grant from the National Institute of Biological Resources (NIBR), funded by the Ministry of Environment (MOE) of the Republic of Korea (NIBR201629101).
References
1. Achary AA, Prapulla SG. 2009. Value addition to corncob: Produc- tion and characterization of xylo-oligosaccharides from alkali pretreated lignin-saccharide complex using Aspergillus oryzae MTCC5154. Bioresour. Technol. 100: 991-995.
2. Al-Bari MAA, Bhuiyan MSA, Flores ME, Petrosyan P, Garcia-Varela M, Islam MAU. 2005. Streptomyces bangladeshensis sp. nov., iso- lated from soil, which produces bis-(2-ethylhexyl) phthalate. Int.
J. Syst. Evol. Microbiol. 55: 1973-1977.
3. Altschul SF, Madden TL, Schaffer AA, Zhang J, Zhang A, Miller W, et al. 1990. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 25: 3389- 402.
4. Bajaj BK, Singh NP. 2010. Production of xylanase from an alkalitol- erant Streptomyces sp. 7b under solid-state fermentation, its puri- fication, and characterization. Appl. Biochem. Biotechnol. 162:
1804-1818.
5. Baker GC, Smith JJ, Cowan DA. 2003. Review and re-analysis of domain-specific 16S primers. J. Microbiol. Methods 55: 541-555.
6. Beg QK, Kapoor M, Mahajan L, Hoondal GS. 2001. Microbial xylanases and their industrial applications: a review. Appl. Micro- biol. Biotechnol. 56: 326-338.
7. Biely P. 1985. Microbial xylanolytic systems. Trends Biotechnol. 11:
286-290.
8. Brennan Y, Callen WN, Christoffersen L, Dupree P, Goubet F, Healey S, et al. 2004. Unusual microbial xylanases from insect guts. Appl. Environ. Microbiol. 70: 3609-3617.
9. Chi WJ, Lim JH, Park DY, Park JS, Hong SK. 2013. Production and characterization of a thermostable endo-type β-xylanase pro- duced by a newly-isolated Streptomyces thermocarboxydus sub- species MW8 strain from Jeju Island. Proc. Biochem. 48: 1736- 1743.
10. Chun J, Lee JH, Jung YY, Kim MJ, Kim SI, Kim BK, et al. 2007.
EzTaxon: a web-based tool for the identification of prokaryotes based on 16S ribosomal RNA gene sequences. Int. J. Syst. Evol.
Microbiol. 57: 2259-2261.
11. Dhiman SS, Sharma J, Battan B. 2008. Industrial applications and future prospects of microbial xylanases: a review. Bioresources 3:
1377-1402.
12. Elegir G, Szakacs G, Jeffries TW. 1994. Purification, characteriza- tion, and substrate specificities of multiple xylanases from Strep- tomyces sp. Strain B-12-2. Appl. Environ. Microbiol. 60: 2609-2615.
13. El-Sersy NA, Abd-Elnaby H, Abou-Elela GH, Ibrahim AHA, El- Toukhy NMK. 2010. Optimization, economization and characteri- zation of cellulase produced by marine Streptomyces ruber. Afr. J.
Biotechnol. 9: 6355-6364.
14. Gause GF, Preobrazhenskaya TP, Sveshnikova MA, Terekhova LP, Maximova TS. 1983. A guide for the determination of actinomy- cetes. Genera Streptomyces, Streptoverticillium, and Chainia. Mos- cow, USSR: Nauka.
15. Georis J, Giannotta F, Buyl ED, Granier B, Frere JM. 2000. Purifica- tion and properties of three endo- β-1,4-xylanases produced by Streptomyces sp. strain S38 which differ in their ability to enhance the bleaching of kraft pulps. Enzyme Microb. Technol. 26: 178- 186.
16. Grabski AC, Forrester IT, Patel R, Jeffries TW. 1993. Characteriza- tion and N-terminal amino acid sequences of beta-(1-4) endox- ylanases from Streptomyces roseiscleroticus: purification incorporating a bioprocessing agent. Protein Expr. Purif. 4: 120-129.
17. Gupta S, Kuhad RC, Bhushan B, Hoondal GS. 2001. Improved xylanase production from a haloalkaliphilic Staphylococcus sp.
SG-13 using inexpensive agricultural residues. World J. Microbiol.
Biotechnol. 17: 5-8.
18. Hopwood DA, Bibb MJ, Chater KF, Kieser T, Bruton CJ, Lydiate DJ, et al. 1985. Genetic manipulation of Streptomyces, A Laboratory Manual, Norwich, UK: The John Innes Foundation.
19. Hwang IT, Lim HK, Song HY, Cho SJ, Chang JS, Park NJ. 2010.
Cloning and characterization of a xylanase, KRICT PX1 from the strain Paenibacillus sp. HPL-001. Biotechnol. Adv. 28: 594-601.
20. Kallel F, Driss D, Chaabouni SE, Ghorbel R. 2015. Biological activi- ties of xylooligosaccharides generated from garlic straw xylan by purified xylanase from Bacillus mojavensis UEB-FK. Appl. Biochem.
Biotechnol. 175: 950-964.
21. Kieser H, Bibb MJ, Buttner MJ, Chater KF, Hopwood DA. 2000.
Practical Streptomyces genetics. The John Innes Foundation, Nor- wich, United Kingdom.
22. Kimura M. 1983. The neutral theory of molecular evolution. Cam- bridge, UK: Cambridge University Press.
23. La Duc MT, Kern R, Venkateswaran K. 2004. Microbial monitoring of spacecraft and associated environments. Microbial. Ecol. 47:
150-158.
24. Lee CC, Kibblewhite-Accinelli RE, Wagschal K, Robertson GH, Wong DW. 2006. Cloning and characterization of a cold-active xylanase enzyme from an environmental DNA library. Extremo- philes 10: 295-300.
25. Mesbah M, Premachandran U, Whitman WB. 1989. Precise mea- surement of the G+C content of deoxyribonucleic acid by high- performance liquid chromatography. Int. J. Syst. Bacteriol. 39:
159-167.
26. Miller GL. 1959. Use of dinitrosalicylic acid reagent for determina- tion of reducing sugar. Anal. Chem. 31: 426-428.
27. Nath D, Rao M. 2001. pH dependent conformational and struc- tural changes of xylanase from an alkalophilic thermophilic Bacil-
lus sp (NCIM 59). Enzyme Microb. Technol. 28: 397-403.
28. Ninawe S, Kapoor M, Kuhad RC. 2008. Purification and character- ization of extracellular xylanase from Streptomyces cyaneus SN32.
Bioresour. Technol. 99: 1252-1258.
29. Ruiz-Arribas A, Fernandez-Abalos JM, Sanchez P, Garda AL, San- tamaria RI. 1995. Overproduction, purification, and biochemical characterization of a xylanase (Xys1) from Streptomyces halstedii JM8. Appl. Environ. Microbiol. 35: 2414-2419.
30. Satomi M, Kimura B, Hamada T, Harayama S, Fujii T. 2002. Phylo- genetic study of the genus Oceanospirillum based on 16S rRNA and gyrB genes: emended description of the genus Oceanospiril- lum, description of Pseudospirillum gen. nov., Oceanobacter gen.
nov. and Terasakiella gen. nov. and transfer of Oceanospirillum jannaschii and Pseudomonas stanieri to Marinobacterium as Mari- nobacterium jannaschii comb. nov. and Marinobacterium stanieri comb. nov. Int. J. Syst. Evol. Microbiol. 52: 739-747.
31. Shin JH, Choi JH, Lee OS, Kim YM, Lee DS, Kwak YY, et al. 2009.
Thermostable xyalanse from Streptomyces thermocyaneoviola- ceus for optimal production of xylooligosaccharides. Biotechnol.
Bioprocess Eng. 14: 391-399.
32. Subramanian S, Sandhia GS, Prema P. 2001. Control of xylanase production without protease activity in Bacillus sp. by selection of nitrogen source. Biotechnol. Lett. 23: 369-371.
33. Techapum C, Charoenrat T, Watanabe M, Sasaki K, Poosara N.
2002. Optimization of thermostable and alkaline-tolerant cellu- lase-free xylanase production from agricultural waste by ther- motolerant Streptomyces sp. Ab106, using the central composite experimental design. Biochem. Eng. J. 12: 99-105.
34. Temuujin U, Chi WJ, Lee SY, Chang YK, Hong SK. 2011. Overex- pression and biochemical characterization of DagA from Strepto- myces coelicolor A3(2): an endo-type β-agarase producing neoagarotetraose and neoagarohexaose. Appl. Microbiol. Bio- technol. 92: 749-759.
35. Van Trappen S, Tan TL, Yang J, Mergaert J, Swings J. 2004. Alter- monas stellipolaris sp. nov., a novel, budding, prosthecate bacte- rium from Antarctic seas, and emended description of the genus Altermonas. Int. J. Syst. Evol. Microbiol. 54: 1157-1163.
36. Vazquez MJ, Alonso JL, Dominguez H, Parajo JC. 2001. Xylooligo- saccharides: Manufacture and applications. Trends Food Sci. Tech- nol. 11: 387-393.
37. Wang SL, Yen YH, Shih IL, Chang AC, Chang WT, Wu WC, et al.
2003. Production of xylanases from rice bran by Streptomyces actuosus A-151. Enzyme Microb. Technol. 33: 917-925.
38. Yan Q, Hao S, Jiang Z, Zhai Q, Chen W. 2009. Properties of a
xylanase from Streptomyces matensis being suitable for xylooli-
gosaccharides production. J. Mol. Catal. B Enzym. 58: 72-77.
국문초록