- 50 -
Upregulation of Prostasin in Nasal Polyps
Joon Hyuk Yoo, MD, Se Jin Park, MD, Heung Man Lee, MD, PhD, Seung Hoon Lee, MD, PhD, Sang Hag Lee, MD, PhD and Tae Hoon Kim, MD, PhD
Department of Otorhinolaryngology-Head and Neck Surgery, Korea University College of Medicine, Seoul, Korea
ABSTRACT
Objectives /Hypothesis : Prostasin is a protease that activates epithelial sodium channels (ENaC), which medi- ate Na+ absorption across epithelial surfaces. Human nasal polyps absorb more ENaC-mediated Na+ than normal mucosa. We investigated the expression and distribution of prostasin in normal mucosa and nasal polyp. Study Design : This was a controlled, prospective study. Methods: The distribution patterns and levels of expression of prostasin in normal sinus mucosa and nasal polyps were evaluated using real-time polymerase chain reaction (PCR), immunohistochemical staining and western blotting. Results : Real-time PCR revealed that the mRNA expression of prostasin was higher in nasal polyps than in normal sinus mucosa. The expression of prostasin was faint in immunohistochemical staining of superficial epithelial cells and submucosal glandular epithelial cells of normal sinus mucosa, but was intense in superficial epithelial cells and submucosal glandular epithelial cells in nasal polyps. Inflammatory cells infiltrating into the nasal mucosa also showed prostasin immunoreactivity in nasal polyps. Western blot analysis with prostasin antiserum detected prostasin protein in normal sinus mucosa and nasal polyps. The expression levels of prostasin were increased in nasal polyps compared with normal sinus mucosa. Conclusions : The levels of expression of prostasin were upregulated in nasal polyps compared to normal sinus mucosa, suggesting a role in the pathogenesis of nasal polyps. However, the exact pathophysiologic function of prostasin in nasal epithelium warrants further investigation.
KEY WORDS : Epithelial Sodium Channels · Nasal Polyps · Prostasin.
INTRODUCTION
The epithelium of the nasal airways is an efficient bar- rier against dust, chemical irritants, and infectious agents.
The coordinated function of the mucous layer, pericilliary layer, and cilia of the epithelium propels the mucus to- ward the nasopharynx, thereby maintaining the integrity of the nasal airway and paranasal sinuses, and providing an important nasal defense mechanism. Electrolyte and water transport by nasal epithelia has been suggested to be important for controlling the quantity and composi- tion of the mucus.1) Previous studies have documented the expression and distribution of various transport proteins in human nasal mucosa, indicating their participation in
nasal fluid production, and have also suggested that altera- tion of their function or expression may contribute to the pathogenesis of nasal polyps.2) 3) Furthermore, the polyp- derived epithelia have a greater rate of transepithelial ion transport than the turbinate-derived epithelia in patients with and without cystic fibrosis, suggesting that increased epithelial fluid absorption contributes to the development of nasal polyps.4)
Prostasin is a glycosylphosphatidylinositol (GPI)-an- chored serine protease that activates epithelial sodium channels (ENaC), which mediate sodium ion (Na+) absorp- tion across human epithelial surfaces.5) 6) Dysregulation of ENaC causes various disorders in the body.7) 8) Interest- ingly, human nasal polyp absorbs more ENaC-mediated Na+ than normal paranasal sinus mucosa.9) Prostasin is produced in a variety of fluid transporting organs, includ- ing the gastrointestinal tract, salivary glands, kidney, and lung.10) 11) Moreover, altered expression of prostasin is also known to cause many human diseases including cystic fi- brosis.11)
In this respect, prostasin may play an important role in fluid and Na+ transport across the epithelium of the na- Address Correspondences and reprint requests to Tae Hoon Kim, MD,
PhD, Department of Otorhinolaryngology-Head and Neck Surgery, Ko- rea University Anam Hospital, Korea University College of Medicine, 126-1, Anam-dong 5-ga, Seoungbuk-gu, Seoul 136-705, Korea Tel : +82-2-920-5486. Fax : +82-2-925-5233.
E-mail : [email protected] Received for publication on March 7, 2012 Accepted for publicatoin on April 9, 2012
sal mucosa and nasal polyps. However, until now, the expression and distributional characteristics of prostasin have not been well-defined in normal human nasal sinus mucosa and nasal polyp. Therefore, we investigated the expression profile and pattern of distribution of prostasin in normal human nasal sinus mucosa and nasal polyp to elucidate the role of prostasin in the development of nasal polyps.
MATERIAL AND METHODS
Tissue Samples and Preparation
Normal ethmoid sinus mucosa (n = 20) was removed from the ethmoid sinus during endoscopic reduction in 20 patients (15 men and five women; age range, 24-42 years) with blowout fracture. The subjects had no history of na- sal infection, allergy, smoking, or ongoing drug treatment.
During the operation, normal-appearing sinus mucosa that was not injured by fracture was removed and used as the normal control. Nasal polyps (n = 20) were taken from 20 patients (12 men and eight women; age range, 24-47 years) undergoing endoscopic sinus surgery for chronic rhinosinusitis with nasal polyps. None of these patients had a history of nasal allergy, asthma, aspirin sensitivity, or ongoing drug treatment. Prior to obtaining the tissue specimens, the protocol and informed consent form were approved by the institutional review board for humans at our institution. Tissue samples (12 normal sinus mucosa and 12 nasal polyps) were cut into two portions. One por- tion was dissected, frozen in liquid nitrogen, and stored at -80°C for subsequent RNA isolation, and the other portion was used for protein isolation. For immunohistochemical analysis, other samples (eight normal sinus mucosae and eight nasal polyps) were fixed overnight in a freshly pre- pared fixative containing 4% paraformaldehyde in phos- phate-buffered saline (PBS, pH 7.4).
Total RNA Isolation and Real Time Polymerase Chain Reaction
The frozen tissues were homogenized in TRIzol re- agent (Gibco BRL, Grand Island, NY, USA), and equal amounts of total RNA from each sample were reverse transcribed in 20 μL of a reaction mixture contain- ing 2.5 U of Moloney murine leukemia virus reverse transcriptase (RT; Gibco BRL) and 50 pmol of random hexanucleotides at 42°C for 60 minutes. Real-time polymerase chain reaction (PCR) was performed using an iQTM Cycler (Bio-Rad, Hercules, CA, USA); each reaction mixture contained 20 μL of iQTM SYBER Green Supermix (Bio-Rad), 0.3 μL of forward primer,
0.3 μL of reverse primer, 5 μL of a 1:10 dilution of a complementary DNA, and 4.4 μL H2O. PCR conditions included a denaturation step (94°C for 4 minutes), am- plification and quantification repeated 35 times (94°C for 15 seconds, 59°C for 20 seconds, and 72°C for 15 seconds), and melting curve analysis (55-95°C with a heating rate of 0.05°C/s). The primers used for the PCR were: prostasin (123 base pairs [bp], sense: 5’- GTGGCTCTCTCGTGTCTGAG-3’; and antisense:5’- CGTCCTCGGAGTAGGAGTCT-3’); glyceraldehyde- 3-phosphate dehydrogenase(GAPDH) (122 bp, sense:
5’-ACAGTCAGCCGCATCTTCTT-3’; and antisense:
5’-GTTAAAAGCAGCCCTGGTGA-3’). To select the primers for real-time PCR, we designed intron span- ning primers to avoid genomic DNA amplification. We then selected the primers that generated a single band of PCR product of the expected size by performing a standard RT-PCR. The final primers were selected based on the linearity of threshold cycle (Ct) values obtained in the serial dilutions of the template, with negative con- trols containing no templates. The measurement of gene expression was assayed in triplicate. The relative gene expression levels were quantified based on the Ct, and where normalized to the reference gene GAPDH.
Immunohistochemical and Western Blot Analysis Immunohistochemical staining was performed using a peroxidase-labeled streptavidin-biotin technique. Brief- ly, paraffin sections (4 μm) were incubated overnight at 4°C with a 1:250 dilution of anti-prostasin polyclonal antibody (ProteinTech Group, Chicago, IL, USA). As a negative control, the primary antibodies were replaced with non-immune rat immunoglobulin G. The color was developed using 3,3'-diaminobenzidine. For Western blot analysis, frozen tissue was crushed and vigorously vor- texed in a buffer solution containing 50 mM Tris-HCl (pH 8.0), 150 mM NaCl, and a proteinase inhibitor cock- tail. The extracted protein (50 μg) was resuspended in sodium dodecyl sulfate (SDS) sample buffer and boiled for 5 minutes. Equal amounts of total protein were sepa- rated by 12% SDS-polyacrylamide gel electrophoresis and transferred to immobilon (Millipore, Bedford, MA, USA). The blots were incubated with anti-prostasin poly- clonal antibody in PBS with Tween 20 overnight at room temperature. Antibody reactions were detected using the enhanced chemiluminescence detection kit (Amersham Biosciences, Piscataway, NJ, USA), followed by detec- tion of chemiluminescence on X-ray film. As an internal control, β-actin expression was analyzed in parallel blots using β-actin antibody (Santa Cruz Biotechnology, Santa Cruz, CA, USA). The intensity of the detected bands was
quantified using Scion Image Beta 4.0.2 software (Scion, Frederick, MD, USA). Relative intensities of each protein signal were obtained by dividing the intensities of each protein signal by β-actin signals. Statistical significance of differences was evaluated by the Mann-Whitney U test, and the level of significance was set at p < .05.
RESULTS
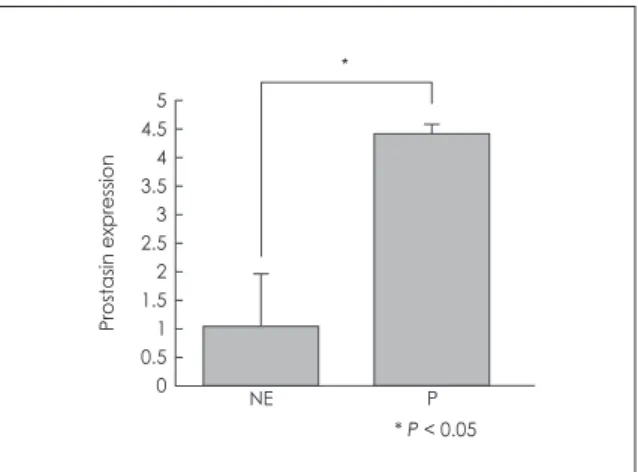
To quantify the amount of prostasin mRNA in normal sinus mucosa and nasal polyp, real-time PCR was per- formed. The mRNA expression of prostasin was higher in nasal polyp compared with that in normal sinus mucosa (Fig. 1).
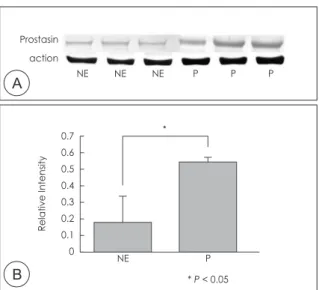
In immunohistochemical staining, expression of prosta- sin was faint in superficial epithelial cells and submucosal glandular epithelial cells of normal sinus mucosa, whereas prostasin was intensely expressed in superficial epithelial cells and submucosal glandular epithelial cells in nasal polyp. In addition, inflammatory cells infiltrating into the nasal mucosa also showed prostasin immunoreactiv- ity in nasal polyp (Fig. 2). Western blot analysis with the prostasin antiserum was able to detect prostasin protein in normal sinus mucosa and nasal polyp. The expression level of prostasin expression was significantly increased in nasal polyp compared with that in the normal sinus mu- cosa (Fig. 3) (p < .05).
DISCUSSION
To ascertain the expression of prostasin in human na- sal mucosa and better understand the pathophysiological role in nasal polyp, the expression and distribution pat-
tern of prostasin in normal sinus mucosa and nasal polyp was studied. The data clearly demonstrate the presence of prostasin mRNA in normal sinus mucosa and nasal pol- yp. Furthermore, the mRNA expression of prostasin was higher in nasal polyp compared with that in normal sinus mucosa. Prostasin immunostaining was detected on epi- thelial cells and submucosal glands in normal sinus mu- cosa and nasal polyp. Furthermore, more immunostaining was detected in nasal polyp than normal mucosa. These tendencies were also found in Western blot analysis. The expression level of prostasin expression was increased in nasal polyp compared with that in the normal sinus mu- cosa. The collective data suggest that prostasin may be involved in the regulation of nasal secretion and the patho- genesis of nasal polyp.
Respiratory mucous membranes begin at the nasal ves- tibule and continue from the pharynx, larynx, and trachea to the bronchioles. By secreting or absorbing solutes, they transport fluid across the epithelium and thereby modify the quantity and composition of the fluid in the luminal compartment. Ion channels in various fluid-transporting epithelia are fundamental to the process of transepithelial electrolytes and fluid absorption or secretion, and have been implicated in the regulation of mucus production.12) 13) Among them, ENaC is critical for mediating Na+ absorp- tion and fluid balance across the epithelial surfaces lining respiratory mucous membranes.14) Recent studies have reported that the protease activity of membrane-tethered channel-activating proteases (CAPs) can activate ENaC and modulate Na+ absorption in human airway epithelial
Fig. 1. Real-time PCR analysis of prostasin in normal ethmoid si- nus mucosa and nasal polyp. The relative gene expression was quantified based on the threshold cycle (Ct), and normalized to the reference gene GAPDH. The bar indicates the mean ± SD. NE, normal ethmoid sinus mucosa; P, nasal polyp. * P < .05.
Fig. 2. Immunohistochemical localization of prostasin in normal nasal ethmoid sinus mucosa (A) and nasal polyp (B). Expression of prostasin was faint in superficial epithelial cells and submu- cosal glandular epithelial cells of normal sinus mucosa, whereas prostasin was intensely expressed in superficial epithelial cells and submucosal glandular epithelial cells in nasal polyp. In ad- dition, inflammatory cells infiltrating into the nasal mucosa also showed prostasin immunoreactivity in nasal polyp. Vertical ar- row indicates epithelial cells, horizontal arrow indicates submu- cosal glands respectively and arrowhead shows inflammatory cells (panels A and B, original magnification, ×100).
A B
NE P
* P < 0.05 5
4.5 4 3.5 3 2.5 2 1.5 1 0.5 0
Prostasin expression
cells.15) Prostasin, the human ortholog of CAP1, is a GPI- anchored serine protease that cleaves the extracellular loop of the ENaC subunit, which increases the probabil- ity that the channel will be in the open configuration.11) Prostasin is detectable in normal human tissues including the prostate gland, liver, salivary gland, kidney, lung, pan- creas, and colon.10) Presently, in the normal sinus mucosa prostasin was localized in the epithelial layer and submu- cosal glands. This finding is consistent with the result that prostasin is expressed in rat kidney in epithelial tissues as ENaC that are known to be the sites of amiloride-sensitive transepithelial Na+ transport.15) Thus, prostasin may be a regulator of Na+/water transport in human sinus mucosa.
Impairment of the physiological condition of respira- tory tracts causes various diseases.16) Previous studies have suggested the possibility that transepithelial ion transport plays an important role in the development of nasal polyposis.4) 17) Furosemide can prevent postsurgical recurrence of sinonasal polyposis, suggesting complex events involving water absorption in the development of nasal polyposis.18) Our previous study using in situ hy- bridization and RT-PCR showed that the levels of expres- sion and pattern of distribution of the mRNA of various ions and water transporters differ between normal nasal mucosa and nasal polyps,2) supporting the suggestion that the altered expression of ion transporters in nasal polyps may be dysfunctional, thus causing impaired electrolyte and water transport across the epithelial cells. Another immunohistochemical staining study for cystic fibrosis transmembrane conductance regulator also revealed a
typical apical distribution in the normal turbinate mucosa, whereas in nasal polyps and postoperative polypoid mu- cosa, a heterogenous pattern of localization was evident.19) In the present study, the expression of prostasin that regu- lates Na+ and water transport occurred at high levels in the epithelial components of nasal polyp compared with those of normal sinus mucosa. These results suggest that the up- regulation of prostasin in nasal polyp may lead to exces- sive proteolytic activation of ENaC, which could play a significant role in the Na+ hyperabsorption characteristic of nasal polyp. Indeed, cystic fibrosis epithelia display increased cell surface expression of processed prostasin, which causes excessive proteolytic activation of ENaC activity and leads to pathological airway surface liquid depletion and impaired mucus clearance from the lung.10) These results indicate that prostasin may participate in the physiology of normal sinus mucosa, but also suggest that their altered expression in nasal polyps may alter Na+ and water transport across the epithelial cells. Further studies are needed to confirm the regulatory roles of prostasin for Na+ and water transport in normal sinus mucosa and nasal polyps.
In conclusion, the present study demonstrates that pros- tasin is expressed in normal sinus mucosa and nasal polyp, where it is localized to the epithelial cells and submucosal glands. In addition, inflammatory cells infiltrating into the nasal mucosa are immunoreactive for prostasin in nasal polyp. The level of expression of prostasin was upregu- lated in nasal polyp compared with normal sinus mucosa, suggesting its roles in the pathogenesis of nasal polyp. Its exact pathophysiologic function in nasal epithelium re- mains to be further investigated.
Acknowledgments
This research was supported by Basic Science Research Program through the National Research Foundation of Korea(NRF) funded by the Ministry of Education, Science and Technology(2011-0014068).
The institutional review boards for human beings at Korea Univer- sity Hospital institution approved the protocols and informed consent form.
저자역할(Author Contributions)
유준혁, 김태훈은 본 연구에서 모든 자료에 접근할 수 있으며 자료의 완전성과 자료 분석의 정확성에 책임을 지고 있습니다. 연구 기획 : 유 준혁, 김태훈, 이상학, 박세진. 자료 해석 및 분석 : 유준혁, 김태훈, 이상학, 박세진. 논문초안 : 유준혁, 김태훈. 논문수정 : 유준혁, 김태 훈. 연구 총괄 : 김태훈.
REFERENCES
1) Takeuchi K, Suzumura E, Hirata K, Majima Y, Sakakura Y. Role of transepithelial ion transport as a determinant of mucus vis- coelasticity in chronic inflammation of the maxillary sinus. Acta Otolaryngol 1991;111:1133-1138.
Fig. 3. Examination of protein production. Western blot analysis of prostasin in normal normal ethmoid sinus mucosa and na- sal polyp(A). Bands were quantified by densitometric scanning and the relative amount of each gene was calculated by di- viding by the internal control, actin(B). The bar indicates the mean ± SD. NE, normal ethmoid sinus mucosa; P, nasal polyp.
* P < .05.
A
B
Prostasin action
NE NE NE P P P
NE P
* P < 0.05 0.7
0.6 0.5 0.4 0.3 0.2 0.1 0
Relative Intensity
2) Lee SH, Park JH, Jung HH, Oh JW, Lee HM, Jun HS, et al. Ex- pression and distribution of ion transport mRNAs in human nasal mucosa and nasal polyps. Acta oto-laryngologica. 2005;125:745- 3) Yasuda M, Niisato N, Miyazaki H, Hama T, Dejima K, Hisa Y, 52.
et al. Epithelial ion transport of human nasal polyp and paranasal sinus mucosa. American journal of respiratory cell and molecular biology. 2007;36:466-72.
4) Bernstein JM, Yankaskas JR. Increased ion transport in cultured nasal polyp epithelial cells. Arch Otolaryngol Head Neck Surg 1994;120:993-996.
5) Chen LM, Skinner ML, Kauffman SW, Chao J, Chao L, Thaler CD, et al. Prostasin is a glycosylphosphatidylinositol-anchored active serine protease. The Journal of biological chemistry.
2001;276:21434-42.
6) Donaldson SH, Hirsh A, Li DC, Holloway G, Chao J, Boucher RC, et al. Regulation of the epithelial sodium channel by serine proteases in human airways. The Journal of biological chemistry.
2002;277:8338-45.
7) Saxena SK, Kaur S. Rab27a negatively regulates CFTR chloride channel function in colonic epithelia: involvement of the effec- tor proteins in the regulatory mechanism. Biochem Biophys Res Commun 2006;346:259-267.
8) Aoi W, Niisato N, Sawabe Y, Miyazaki H, Tokuda S, Nishio K, et al. Abnormal expression of ENaC and SGK1 mRNA induced by dietary sodium in Dahl salt-sensitively hypertensive rats. Cell biology international. 2007;31:1288-91.
9) Yasuda M, Niisato N, Miyazaki H, Iwasaki Y, Hama T, Dejima K, et al. Epithelial Na+ channel and ion transport in human nasal polyp and paranasal sinus mucosa. Biochemical and biophysical research communications. 2007;362:753-8.
10) Yu JX, Chao L, Chao J. Molecular cloning, tissue-specific expres- sion, and cellular localization of human prostasin mRNA. J Biol
Chem 1995;270:13483-13489.
11) Myerburg MM, McKenna EE, Luke CJ, Frizzell RA, Kleyman TR, Pilewski JM. Prostasin expression is regulated by airway surface liquid volume and is increased in cystic fibrosis. Am J Physiol Lung Cell Mol Physiol 2008;294:L932-941.
12) Pitkanen OM, Smith D, O'Brodovich H, Otulakowski G. Expres- sion of alpha-, beta-, and gamma-hENaC mRNA in the human na- sal, bronchial, and distal lung epithelium. Am J Respir Crit Care Med 2001;163:273-276.
13) O'Grady SM, Lee SY. Chloride and potassium channel function in alveolar epithelial cells. Am J Physiol Lung Cell Mol Physiol 2003;284:L689-700.
14) Schild L, Kellenberger S. Structure function relationships of ENaC and its role in sodium handling. Adv Exp Med Biol 2001;502:305-314.
15) Narikiyo T, Kitamura K, Adachi M, Miyoshi T, Iwashita K, Shi- raishi N, et al. Regulation of prostasin by aldosterone in the kid- ney. The Journal of clinical investigation. 2002;109:401-8.
16) Noone PG, Olivier KN, Knowles MR. Modulation of the ion- ic milieu of the airway in health and disease. Annu Rev Med 1994;45:421-434.
17) Bernstein JM, Gorfien J, Noble B, Yankaskas JR. Nasal polyposis:
immunohistochemistry and bioelectrical findings(a hypothesis for the development of nasal polyps). J Allergy Clin Immunol 1997;99:165-175.
18) Passali D, Mezzedimi C, Passali GC, Bellussi L. Efficacy of inhalation form of furosemide to prevent postsurgical relapses of rhinosinusal polyposis. ORL J Otorhinolaryngol Relat Spec 2000;62:307-310.
19) Jang YJ, Lee CH. Localization of cystic fibrosis transmembrane conductance regulator in epithelial cells of nasal polyps and post- operative polypoid mucosae. Acta Otolaryngol 2001;121:93-97.